3.2. Decolorization of Dye CR and RTE in WB Medium
While numerous studies on the removal of azo dyes by biodegradation exist, there is a lack of information on the use of agricultural wastes as microbial growth media [
1,
17,
19,
27]. Agricultural waste has begun to attract attention as microbial growth media as a solution to the high cost of routinely used defined growth media. Recently, the widely generated and abundantly available major agricultural waste wheat bran has been used as the growth medium for the enrichment of microbial consortia and their use in the biodegradation of the benzidine-based azo dye Trypan blue [
22]. Since wheat bran has been previously found suitable for the growth of microorganisms and their use in the degradation of azo dyes at flask studies [
22]; it has now been further evaluated with a PUF-immobilized microbial consortium for the mineralization and detoxification of the model azo dye CR and RTE under batch and laboratory scale continuous upflow column bioreactor conditions.
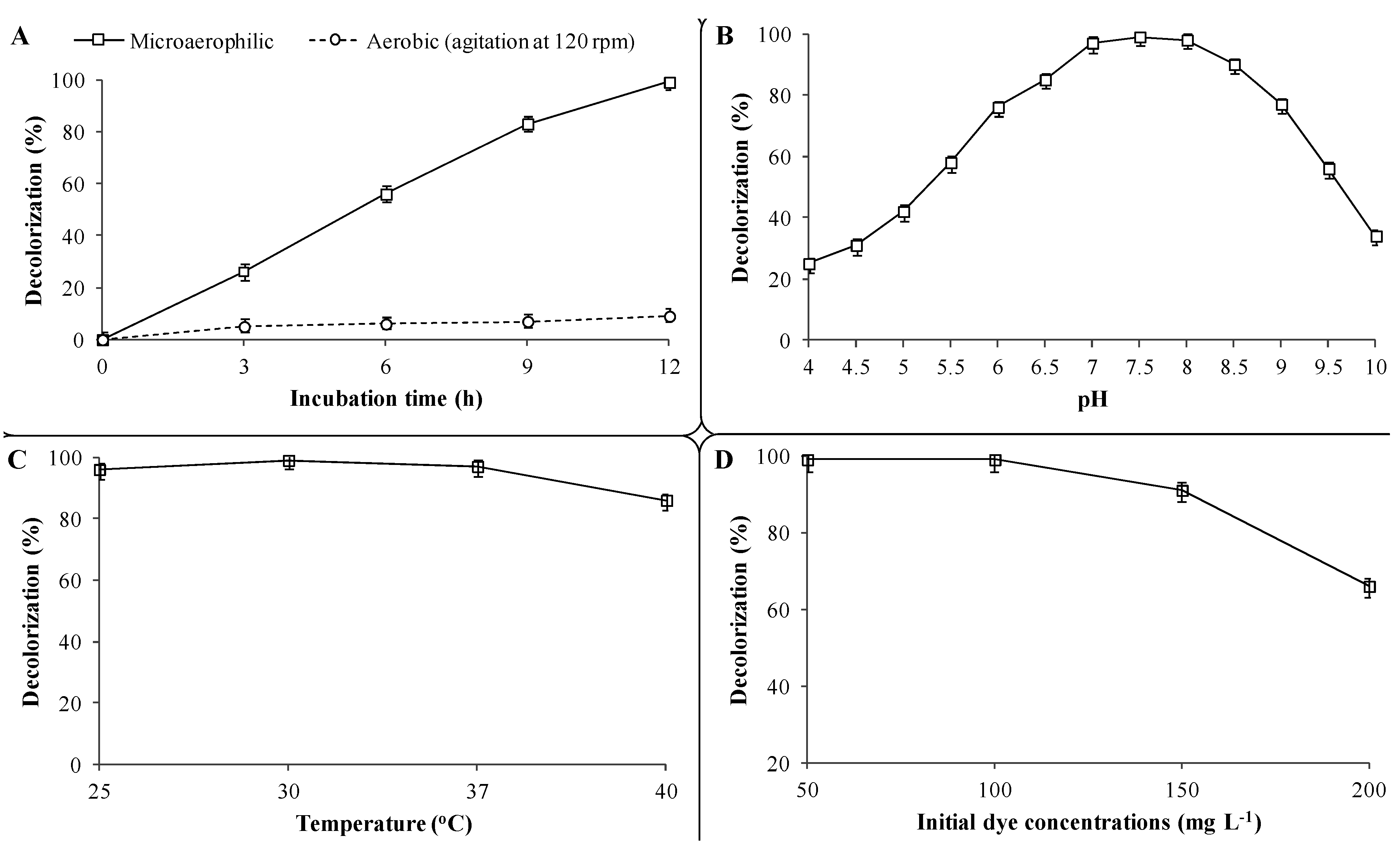
Results of the batch experiments with the PUF-immobilized microbial consortium indicated that microaerophilic incubation favors CR decolorization as 99% of dye (100 mg·L
−1) removal was observed within 12 h at 30 ± 0.2 °C (
Figure 3A). On the contrary, only 9% dye removal was achieved within the same time and a maximum of 14% after 48 h under aerobic incubation conditions. The microbial decolorization of azo dyes is an enzymatic reaction and the presence of molecular oxygen has a strong inhibitory effect on dye removal [
43] primarily due to competition in the oxidation of reduced electron carriers like NADH with either oxygen or azo groups as the electron receptor [
44]. This perception has been well supported in a previous study, where oxygen-rich conditions had a strong inhibitory effect on the decolorization of the diazo dye Reactive blue 160 by mixed BDN cultures as only 24% dye removal was observed within 4 h, whereas more than 90% removal was achieved within same time under microaerophilic conditions [
45]. Therefore, microaerophilic incubation conditions are necessary for azo dye reduction and thus further CR azo dye decolorization experiments with the PUF-immobilized microbial consortium were only performed using microaerophilic incubations.
Figure 3.
Batch studies on the effect of operational variables on the decolorization of CR dye by a PUF-immobilized microbial consortium. (A) Microaerophilic and aerobic incubation; (B) pre-grown culture medium Ph; (C) incubation temperatures and (D) initial CR dye concentrations. The percent decolorization was measured after 12 h of incubation at 30 ± 0.2 °C unless otherwise stated. Data points indicate the mean of three independent replicates; ±Standard errors of mean (SEM) is indicated by error bars.
Figure 3.
Batch studies on the effect of operational variables on the decolorization of CR dye by a PUF-immobilized microbial consortium. (A) Microaerophilic and aerobic incubation; (B) pre-grown culture medium Ph; (C) incubation temperatures and (D) initial CR dye concentrations. The percent decolorization was measured after 12 h of incubation at 30 ± 0.2 °C unless otherwise stated. Data points indicate the mean of three independent replicates; ±Standard errors of mean (SEM) is indicated by error bars.
The effect of different pre-grown culture medium pH on CR dye (100 mg·L
−1) decolorization by the PUF-immobilized microbial consortium showed that the maximum dye removal (99%) was achieved at pH 7.5 within 12 h (
Figure 3B). Further increases or decreases in culture medium pH towards slightly basic (7.0) or alkaline (8.0) conditions showed marginal decreases in dye removal (97% and 98%, respectively) within the same timeframe, whereas extreme acidic (4–6) or alkaline (9.0–10) pH values strongly retarded the decolorization performance (25% and 34%, respectively). Although the immobilized microbial consortium was active at extreme acidic and alkaline pHs, the enzyme activities might not be induced sufficiently to achieve optimum dye removal. The original pH of the pre-grown microbial consortium culture used in this study was 7.55 (without any pH adjustment). These results are in good agreement with a study where a microbial consortium composed of 15 different bacteria was proficient in the decolorization of the diazo dye Trypan blue (50 mg·L
−1) at pH 7.0, while a slight decrease (6.0) or an increase (8.5) in pH did not affect decolorization rates much [
22]. These results indicated that the PUF-immobilized microbial consortium was able to decolorize the azo CR dye in a basic to alkaline pH range and thus could be used for treatment of dye-containing wastewaters with variable pH ranges. It is known that textile effluents have a characteristic alkaline pH range [
46].
Decolorization at different incubation temperatures revealed that the PUF-immobilized microbial consortium was able to remove the CR dye (100 mg·L
−1) over a wide range of temperatures
i.e., 25–40 ± 0.2 °C. However, maximum dye removal (99%) was observed at an optimum incubation temperature of 30 ± 0.2 °C within 12 h, while 96%, 97% and 86% removal was achieved at 25, 37 and 40 ± 0.2 °C temperatures, respectively, within same time (
Figure 3C). The same results were reported by Kadam
et al. for the decolorization of the diazo dye Reactive rd 120 by a developed microbial consortium consisting of 10 different bacteria, which exhibited maximum dye removal at 30 ± 0.2 °C incubation temperature [
29]. A previous study also showed that the enriched microbial consortium gave the maximum decolorization at 30 ± 0.2 °C while incubation at lower and higher temperatures than the optimum marginally affected decolorization rates [
22]. The incubation temperature affects both microbial growth and enzyme activities, and thus the rate of dye decolorization [
47]. Beyond the optimum temperature, the loss of cell viability and decreased rates of enzymes production occur, which result in reduced azo dye decolorization [
2]. Thus, it was clear that, though the maximum dye decolorization often corresponds to the optimum incubation temperature, the PUF-immobilized microbial consortium was well adapted to a wide range of temperatures
i.e., 25 to 40 ± 0.2 °C for efficient CR dye decolorization.
Textile wastewaters are known to contain structurally different and varied concentrations of dyes ranging from 10 to 200 mg·L
−1 [
48]. Therefore, the microorganisms employed for decolorization of dyes-containing wastewaters must be able to tolerate high concentrations of dyes and should be metabolically active to mineralize them. The result of the decolorization at different dye concentrations showed that a maximum 100 mg·L
−1 of dye CR was efficiently decolorized by the PUF-immobilized microbial consortium within 12 h (
Figure 3D). Further increases in dye concentration up to 150 mg·L
−1 slightly reduced the decolorization rate as 91% removal was observed within 12 h. However, at 200 mg·L
−1 of CR dye concentration, a strong inhibition of decolorization was observed and only 66% removal was achieved within 12 h and as much as 20 h were needed for complete decolorization (data not shown). The ability of PUF-immobilized microbial consortium to decolorize a maximum of 100 mg·L
−1 of CR dye in agricultural waste WB medium thus indicated its great potential to be used for treatment of azo dye-containing wastewater at low-cost.
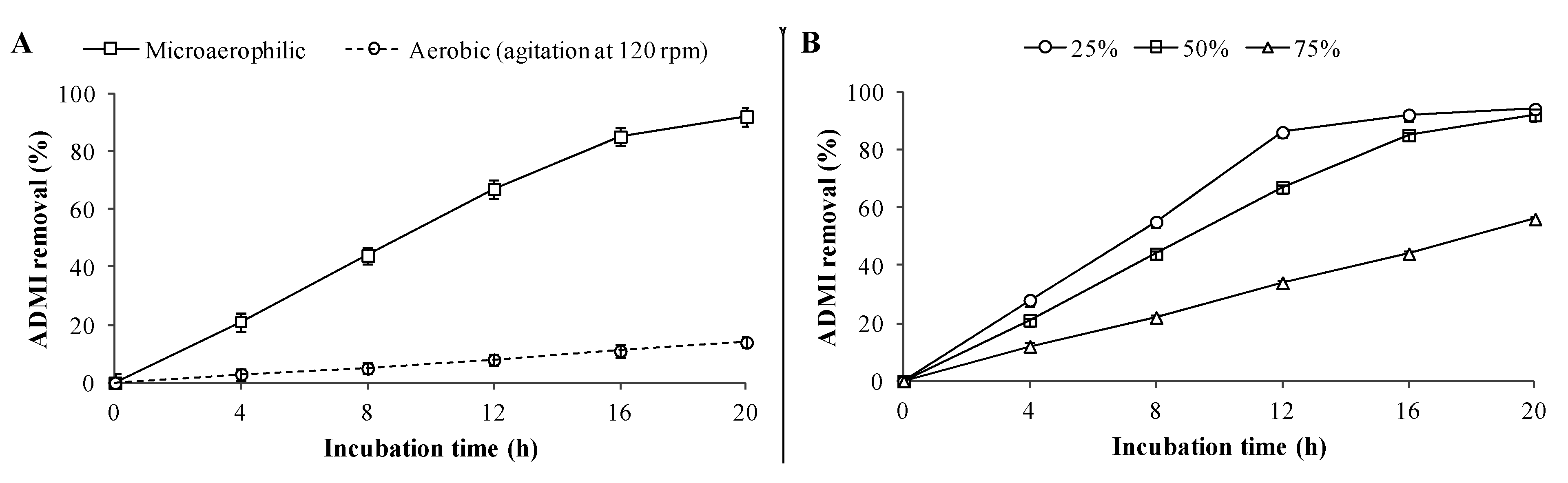
Additionally, batch studies on the decolorization of RTE (50% in WB medium) using PUF-immobilized microbial consortium were also carried out, which showed 92% ADMI removal within 20 h under microaerophilic incubation conditions (
Figure 4A), whereas only 12% ADMI removal was observed under aerobic incubation and a maximum of 16% within 48 h (data not shown). Thus, it would be essential to use microaerophilic conditions to achieve enhanced decolorization rates of textile effluents. The reduced decolorization performance under aerobic incubation conditions might be due to the presence of higher amounts of azo dyes in the RTE as the chromophoric azo groups of azo dyes get cleaved by azoreductase only in the absence of or with a limited supply of oxygen [
49]. The decolorization results for different RTE concentrations revealed that a maximum of 94% and 92% ADMI removal were achieved for 25% and 50% diluted RTE respectively within 20 h by PUF-immobilized microbial consortium (
Figure 4B). However, decolorization at higher RTE concentration (75%) was strongly inhibited as only 56% AMDI removal was achieved within 20 h and a maximum of 62% in 48 h (data not shown). Since the textile effluent treatment goal is to remove dyes, the maximum ADMI removal value obtained in this experiment (92% ADMI removal at 50% RTE) by PUF-immobilized microbial consortium was within the acceptable limits (≤150 ADMI units) set for the final discharge of effluent to streams or water bodies [
50].
Figure 4.
Batch studies on decolorization of RTE by PUF-immobilized microbial consortium at 30 ± 0.2 °C. (A) Microaerophilic and aerobic incubation; and (B) different RTE concentrations v.z. 25%, 50% and 75%. Decolorization was measured in terms of percent ADMI removal. Data points indicate the mean of three independent replicates; ±SEM is indicated by error bars.
Figure 4.
Batch studies on decolorization of RTE by PUF-immobilized microbial consortium at 30 ± 0.2 °C. (A) Microaerophilic and aerobic incubation; and (B) different RTE concentrations v.z. 25%, 50% and 75%. Decolorization was measured in terms of percent ADMI removal. Data points indicate the mean of three independent replicates; ±SEM is indicated by error bars.
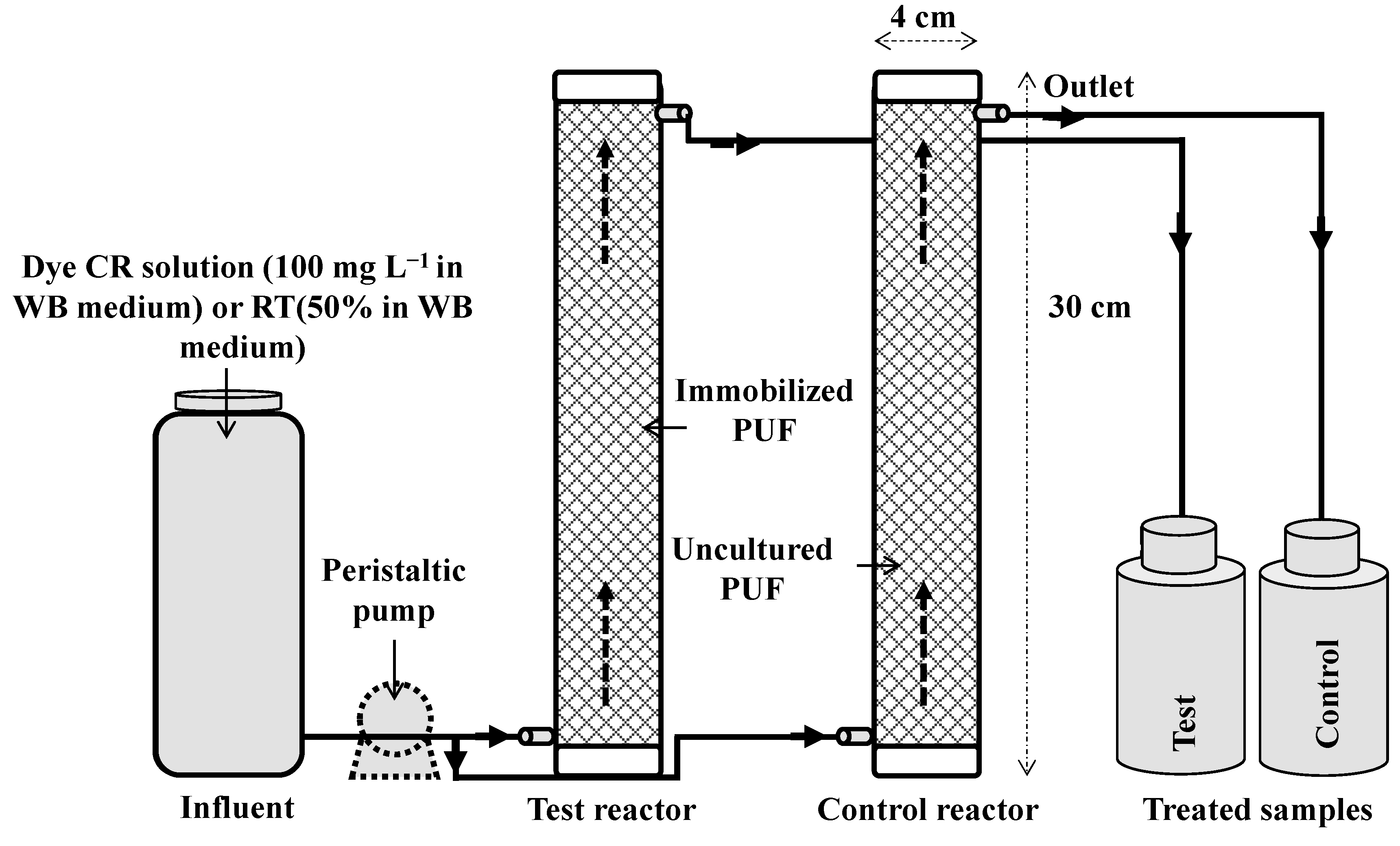
3.4. Upflow Column Bioreactor Study
To remove the dyes from a substantial amount of wastewater generated from the textile and dyestuff industries, there is a need for a continuous treatment process like immobilized microbial culture column reactors. Recently single culture immobilized bioreactors for the treatment of azo dyes and textile effluents under continuous operation using
Luffa cylindrica as the immobilization material were reported [
20,
53]. However, single culture could not remove the mixture of structurally different dyes presents in the textile effluents as compared to microbial consortia [
19], while, the mixed culture immobilized bioreactors are considered to represent a high rate continuous process suitable for the treatment of large amounts of dye-containing wastewaters [
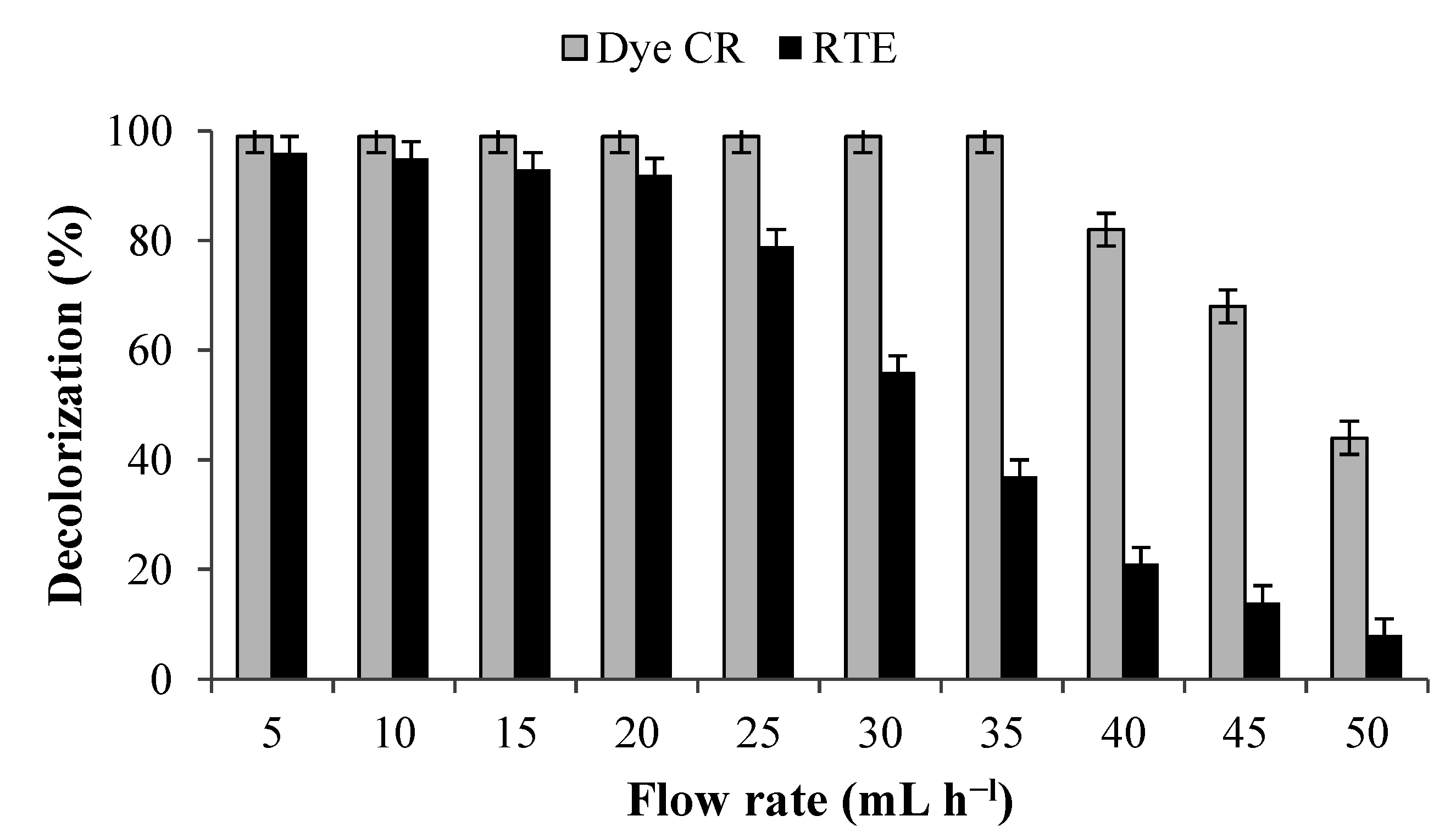
21]. Hence, in the present study a PUF-immobilized microbial consortium consisting of eight different bacteria has been investigated for the mineralization of azo dye and textile effluent under upflow column bioreactor conditions. The results of the decolorization of CR dye (100 mg·L
−1) and RTE (50% in WB medium) at different volumetric flow rates revealed that maximum dye removal was obtained at 35 mL·h
−l (HRT 5.7 h) and 20 mL·h
−l (HRT 10 h) respectively (
Figure 5). A further increase in flow rates beyond the optimum strongly inhibited the decolorization performance. The decolorization of both dye and textile effluent in a control reactor that contained uncultured PUF cubes was negligible and hence the dye removal with PUF immobilized microbial consortium was not because of adsorption by the PUF bed.
The analysis of TOC, COD and BOD of the dye and textile effluent before and after treatment was performed in order to confirm their mineralization. The dye decolorized at the optimized volumetric flow rate of 35 mL·h
−l and HRT 5.7 h showed 83%, 85% and 79% reduction in TOC, COD and BOD contents, respectively (
Table 2). In the case of RTE (50%) treatment, 92% ADMI removal was observed with 79%, 83% and 77% reduction in TOC, COD and BOD content at a flow rate of 20 mL·h
−l and HRT 10 h. The high COD removal efficiency suggested the potential of the developed upflow column bioreactor for the mineralization of azo dyes and real textile effluents. The performance of textile effluent treatment processes has been known to be directly associated with removal of COD [
54]. The enhanced reduction in TOC, COD and BOD contents indicates the efficient biodegradation of the CR dye and RTE in the column bioreactor by the immobilized microbial consortium.
Figure 5.
Decolorization of CR dye (100 mg·L−1) and RTE (50%) by PUF-immobilized microbial consortium under upflow column bioreactor conditions at various flow rates. Data points indicate the mean of three independent replicates; ±SEM is indicated by error bars.
Figure 5.
Decolorization of CR dye (100 mg·L−1) and RTE (50%) by PUF-immobilized microbial consortium under upflow column bioreactor conditions at various flow rates. Data points indicate the mean of three independent replicates; ±SEM is indicated by error bars.
Table 2.
Characterization of CR dye and RTE before and after treatment with PUF-mmobilized microbial consortium under upflow column bioreactor conditions.
Table 2.
Characterization of CR dye and RTE before and after treatment with PUF-mmobilized microbial consortium under upflow column bioreactor conditions.
| Parameters | Dye CR (100 mg·L−1) | RTE (50%) |
|---|
| Untreated | Treated (35 mL·h−l) | Untreated | Treated (20 mL·h−l) |
|---|
| TOC (mg·L−1) | 1750 ± 5.0 | 295 ± 3.0 | 1887 ± 6.0 | 390 ± 4.0 |
| COD (mg·L−1) | 1325 ± 3.0 | 195 ± 1.0 | 1620 ± 4.0 | 275 ± 2.0 |
| BOD (mg·L−1) | 210 ± 2.0 | 44 ± 1.0 | 338 ± 3.0 | 78 ± 1.0 |
| Color (%)/ADMI | 100 ± 0.0 | 1 ± 1.0 | 1943 ± 10.0 | 150 ± 2.0 |
3.5. Enzymatic Analysis in Free Microbial Consortium Culture
The potential of enzymes like laccases, tyrosinase, veratryl alcohol oxidase, azoreductase and NADH-DCIP reductase in the decolorization and degradation of dyes has been well documented [
55]. Several bacteria are known to possess various oxidative and reductive enzymes that can break down dyes into low molecular weight metabolites [
55]. In the present study, the oxidative enzymes laccase (275%), tryrosinase (110%) and veratryl alcohol oxidase (46%) were found to be induced significantly in CR dye (100 mg·L
−1) samples (12 h) decolorized by the developed microbial consortium when compared with control (
Table 3). It was also observed that the reductive enzymes azoreductase (700%) and DCIP reductase (329%) showed higher induction in decolorized samples. The significant increase in both oxidative and reductive enzyme activities in decolorized broth indicated their possible involvement in the metabolism of CR dye. The same observation was reported by Lade
et al. [
22] that increase in oxidoreductive enzyme activities in the microbial consortium culture was observed when exposed to the azo dye Trypan blue in microaerophilic conditions. In the case of RTE (50%), a significant induction in the activities of all studied enzymes
i.e., laccase (350%), tryrosinase (170%), veratryl alcohol oxidase (77%), azoreductase (850%) and DCIP reductase (376%) was observed after complete decolorization (20 h) by the microbial consortium. The overall enzymatic analysis results revealed that different sets of oxidoreductive enzymes were significantly induced in the microbial consortium when exposed to CR dye (100 mg·L
−1) and RTE (50%) which results in their decolorization. This is in agreement with the previous reports, where induction and involvement of the enzymes laccase, tyrosinase, veratryl alcohol oxidase, azoreductase and DCIP reductase in the metabolism of azo dyes by bacteria has been shown [
17,
34,
56,
57,
58].
Table 3.
Enzyme activities in microbial consortium control at 0 h (before addition of dye or textile effluent) and after CR dye (100 mg·L−1) and RTE (50%) additions.
Table 3.
Enzyme activities in microbial consortium control at 0 h (before addition of dye or textile effluent) and after CR dye (100 mg·L−1) and RTE (50%) additions.
| Enzymes | Control (0 h) | After Decolorization of CR Dye (12 h) | After Decolorization of RTE (20 h) |
|---|
| Laccase 1 | 0.4 ± 0.02 | 1.5 ± 0.02 *** | 1.8 ± 0.02 *** |
| Tyrosinase 2 | 1.0 ± 0.01 | 2.1 ± 0.03 ** | 2.7 ± 0.03 ** |
| Veratryl alcohol oxidase 3 | 1.3 ± 0.03 | 1.9 ± 0.04 ** | 2.3 ± 0.04 ** |
| Azoreductase 4 | 0.4 ± 0.02 | 3.2 ± 0.03 *** | 3.8 ± 0.03 *** |
| DCIP reductase 5 | 9.0 ± 1.6 | 38.6 ± 2.0 *** | 42.8 ± 2.2 *** |
3.6. Analysis of Dye CR Degradation
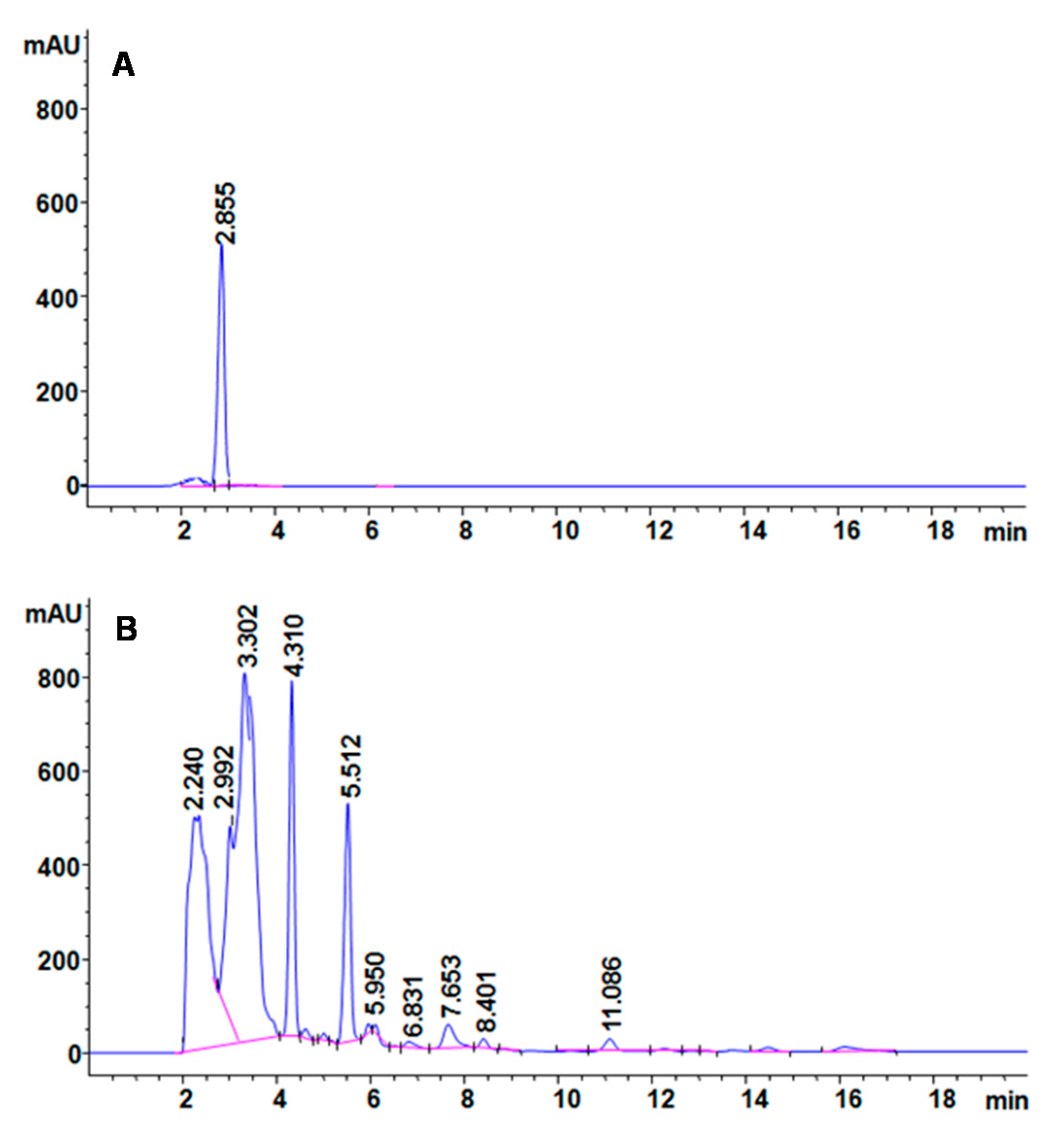
The mechanism behind decolorization of CR azo dye by the PUF-immobilized microbial consortium in an upflow column bioreactor was analyzed with different analytical techniques. The HPLC spectra of CR dye control showed a single peak at a retention time of 2.855 min (
Figure 6A). After treatment with PUF-immobilized microbial consortium in an upflow column bioreactor, the dye control peak disappeared and ten new peaks with different retention times of 2.240, 2.992, 3.302, 4.310, 5.512, 5.950, 6.831, 7.653, 8.401 and 11.086 min appeared (
Figure 6B). The disappearance of the control dye peak and appearance of several new peaks in the treated samples suggests the biotransformation of dye CR into different metabolites.
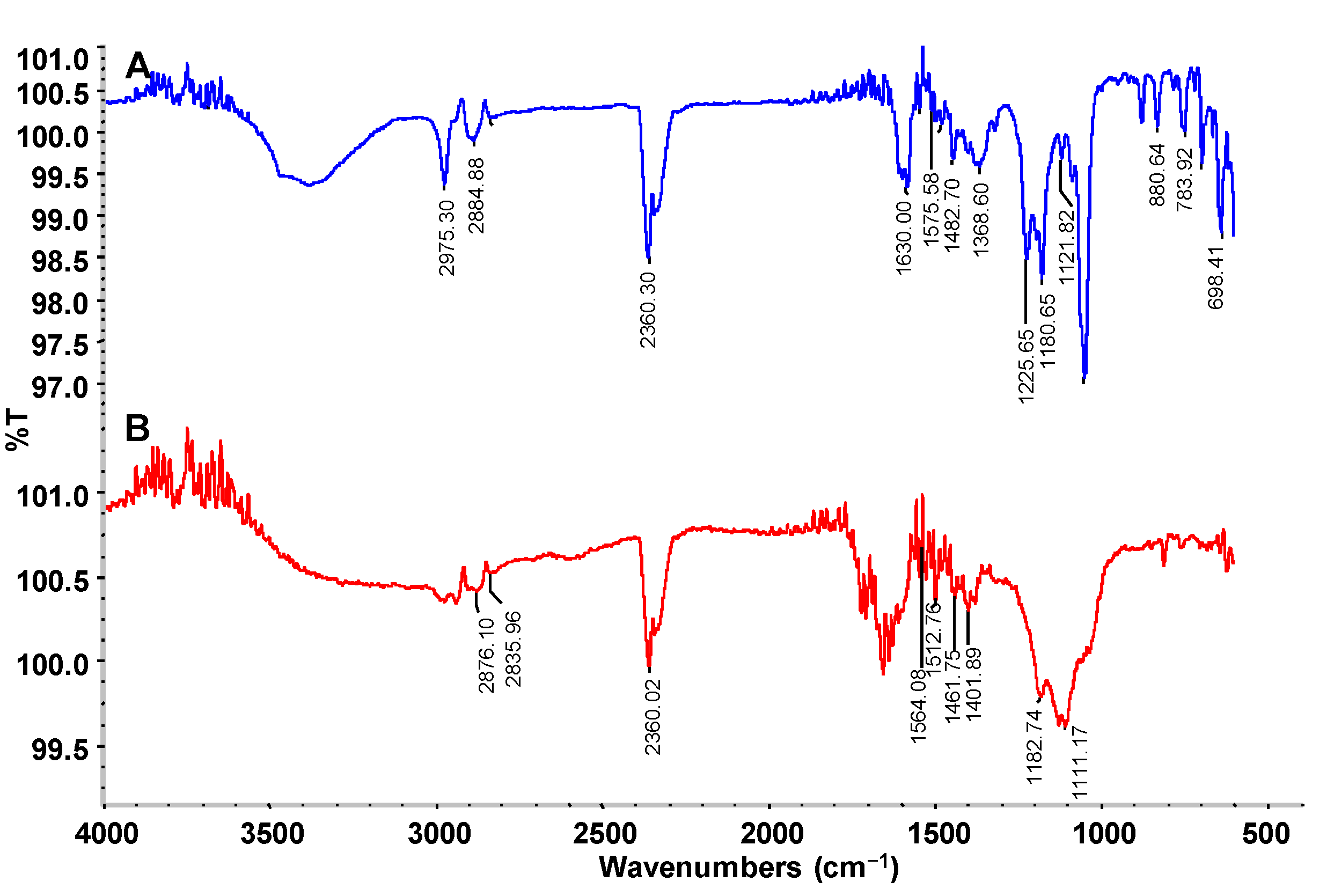
FTIR analysis of the dye control and its products after decolorization by the PUF-immobilized microbial consortium in an upflow column bioreactor was carried out to check the changes in functional groups. The FTIR spectral peaks 2975.30 and 2884.88 cm
−1 observed in the CR dye control represent C–H stretching as in alkanes (–CH
3–) while the 2360.30 cm
−1 band represents NH
+ stretching as in charged amines (C=NH
+) (
Figure 7A). The specific peaks at 1630.00 and 1575.58 cm
−1 indicate the presence of azo –N=N‒ double bond stretching as in azo compounds. In addition, the 1482.70, 1368.60, 1225.65, 1180.65 and 1121.82 peaks represent C–H deformation of alkanes, CH
3 deformation of tertiary butyl symmetric CH
3 bending, skeletal stretching of alkanes and C–N vibration of aliphatic amines, respectively. The 880.64 peak corresponds to the C–H deformation of a tetra or penta-substituted benzene containing one free hydrogen.
Figure 6.
HPLC profile of (A) CR dye and (B) its decolorized products obtained after treatment with PUF-immobilized microbial consortium in an upflow column bioreactor.
Figure 6.
HPLC profile of (A) CR dye and (B) its decolorized products obtained after treatment with PUF-immobilized microbial consortium in an upflow column bioreactor.
Figure 7.
FTIR profile of (A) CR dye and (B) its decolorized products obtained after treatment with PUF-immobilized microbial consortium in an upflow column bioreactor.
Figure 7.
FTIR profile of (A) CR dye and (B) its decolorized products obtained after treatment with PUF-immobilized microbial consortium in an upflow column bioreactor.
On the other hand, the FTIR spectra of the dye products decolorized by the microbial consortium showed a significantly different pattern of absorption peaks when compared to the parent CR dye. The specific azo bond peaks at1630.00 and 1575.58 cm
−1 disappeared after microbial consortium treatment, indicating the reductive breakdown of the CR azo dye (
Figure 7B). Additionally, the peaks from 880.64 to 698.41 cm
−1 representing the benzene ring structures of the parent CR dye also disappeared suggesting the cleavage of the dye molecule. Overall, the FTIR spectra of CR dye control differed significantly from the products decolorized by the microbial consortium in the upflow column bioreactor, supporting the biotransformation of the dye into different metabolites.
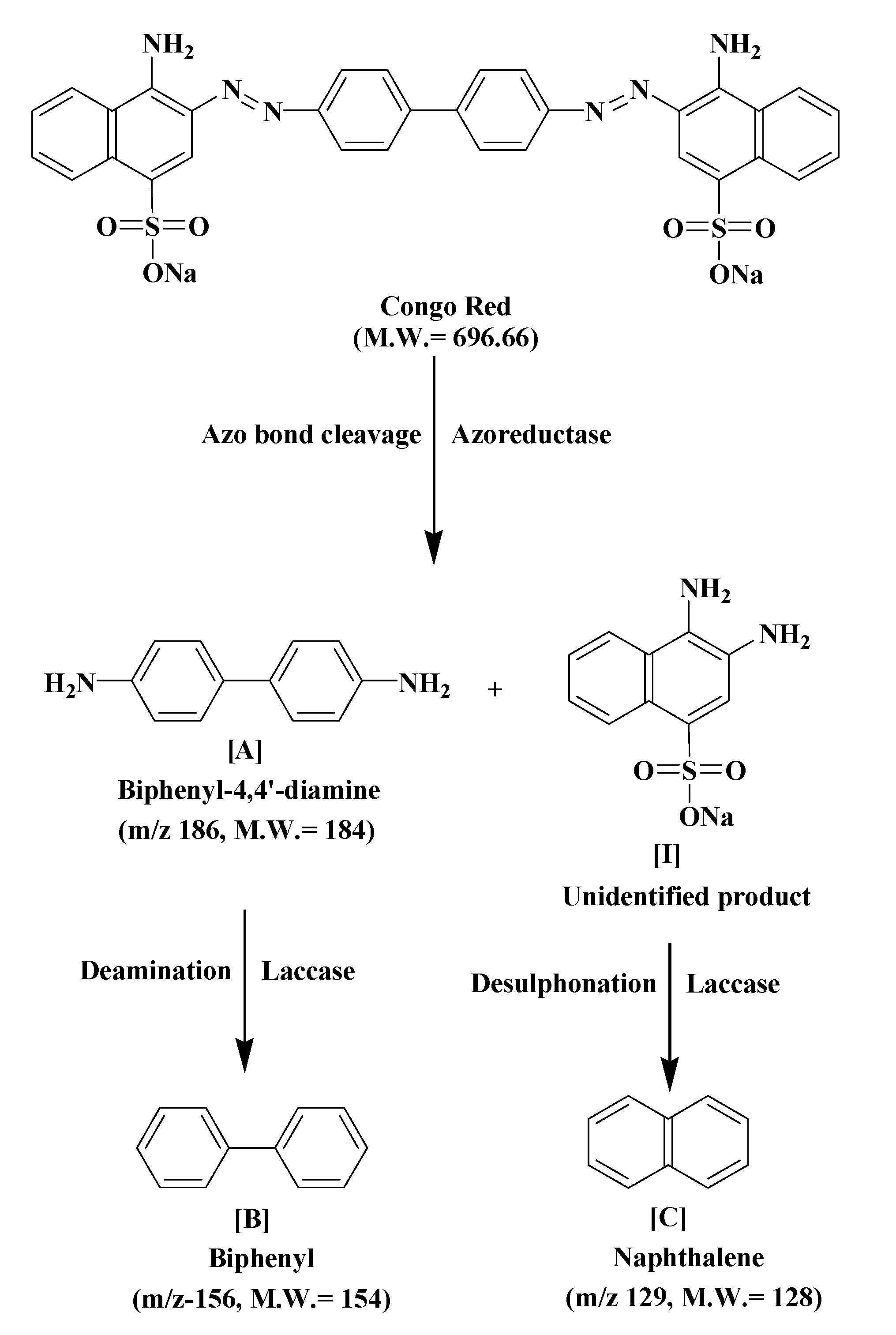
GC–MS analysis data was used to determine the probable metabolites produced and also to propose a degradation pathway of CR dye by PUF-immobilized microbial consortium. The structures of detected metabolites were assigned from the fragmentation pattern and
m/z values obtained (
Table 4). Result of the mass spectrum analysis revealed the presence of three intermediate CR dye degradation metabolites,
viz. biphenyl-1,4′-diamine, biphenyl and naphthalene.
Table 4.
GC–MS analysis data of degraded products of dye CR by PUF immobilized microbial consortium under upflow column bioreactor.
Figure 8 depicts the proposed biodegradation pathway of CR dye by PUF-immobilized microbial consortium in an upflow column bioreactor. The possible mineralization of CR dye starts with the initial conversion of the dye to form biphenyl-1,4′-diamine [A] and unidentified intermediate [I] catalyzed by azoreductase that cleaves the azo bond. The initial step in the microbial degradation of azo dyes is cleavage of electrophilic azo bonds which leads to their decolorization [
59]. This cleavage is known to form aromatic amines as end products; however most such amines further get degraded by the actions of oxidative enzymes [
60]. The disappearance of the FTIR spectral peaks at 1630.00 and 1575.58 cm
−1 in the treated dye metabolites support the azo bond cleavage of CR dye as shown in the proposed pathway.
Figure 8.
The proposed biodegradation pathway of CR dye by PUF-immobilized microbial consortium in an upflow column bioreactor.
Figure 8.
The proposed biodegradation pathway of CR dye by PUF-immobilized microbial consortium in an upflow column bioreactor.
It has been shown that a bacterial consortium consisting of
Providencia rettgeri strain HSL1 and
Pseudomonas sp. SUK1 performed the reduction of azo dyes under microaerophilic incubation conditions to produce toxic aromatic amines, but these disappeared in a subsequent aerobic process due to the activities of oxidative enzymes [
61]. Similarly, in the present study the metabolite [A] undergoes deamination by the oxidative enzyme laccase to form biphenyl [B], while intermediate [I] udergoes desulphonation, also by the laccase, to form the low molecular weight compound naphthalene [C] as final dye degradation products. The FTIR spectral peaks from 1564.08–1390 cm
−1 in the decolorized dye samples support the presence of naphthalene as shown in the proposed pathway. The naphthalene ring stretching vibrations are expected in the 1620–1390 cm
−1 infrared region [
62]. The oxidative cleavage of metabolite [B] and intermediate [I] are supposed to be caused by laccase activity, which is known to cleave the dye molecules asymmetrically [
63]. Additionally, considerable induction in the activities of tyrosinase and veratryl alcohol oxidase suggested their involvement in the oxidation of metabolite [A] and [I]. Lade
et al. during their study on biodegradation of the azo dye Trypan blue by an nriched microbial consortium reported that the dye was enzymatically degraded to form the low molecular weight compound naphthalene-1-ol as final intermediate metabolite [
22].
3.7. Toxicity Analysis
Although the present PUF-immobilized microbial consortium treatment in an upflow column reactor was efficient in the complete decolorization and degradation of the azo dye CR and RTE, it is essential to evaluate the toxicity of the treated samples. Treated dye wastewaters are mostly released to nearby farms for irrigation purposes and thus have a direct impact on the growth of crop plants as plant survival under dye stress is stunted. The evaluation of toxicity in terms of seed germination and plant survival is therefore extremely important for determining the suitability of treated wastewaters for agricultural applications. Phytotoxicity analysis of dye and textile effluent using seedlings of common agricultural crops has been suggested as the primary toxicity test to assess the toxic nature of dyes and their degradation metabolites [
64]. Results of the phytotoxicity test showed that the germination rate of seeds exposed to both untreated CR dye (100 mg·L
−1) and RTE (50%) was inhibited by 80% and 90% in
S. vulgare and 80% and 90% in
P. mungo, respectively (
Table 5). At the same time, less elongation in length of shoots
i.e., 4.1 and 4.2 cm for dye and 3.8 and 4.1 cm for textile effluent, as well as roots
i.e., 2.2 and 1.2 cm for dye and 2.0 and 0.9 cm for textile effluent, were also observed among both plants
S. vulgare and
P. mungo, respectively. The lower elongation in roots and shoots of both the plants exposed to untreated dye and textile effluent indicate a toxic effect on plant growth. In contrast, 100% germination of both the seeds with 11.2 and 6.7 cm of shoot and root length for
S. vulgare and 14.3 and 6.2 cm of shoot and root length for
P. mungo were found. It was also observed that seeds exposed to treated dye wastewaters showed similar germination as well as shoot and root lengths as like that of distilled water. These results clearly indicate that the treated dye and textile effluent samples were almost as non-toxic as distilled water. Moreover, the plants grown with treated samples were healthy in terms of shoot and root lengths, suggesting the conversion of complex dyes into simple oxidizable forms of a non-toxic nature.
Table 5.
Phytotoxicity analysis of control CR dye (100 mg·L−1), RTE (50%) and their treated samples using PUF-immobilized microbial consortium in an upflow column bioreactor.
Table 5.
Phytotoxicity analysis of control CR dye (100 mg·L−1), RTE (50%) and their treated samples using PUF-immobilized microbial consortium in an upflow column bioreactor.
| Samples | S. vulgare | P. mungo |
|---|
| Germination (%) | Shoot Length (cm) | Root Length (cm) | Germination (%) | Shoot Length (cm) | Root Length (cm) |
|---|
| Distilled water | 100 | 11.2 ± 0.3 | 6.7 ± 0.2 | 100 | 14.3 ± 0.3 | 6.2 ± 0.3 |
| CR (100 mg·L−1) | 20 | 4.1 ± 0.1 * | 2.2 ± 0.2 * | 20 | 4.2 ± 0.3 * | 1.2 ± 0.2 * |
| CR metabolites | 90 | 10.1 ± 0.2 $ | 5.6 ± 0.3 $ | 80 | 14.1 ± 0.2 | 5.9 ± 0.2 $ |
| RTE (50%) | 10 | 3.8 ± 0.2 * | 2.0 ± 0.2 * | 10 | 4.1 ± 0.2 * | 0.9 ± 0.1 * |
| RTE metabolites | 80 | 9.9 ± 0.2 $ | 5.4 ± 0.2 $ | 70 | 13.9 ± 0.2 | 5.5 ± 0.2 $ |
Acute toxicity testing has been described for the intoxication of mammals by chemical consumption. The mortality test with aquatic crustacean
D. magna is the most common and rapid acute toxicity assay used to assess the toxic effect of chemicals on mammals, including human beings [
65]. Results of the acute toxicity assay showed complete mortality of
D. magna exposed to CR dye (100 mg·L
−1) solution, as well as RTE (50%), whereas only 10% and 20% mortality was observed in treated dye and textile effluent samples, respectively (
Table 6). Similar results were reported by Lade
et al., where complete mortality of
D. magna was observed in the presence of untreated azo dyes [
61]. The complete mortality of
D. magna in untreated dye and textile effluent indicates their toxic nature, while treated samples showed greater crustacean survival. The results of both the phytotoxicity and acute toxicity assays with dye and textile effluent thus confirm the less toxic nature of the treated samples. The detoxification of dye and textile effluent might be due to removal of color and reduced COD levels. This clearly suggests the possible use of PUF-immobilized microbial consortium upflow column bioreactors for the mineralization and detoxification of azo dyes.
Table 6.
Mortality of D. magna exposed to CR dye (100 mg·L−1), RTE (50%) and their culture supernatants obtained after treatment with PUF immobilized microbial consortium in an upflow column bioreactor.
Table 6.
Mortality of D. magna exposed to CR dye (100 mg·L−1), RTE (50%) and their culture supernatants obtained after treatment with PUF immobilized microbial consortium in an upflow column bioreactor.
| Samples | Mortality (%) |
|---|
| Distilled water | 0 ± 0.0 |
| CR (100 mg·L−1) | 100 ± 0.0 |
| CR treated broth supernatant | 10 ± 0.0 |
| RTE (50%) | 100 ± 0.0 |
| RTE treated broth supernatant | 20 ± 0.0 |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






