
Surveillance of Hepatitis E Virus Contamination in Shellfish in China

Abstract
:1. Introduction
2. Experimental Section
2.1. Shellfish Sampling and Processing
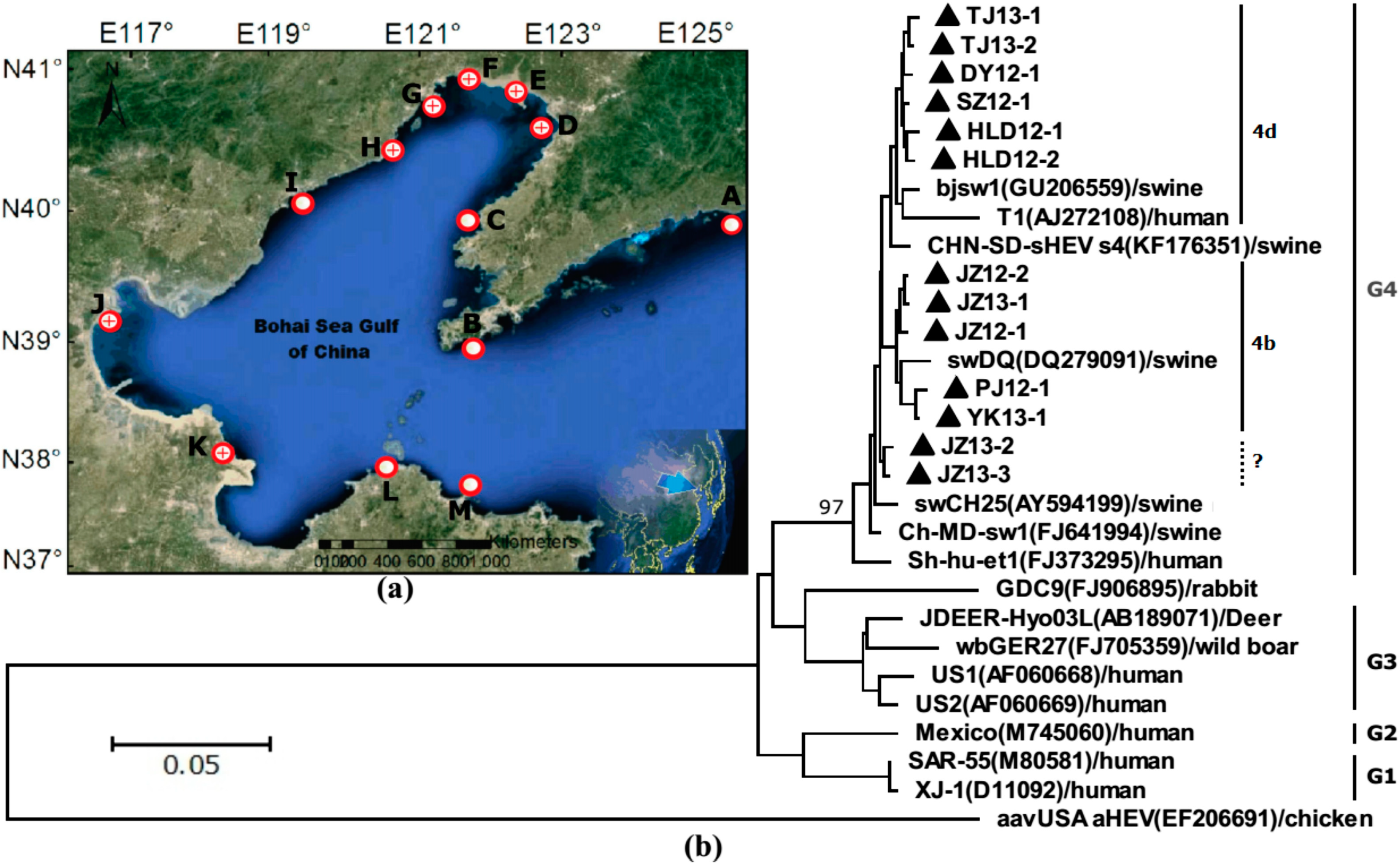
 and that of negative detection’s with a only a red circle
and that of negative detection’s with a only a red circle  . (b) Phylogenetic trees of hepatitis E virus (HEV) were constructed based on partial genomes. Each partial ORF3 (287basepairs) of 13 different clones of JZ12-1, JZ12-2, HLD12-1, HLD12-2, PJ12-1, DY12-1, SZ12-1, JZ13-1, JZ13-2, TJ13-1, TJ13-2, YK13-1and JZ13-3 (with GenBank accession No. KJ816338, KJ816339, KJ816336, KJ816337, KJ816343, KJ816335, KJ816344, KJ816340, KJ816341, KJ816345, KJ816346, KJ816347 and KJ816342, respectively.) was analyzed by the neighbor-joining method. The bootstrap value correspond to 1,000 replications of avian HEV was used as an outgroup. All nucleotide sequences determined in this study were marked by ▲. Other HEV sequences were retrived from GenBank.
and that of negative detection’s with a only a red circle . (b) Phylogenetic trees of hepatitis E virus (HEV) were constructed based on partial genomes. Each partial ORF3 (287basepairs) of 13 different clones of JZ12-1, JZ12-2, HLD12-1, HLD12-2, PJ12-1, DY12-1, SZ12-1, JZ13-1, JZ13-2, TJ13-1, TJ13-2, YK13-1and JZ13-3 (with GenBank accession No. KJ816338, KJ816339, KJ816336, KJ816337, KJ816343, KJ816335, KJ816344, KJ816340, KJ816341, KJ816345, KJ816346, KJ816347 and KJ816342, respectively.) was analyzed by the neighbor-joining method. The bootstrap value correspond to 1,000 replications of avian HEV was used as an outgroup. All nucleotide sequences determined in this study were marked by ▲. Other HEV sequences were retrived from GenBank.
. (b) Phylogenetic trees of hepatitis E virus (HEV) were constructed based on partial genomes. Each partial ORF3 (287basepairs) of 13 different clones of JZ12-1, JZ12-2, HLD12-1, HLD12-2, PJ12-1, DY12-1, SZ12-1, JZ13-1, JZ13-2, TJ13-1, TJ13-2, YK13-1and JZ13-3 (with GenBank accession No. KJ816338, KJ816339, KJ816336, KJ816337, KJ816343, KJ816335, KJ816344, KJ816340, KJ816341, KJ816345, KJ816346, KJ816347 and KJ816342, respectively.) was analyzed by the neighbor-joining method. The bootstrap value correspond to 1,000 replications of avian HEV was used as an outgroup. All nucleotide sequences determined in this study were marked by ▲. Other HEV sequences were retrived from GenBank.
and that of negative detection’s with a only a red circle . (b) Phylogenetic trees of hepatitis E virus (HEV) were constructed based on partial genomes. Each partial ORF3 (287basepairs) of 13 different clones of JZ12-1, JZ12-2, HLD12-1, HLD12-2, PJ12-1, DY12-1, SZ12-1, JZ13-1, JZ13-2, TJ13-1, TJ13-2, YK13-1and JZ13-3 (with GenBank accession No. KJ816338, KJ816339, KJ816336, KJ816337, KJ816343, KJ816335, KJ816344, KJ816340, KJ816341, KJ816345, KJ816346, KJ816347 and KJ816342, respectively.) was analyzed by the neighbor-joining method. The bootstrap value correspond to 1,000 replications of avian HEV was used as an outgroup. All nucleotide sequences determined in this study were marked by ▲. Other HEV sequences were retrived from GenBank.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location A–M | Kg of Shellfish Collected, by Species | HEV RNA Positive R | Final verified S | ||
|---|---|---|---|---|---|
| A. granosa | S. Subcrenata | R. philippinarum | |||
| A: Dandong | NA | 2 | 3 | 0 | |
| B: Dalian | 2 | 3 | 4 | 0 | |
| C: Wafangdian | NA | 3 | 3 | 0 | |
| D: Yingkou | 3 | 5 | 4 | 1 | 1 |
| E: Panjin | 3 | 3 | 4 | 4 | 1 |
| F: Jinzhou | 8 | 6 | 9 | 7 | 5 |
| G: Huludao | 5 | 3 | 3 | 4 | 2 |
| H: Suizhong | 4 | 2 | 2 | 1 | 2 |
| I: Qinhuangdao | NA | 2 | 2 | 0 | 1 |
| J: Tianjin | 5 | 3 | 6 | 4 | 1 |
| K: Dongying | 3 | 4 | 5 | 1 | |
| L: Penglai | NA | 3 | 4 | 0 | |
| M: Yantai | 2 | NA | 3 | 0 | |
| Total ( detection rate C ) | 35 (5/35, 14.3%) | 39 (11/39, 28.2%) | 52 (6/52, 11.5%) | 22 (5 + 11 + 6) (22/126, 17.5%) | 13 |
2.2. HEV Screening and Phylogenetic and Evolutionary Analysis
3. Results and Discussion
| Collection-Dates | Isolation-Sources | Isolates-Tentative Names | GenBank-Accession Numbers |
|---|---|---|---|
| 22/Mar/12 | S. Subcrenata | JZ12-1 | KJ816338 |
| 28/Mar/12 | S. Subcrenata | JZ12-2 | KJ816339 |
| 6/Apr/12 | S. Subcrenata | HLD12-1 | KJ816336 |
| 6/Apr/12 | S. Subcrenata | HLD12-2 | KJ816337 |
| 2/May/12 | R. philippinarum | PJ12-1 | KJ816343 |
| 3/May/12 | R. philippinarum | DY12-1 | KJ816335 |
| 4/Nov/12 | A. granosa | SZ12-1 | KJ816344 |
| 11/Mar/13 | S. Subcrenata | JZ13-1 | KJ816340 |
| 16/Apr/13 | A. granosa | JZ13-2 | KJ816341 |
| 25/Apr/13 | A. granosa | TJ13-1 | KJ816345 |
| 25/Apr/13 | S. Subcrenata | TJ13-2 | KJ816346 |
| 16/Nov/13 | S. Subcrenata | YK13-1 | KJ816347 |
| 19/Dec/13 | R. philippinarum | JZ13-3 | KJ816342 |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yugo, D.M.; Meng, X.J. Hepatitis E virus: Foodborne, waterborne and zoonotic transmission. Int. J. Environ. Res. Public Health 2013, 10, 4507–4533. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Bendall, R.; Legrand-Abravanel, F.; Xia, N.S.; Ijaz, S.; Izopet, J.; Dalton, H.R. Hepatitis E. Lancet 2012, 379, 2477–2488. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Li, L.; Zhu, Y.; Wang, L.; Geng, J.; Chang, Y.; Xue, C.; Du, G.; Li, Y.; Zhuang, H. Hepatitis E virus infection among animals and humans in Xinjiang, China: Possibility of swine to human transmission of sporadic hepatitis E in an endemic area. Am. J. Trop. Med. Hyg. 2010, 82, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; Borentain, P.; Queyriaux, B.; Kaba, M.; Moal, V.; Gallian, P.; Heyries, L.; Raoult, D.; Gerolami, R. Pig liver sausage as a source of hepatitis E virus transmission to humans. J. Infect. Dis. 2010, 202, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Chijiwa, K.; Sera, N.; Ishibashi, T.; Etoh, Y.; Shinohara, Y.; Kurata, Y.; Ishida, M.; Sakamoto, S.; Takeda, N.; et al. Hepatitis E virus transmission from wild boar meat. Emerg. Infect. Dis. 2005, 11, 1958–1960. [Google Scholar] [CrossRef] [PubMed]
- Tei, S.; Kitajima, N.; Takahashi, K.; Mishiro, S. Zoonotic transmission of hepatitis E virus from deer to human beings. Lancet 2003, 362, 371–373. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.Q.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the ICTV; Elsevier/Academic Press: London, UK, 2012; pp. 1021–1028. [Google Scholar]
- Zhu, Y.M.; Dong, S.J.; Si, F.S.; Yu, R.S.; Li, Z.; Yu, X.M.; Zou, S.X. Swine and human hepatitis E virus (HEV) infection in China. J. Clin. Virol. 2011, 52, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Maalouf, H.; Schaeffer, J.; Parnaudeau, S.; Le Pendu, J.; Atmar, R.L.; Crawford, S.E.; Le Guyader, F.S. Strain-dependent norovirus bioaccumulation in oysters. Appl. Environ. Microbiol. 2011, 77, 3189–3196. [Google Scholar] [CrossRef] [PubMed]
- Muniain-Mujika, I.; Calvo, M.; Lucena, F.; Girones, R. Comparative analysis of viral pathogens and potential indicators in shellfish. Int. J. Food Microbiol. 2003, 83, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Grodzki, M.; Schaeffer, J.; Piquet, J.C.; Le Saux, J.C.; Cheve, J.; Ollivier, J.; Le Pendu, J.; Le Guyader, F.S. Bioaccumulation efficiency, tissue distribution, and environmental occurrence of hepatitis E virus in bivalve shellfish from France. Appl. Environ. Microbiol. 2014, 80, 4269–4276. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.D.; Metcalf, T.G. Polyethylene glycol precipitation for recovery of pathogenic viruses, including hepatitis A virus and human rotavirus, from oyster, water, and sediment samples. Appl. Environ. Microbiol. 1988, 54, 1983–1988. [Google Scholar] [PubMed]
- Balayan, M.S. Hepatitis E virus infection in Europe: Regional situation regarding laboratory diagnosis and epidemiology. Clin. Diagn. Virol. 1993, 1, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Said, B.; Ijaz, S.; Kafatos, G.; Booth, L.; Thomas, H.L.; Walsh, A.; Ramsay, M.E.; Morgan, D. Hepatitis E outbreak on cruise ship. Emerg. Infect. Dis. 2009, 15, 1738–1744. [Google Scholar] [CrossRef] [PubMed]
- Torne, J.; Miralles, R.; Tomas, S.; Saballs, P. Typhoid fever and acute non-A non-B hepatitis after shellfish consumption. Eur. J. Clin. Microbiol. Infect. Dis. 1988, 7, 581–582. [Google Scholar] [CrossRef] [PubMed]
- Li, T.C.; Miyamura, T.; Takeda, N. Detection of hepatitis E virus RNA from the bivalve Yamato-Shijimi (Corbicula japonica) in Japan. Am. J. Trop. Med. Hyg. 2007, 76, 170–172. [Google Scholar] [PubMed]
- Crossan, C.; Baker, P.J.; Craft, J.; Takeuchi, Y.; Dalton, H.R.; Scobie, L. Hepatitis E virus genotype 3 in shellfish, United Kingdom. Emerg. Infect. Dis. 2012, 18, 2085–2087. [Google Scholar] [CrossRef] [PubMed]
- Namsai, A.; Louisirirotchanakul, S.; Wongchinda, N.; Siripanyaphinyo, U.; Virulhakul, P.; Puthavathana, P.; Myint, K.S.; Gannarong, M.; Ittapong, R. Surveillance of hepatitis A and E viruses contamination in shellfish in Thailand. Lett. Appl. Microbiol. 2011, 53, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, J.; Yuan, Q.; Ge, S.; Zhang, J.; Xia, N.; Tian, D. Clinical characteristics and risk factors of sporadic Hepatitis E in central China. Virol. J. 2011, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; He, Y.; Wang, H.; Shen, Q.; Cui, L.; Wang, X.; Shao, S.; Hua, X. Hepatitis E virus genotype diversity in eastern China. Emerg. Infect. Dis. 2010, 16, 1630–1632. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization. Microbiology of Food and Animal Feed: Horizontal Method for Detection of Hepatitis A Virus and Norovirus in Food Using Real-Time RT-PCR: Part 2: Method for Qualitative Detection (ISO/TS 15216–2:2013); International Organization for Standardization: Geneva, Switzerland, 2013. [Google Scholar]
Appendix





© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Li, D.; Zha, E.; Zhou, T.; Wang, S.; Yue, X. Surveillance of Hepatitis E Virus Contamination in Shellfish in China. Int. J. Environ. Res. Public Health 2015, 12, 2026-2036. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph120202026
Gao S, Li D, Zha E, Zhou T, Wang S, Yue X. Surveillance of Hepatitis E Virus Contamination in Shellfish in China. International Journal of Environmental Research and Public Health. 2015; 12(2):2026-2036. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph120202026
Chicago/Turabian StyleGao, Shenyang, Dandan Li, Enhui Zha, Tiezhong Zhou, Shen Wang, and Xiqing Yue. 2015. "Surveillance of Hepatitis E Virus Contamination in Shellfish in China" International Journal of Environmental Research and Public Health 12, no. 2: 2026-2036. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph120202026