
Characterization of the Bacterial Community Naturally Present on Commercially Grown Basil Leaves: Evaluation of Sample Preparation Prior to Culture-Independent Techniques

Abstract
:1. Introduction
2. Experimental Section
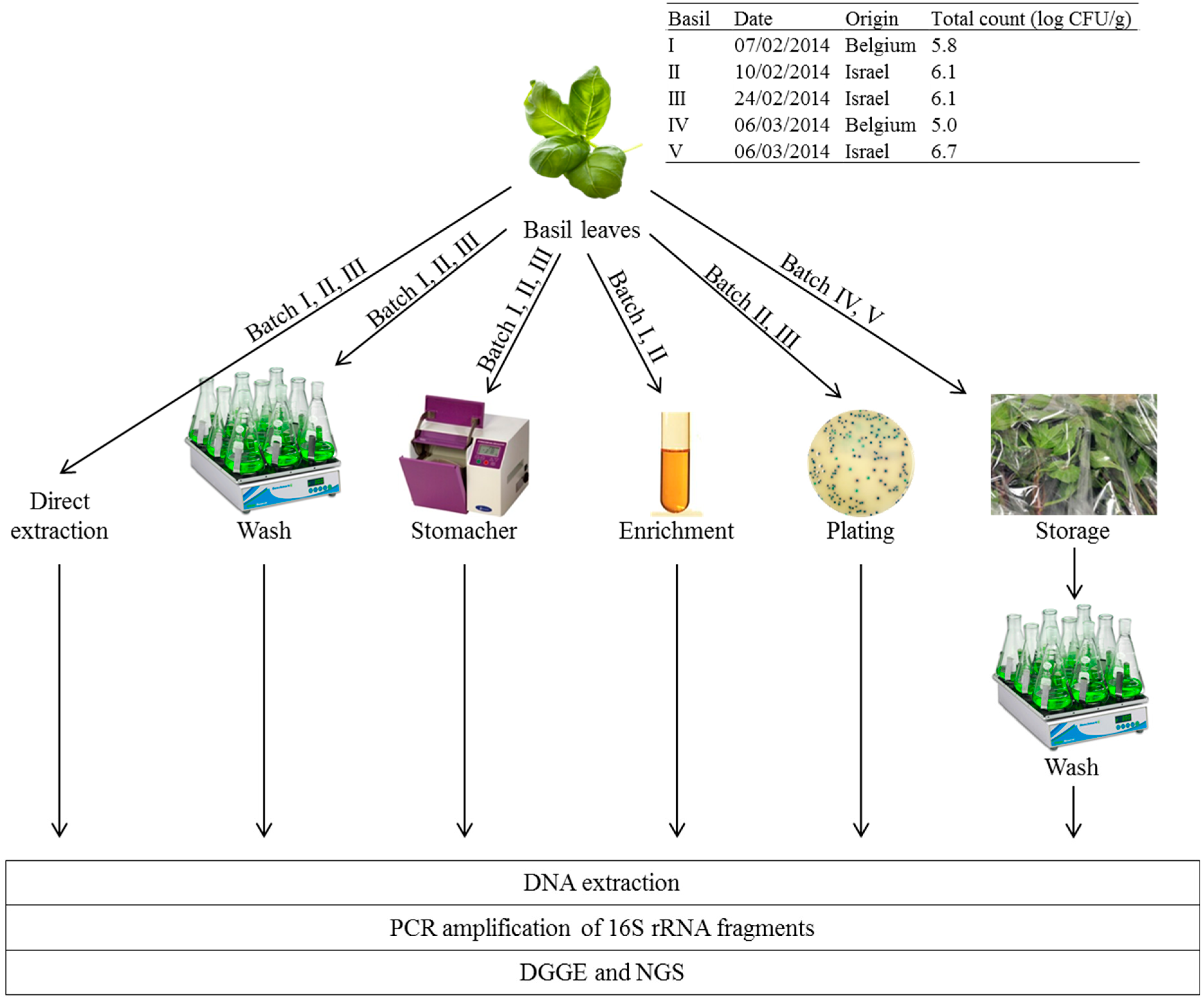
2.1. Basil
2.2. Molecular Microbiological Analyses
2.2.1. Sample Preparation

2.2.2. DNA Extraction
2.2.3. PCR Amplification of 16S rRNA Gene
2.2.4. Denaturing Gradient Gel Electrophoresis (DGGE)
2.2.5. Next-Generation Sequencing (NGS)
2.3. Culture-Based Microbiological Analyses
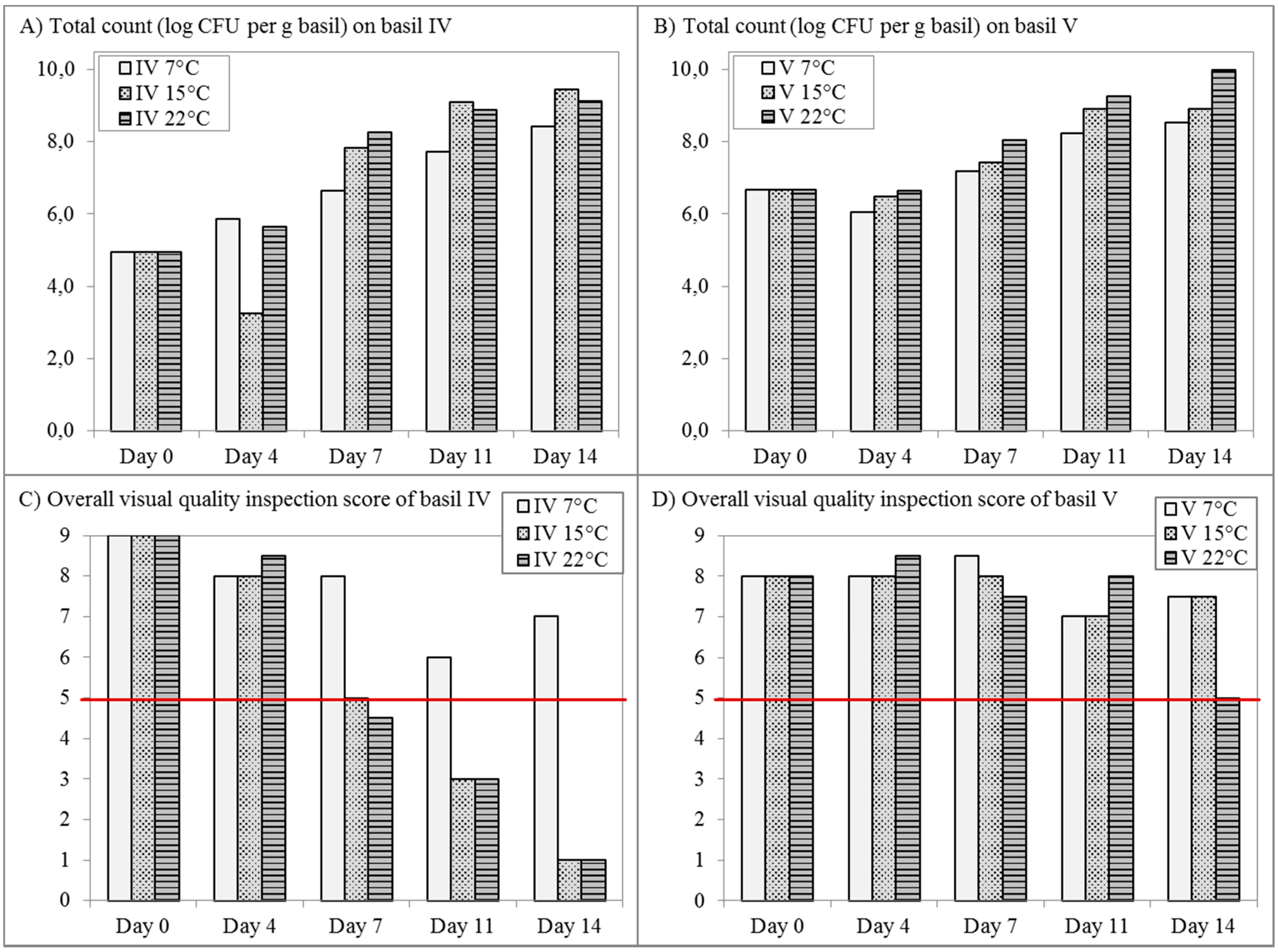
2.4. Storage Experiments
3. Results
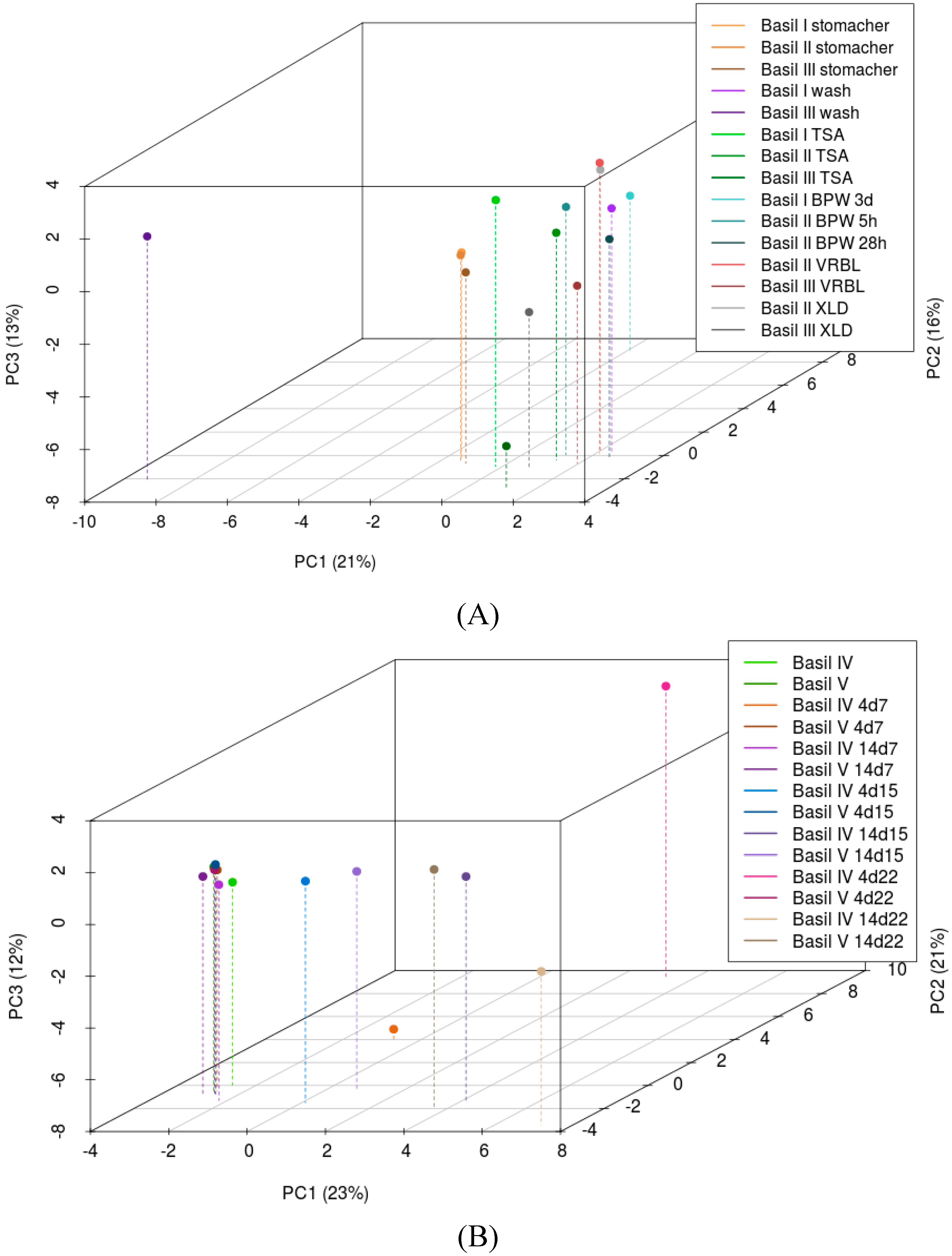
3.1. Culture-Independent Characterisation of the Bacterial Community


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Basil I, Stomacher | Basil I, Wash | Basil II, Stomacher | Basil II, Wash | Basil III, Stomacher | Basil III, Wash |
|---|---|---|---|---|---|---|
| Total number of reads | 36,887 | 40,630 | 33,794 | Failed | 64,772 | 10,772 |
| Median length of reads (bp) | 473 | 474 | 473 | 473 | 473 | |
| Not assigned | 29 | 694 | 51 | 20 | 6 | |
| Eukaryota | 33,809 | 22,490 | 28,242 | 63,837 | 10,202 | |
| Bacteria (rescaled to 100% below) | 3,050 | 17,446 | 5,501 | 915 | 564 | |
| Bacteroidetes | 1% | 1% | 74% | |||
| Arcicella | 1% | |||||
| Chryseobacterium | 1% | 5% | ||||
| Flavobacterium | 11% | |||||
| Sphingobacterium | 56% | |||||
| Alphaproteobacteria | 87% | 5% | 83% | 71% | 17% | |
| Altererythrobacter | 2% | |||||
| Novosphingobium | 86% | 4% | 81% | 71% | 9% | |
| Sphingobium | 1% | |||||
| Sphingomonas | 1% | 2% | ||||
| Betaproteobacteria | 1% | 6% | 1% | |||
| Herbaspirillum | 4% | |||||
| Gammaproteobacteria | 10% | 88% | 16% | 26% | 5% | |
| Acinetobacter | 4% | 18% | 5% | |||
| Pseudomonas | 1% | 40% | 4% | 5% | ||
| Rheinheimera | 1% | |||||
| Enterobacteriaceae | 8% | 43% | 11% | 2% | ||
| Enterobacter | 15% | 2% | ||||
| Erwinia | 1% | 2% | ||||
| Klebsiella | 3% | 11% | 1% | |||
| Kluyvera | 1% | |||||
| Pantoea | 6% | 6% | ||||
| Rahnella | 4% | |||||
| Raoultella | 2% | 2% | ||||
| Unclassified | 1% | 1% | 1% | 2% |

3.2. Culturing of the Bacterial Community
3.3. The Bacterial Community on Basil Leaves throughout Storage and Spoilage
| Sample | Basil I, BPW 3d | Basil II, BPW 5h | Basil II, BPW 28h | Basil I, TSA | Basil II, TSA | Basil III, TSA | Basil II, VRBL | Basil III, VRBL | Basil II, XLD | Basil III, XLD |
|---|---|---|---|---|---|---|---|---|---|---|
| Total number of reads | 42,703 | 29,757 | 36,870 | 13,533 | 47,182 | 39,389 | 23,565 | 38,841 | 40,201 | 31,964 |
| Median length of reads (bp) | 509 | 494 | 500 | 503 | 502 | 496 | 500 | 496 | 499 | 503 |
| Not assigned | 1,829 | 1,985 | 2,294 | 299 | 3,839 | 378 | 114 | 712 | 226 | 198 |
| Eukaryota | 0 | 14,201 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bacteria (rescaled to 100% below) | 40,874 | 13,572 | 34,576 | 13,234 | 43,343 | 39,011 | 23,451 | 38,129 | 39,975 | 31,766 |
| Actinobacteria | 12% | |||||||||
| Arthrobacter | 8% | |||||||||
| Kocuria | 2% | |||||||||
| Bacteroidetes | 55% | 4% | ||||||||
| Bacteroides | 42% | |||||||||
| Macellibacteroides | 3% | |||||||||
| Parabacteroides | 11% | |||||||||
| Chryseobacterium | 3% | |||||||||
| Clostridia | 6% | 1% | ||||||||
| Clostridium | 5% | |||||||||
| Alphaproteobacteria | 5% | |||||||||
| Novosphingobium | 5% | |||||||||
| Betaproteobacteria | 6% | 3% | 2% | |||||||
| Comamonas | 5% | 2% | 2% | |||||||
| Gammaproteobacteria | 38% | 95% | 93% | 83% | 100% | 97% | 100% | 98% | 100% | 100% |
| Aeromonas | 38% | 31% | 36% | |||||||
| Alishewanella | 6% | |||||||||
| Shewanella | 1% | 1% | ||||||||
| Rheinheimera | 13% | |||||||||
| Acinetobacter | 4% | 1% | 45% | 17% | 6% | |||||
| Pseudomonas | 1% | 37% | 14% | 16% | 13% | 6% | 23% | 1% | 55% | |
| Stenotrophomonas | 1% | 1% | ||||||||
| Enterobacteriaceae | 34% | 94% | 55% | 23% | 84% | 10% | 92% | 36% | 99% | 7% |
| Aranicola | 2% | |||||||||
| Cedecea | 2% | |||||||||
| Citrobacter | 17% | 1% | 1% | 2% | ||||||
| Enterobacter | 8% | 10% | 15% | 5% | 5% | 1% | 11% | 16% | 11% | 3% |
| Erwinia | 10% | 5% | 6% | 13% | 28% | 28% | ||||
| Klebsiella | 3% | 1% | 7% | 4% | 5% | 7% | 6% | |||
| Kluyvera | 1% | 1% | 1% | 1% | 4% | 6% | ||||
| Pantoea | 2% | 70% | 21% | 3% | 61% | 1% | 41% | 7% | 45% | 1% |
| Pectobacterium | 3% | 7% | ||||||||
| Raoultella | 1% | 2% |


| Sample | Basil IV Day0 | Basil IV 7d 4°C | Basil IV 7d 14°C | Basil IV 15d 4°C | Basil IV 15d 14°C | Basil IV 22d 4°C | Basil IV 22d 14°C | Basil V Day0 | Basil V 7d 4°C | Basil V 7d 14°C | Basil V 15d 4°C | Basil V 15d 14°C | Basil V 22d 4°C | Basil V 22d 14°C |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total number of reads | 36.746 | 35.279 | 63.993 | 11.892 | 24.615 | 49.852 | 49.347 | 60.524 | 57.824 | 83.574 | 37.559 | 51.080 | 70.829 | 64.761 |
| Median length of reads (bp) | 438 | 456 | 404 | 437 | 405 | 464 | 393 | 417 | 459 | 402 | 437 | 457 | 453 | 393 |
| Not assigned | 56 | 40 | 354 | 109 | 788 | 32 | 700 | 94 | 90 | 928 | 181 | 151 | 208 | 3.815 |
| Eukaryota | 35.626 | 34.556 | 26.747 | 9.870 | 51 | 49.363 | 222 | 46.475 | 53.834 | 1.735 | 27.767 | 48.859 | 56.755 | 9.132 |
| Bacteria (rescaled to 100 % below) | 1.065 | 682 | 36.892 | 1.913 | 23.776 | 457 | 48.425 | 13.954 | 3.900 | 80.911 | 9.611 | 2.070 | 13.867 | 51.815 |
| Actinobacteria | 14% | 1% | 1% | 14% | 1% | |||||||||
| Microbacterium | 2% | 1% | ||||||||||||
| Arthrobacter | 10% | 1% | 8% | |||||||||||
| Bacteroidetes | 4% | 2% | 17% | 29% | 18% | 14% | 16% | 19% | ||||||
| Chryseobacterium | 2% | 1% | 11% | 3% | 1% | |||||||||
| Flavobacterium | 2% | 17% | 25% | 1% | 3% | |||||||||
| Pedobacter | 2% | 18% | 2% | |||||||||||
| Sphingobacterium | 9% | 18% | ||||||||||||
| Bacilli | 2% | 1% | ||||||||||||
| Bacillus | 2% | |||||||||||||
| Clostridia | 3% | |||||||||||||
| Clostridium | 2% | |||||||||||||
| Alphaproteobacteria | 5% | 20% | 8% | 11% | 21% | 8% | 2% | 8% | 2% | 23% | 4% | 8% | ||
| Methylobacterium | 9% | 1% | ||||||||||||
| Agrobacterium | 1% | 1% | 8% | 1% | ||||||||||
| Rhizobium | 2% | 2% | 1% | 2% | 2% | |||||||||
| Novosphingobium | 5% | 19% | 4% | 6% | 8% | 3% | 2% | 7% | 1% | 11% | 3% | 3% | ||
| Sphingomonas | 1% | 1% | 1% | 1% | 1% | |||||||||
| Betaproteobacteria | 28% | 15% | 2% | 9% | 15% | 36% | 20% | 6% | 28% | |||||
| Achromobacter | 3% | 1% | 1% | |||||||||||
| Burkholderia | 2% | 1% | 1% | |||||||||||
| Acidovorax | 5% | |||||||||||||
| Comamonas | 1% | 1% | ||||||||||||
| Delftia | 1% | 8% | ||||||||||||
| Variovorax | 2% | |||||||||||||
| Duganella | 1% | 3% | ||||||||||||
| Herbaspirillum | 5% | 2% | 6% | |||||||||||
| Herminiimonas | 1% | |||||||||||||
| Janthinobacterium | 17% | 1% | 2% | 2% | ||||||||||
| Oxalicibacterium | 1% | 2% | 5% | |||||||||||
| Methylobacillus | 12% | |||||||||||||
| Methylophilus | 2% | 4% | 32% | 4% | ||||||||||
| Zoogloea | 2% | |||||||||||||
| Gammaproteobacteria | 66% | 44% | 94% | 64% | 44% | 7% | 54% | 97% | 91% | 100% | 97% | 54% | 95% | 45% |
| Pseudoalteromonas | 3% | 1% | ||||||||||||
| Acinetobacter | 4% | |||||||||||||
| Pseudomonas | 56% | 30% | 71% | 42% | 10% | 3% | 22% | 96% | 85% | 95% | 95% | 44% | 89% | 24% |
| Luteibacter | 1% | 6% | 1% | |||||||||||
| Stenotrophomonas | 1% | 22% | 2% | 17% | 4% | 12% | ||||||||
| Xanthomonas | 1% | 2% | 1% | |||||||||||
| Enterobacteriaceae | 8% | 7% | 22% | 21% | 5% | 10% | 1% | 6% | 3% | 2% | 5% | 5% | 8% | |
| Buchnera | 2% | |||||||||||||
| Enterobacter | 3% | 3% | 2% | 7% | 1% | 6% | ||||||||
| Erwinia | 3% | |||||||||||||
| Ewingella | 4% | 11% | ||||||||||||
| Pantoea | 4% | 1% | 1% | 5% | 2% | 1% | 5% | 3% | 1% | |||||
| Rahnella | 4% | 10% | 5% | 1% | 1% | |||||||||
| Serratia | 5% | 1% | ||||||||||||
| Unclassified | 2% |
4. Discussion
| Genus | Gram Staining | Respiratory Metabolism | Motility | Temperature Range Growth | Habitat | Pathogenicity |
|---|---|---|---|---|---|---|
| Flavobacterium | Gram negative | Aerobic | Nonmotile or motile by gliding | −7 to 45 °C | Soil, freshwater, marine and saline environments | Some species, such as F. columnare, F. psychrophilum and F. branchiophilum, are pathogenic for freshwater fish. Some strains of F. johnsoniae are plant pathogens causing soft rot in various plants. |
| Sphingobacterium | Gram negative | Aerobic | Sliding motility | 2 to 45 °C | Soil and composted manure | Some species are opportunistic pathogens for humans. |
| Acinetobacter | Gram negative | Aerobic | Twitching motility by fimbriae | 20 to 37 °C | Soil, water, sewage and plants | Although considered normally nonpathogenic, they may cause nosocomial infections such as bacteremia, secondary meningitis, pneumonia, and urinary tract infections in humans. |
| Pseudomonas | Gram negative | Aerobic | Motile by one or several polar flagella and fimbriae | 4 to 45 °C | Plants (rhizospheres and leave surfaces) and soil | Some species are pathogenic for humans, animals, or plants. Plant pathogenic species such as P. syringae may cause tumorous outgrowth, rot, blight or chlorosis, and necrosis in plants due to secretion of substances (such as toxins, plant hormones and enzymes) which alter the normal metabolism of plant cells. Others are opportunistic pathogens for animals and humans, such as P. aeruginosa. |
| Citrobacter | Gram negative | Facultatively anaerobic | Usually motile by peritrichous flagella | 5 to >37 °C | Intestinal tract of humans and some animals, soil, water, sewage, plants and food (vegetables, dairy, fish) | Some species are opportunistic pathogens for humans. |
| Enterobacter | Gram negative | Facultatively anaerobic | Motile by peritrichous flagella | 4 to 44 °C | Plants (rhizophere and leaves) and the intestinal tract of humans and animals | Some species are plant pathogens, such as E. nimipressuralis (wetwood in elm trees), E. cancerogenus (canker disease of Populus species) and E. pyrinus (brown leaf spot disease in pears). |
| Erwinia | Gram negative | Facultatively anaerobic | Motile by peritrichous flagella | 0 to 40 °C | Plants | Plant pathogens which cause mainly blights and wilts. Infection through natural openings and wounds, followed by spread through the vascular tissue. |
| Klebsiella | Gram negative | Facultatively anaerobic | Nonmotile (except K. mobilis) | 5 to 45 °C | Intestinal tract of humans and animals, soil, water, sewage and plants | Opportunistic and nosocomial human pathogens, e.g. K. pneumoniae, causing pneumonia, urinary tract infections, bacteremia and sepsis. |
| Kluyvera | Gram negative | Facultatively anaerobic | Motile | 4 to 40 °C | Intestinal tract of humans and animals, soil, sewage and food (milk, dairy and other food products of animal origin) | Opportunistic human pathogen. |
| Pantoea | Gram negative | Facultatively anaerobic | Most strains are motile by peritrichous flagella | 4 to 41 °C | Plants, seeds, fruits, soil and water | Some strains are opportunistic pathogens for plants, humans and animals. |
| Rahnella | Gram negative | Facultatively anaerobic | Motile by peritrichous flagella | 1 to ≥37 °C | Fresh water, soil, plant rhizosphere, intestinal tract of snails | Opportunistic human pathogens causing wound infections, bacteremias, acute gastroenteritis and septicemia. |
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Ann. Rev. Plant. Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed]
- Leveau, J.H.J.; Lindow, S.E. Appetite of an epiphyte: Quantitative monitoring of bacterial sugar consumption in the phyllosphere. Proc. Natl. Acad. Sci. USA 2001, 98, 3446–3453. [Google Scholar] [CrossRef] [PubMed]
- Axtell, C.A.; Beattie, G.A. Construction and characterization of a proU-gfp transcriptional fusion that measures water availability in a microbial habitat. Appl. Environ. Microbiol. 2002, 68, 4604–4612. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, K.M.; Ji, P.S.; Firestone, M.K.; Lindow, S.E. Two novel bacterial biosensors for detection of nitrate availability in the rhizosphere. Appl. Environ. Microbiol. 2005, 71, 8537–8547. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.L.; Carroll, T.L.; Sundin, G.W. The role of pigmentation, ultraviolet radiation tolerance, and leaf colonization strategies in the epiphytic survival of phyllosphere bacteria. Microb. Ecol. 2005, 49, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Velasco, G.; Tydings, H.A.; Boyer, R.R.; Falkinham, J.O.; Ponder, M.A. Characterization of interactions between Escherichia coli O157:H7 with epiphytic bacteria in vitro and on spinach leaf surfaces. Int. J. Food Microbiol. 2012, 153, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.M.; Butterfield, J.E. Salmonella contamination associated with bacterial soft rot of fresh fruits and vegetables in the marketplace. Plant. Dis. 1997, 81, 867–872. [Google Scholar] [CrossRef]
- Allen, K.J.; Kovacevic, J.; Cancarevic, A.; Wood, J.; Xu, J.Q.; Gill, B.; Allen, J.K.; Mesak, L.R. Microbiological survey of imported produce available at retail across Canada. Int. J. Food Microbiol. 2013, 162, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Jacxsens, L.; Ibanez, I.C.; Gomez-Lopez, V.M.; Fernandes, J.A.; Allende, A.; Uyttendaele, M.; Huybrechts, I. Belgian and spanish consumption data and consumer handling practices for fresh fruits and vegetables useful for further microbiological and chemical exposure assessment. J. Food Prot. 2015, 78, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, M.H.; Blomhoff, R.; Andersen, L.F. Intakes of culinary herbs and spices from a food frequency questionnaire evaluated against 28-days estimated records. Nutr. J. 2011, 10. [Google Scholar] [CrossRef] [PubMed]
- International Commission on Microbiological Specifications for Foods (ICMSF). Chapter 5. Vegetables and Vegetable Products, Introduction. In Microorganisms in Foods 6: Microbial Ecology of Food Commodities, 2nd ed.; Chapman & Hall: New York, NY, USA, 2005; pp. 277–325. [Google Scholar]
- Lahou, E.; Jacxsens, L.; Van Landeghem, F.; Uyttendaele, M. Microbiological sampling plan based on risk classification to verify supplier selection and production of served meals in food service operation. Food Microbiol. 2014, 41, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Handelsman, J. Metagenomics: Application of genomics to uncultured microorganisms. Microbiol. Mol. Biol. Rev. 2004, 68, 669–685. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [PubMed]
- Muyzer, G.; Dewaal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Ercolini, D. PCR-DGGE fingerprinting: Novel strategies for detection of microbes in food. J. Microbiol. Meth. 2004, 56, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Mertz, A.W.; Koo, O.K.; O'Bryan, C.A.; Morawicki, R.; Sirsat, S.A.; Neal, J.A.; Crandall, P.G.; Ricke, S.C. Microbial ecology of meat slicers as determined by denaturing gradient gel electrophoresis. Food Control. 2014, 42, 242–247. [Google Scholar] [CrossRef]
- Chen, H.B.; Liu, Z.Y.; Wang, M.Y.; Chen, S.J.; Chen, T.W. Characterisation of the spoilage bacterial microbiota in oyster gills during storage at different temperatures. J. Sci. Food Agric. 2013, 93, 3748–3754. [Google Scholar] [CrossRef] [PubMed]
- Alfonzo, A.; Ventimiglia, G.; Corona, O.; Di Gerlando, R.; Gaglio, R.; Francesca, N.; Moschetti, G.; Settanni, L. Diversity and technological potential of lactic acid bacteria of wheat flours. Food Microbiol. 2013, 36, 343–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, G.V.D.; Magalhaes-Guedes, K.T.; Schwan, R.F. Rdna-based dgge analysis and electron microscopic observation of cocoa beans to monitor microbial diversity and distribution during the fermentation process. Food Res. Int. 2013, 53, 482–486. [Google Scholar] [CrossRef]
- Johansen, P.; Vindelov, J.; Arneborg, N.; Brockmann, E. Development of quantitative PCR and metagenomics-based approaches for strain quantification of a defined mixed-strain starter culture. Syst. Appl. Microbiol. 2014, 37, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Solieri, L.; Dakal, T.C.; Giudici, P. Next-generation sequencing and its potential impact on food microbial genomics. Ann. Microbiol. 2013, 63, 21–37. [Google Scholar] [CrossRef]
- Diaz-Sanchez, S.; Hanning, I.; Pendleton, S.; D'Souza, D. Next-generation sequencing: The future of molecular genetics in poultry production and food safety. Poultry Sci. 2013, 92, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Lee, S.Y.; An, C.M.; Kang, J.H.; Kim, J.H.; Chai, J.C.; Chen, J.; Kang, J.S.; Ahn, J.J.; Lee, Y.S.; et al. Comparative study between next generation sequencing technique and identification of microarray for species identification within blended food products. Biochip. J. 2012, 6, 354–361. [Google Scholar] [CrossRef]
- Naccache, S.N.; Federman, S.; Veeraraghavan, N.; Zaharia, M.; Lee, D.; Samayoa, E.; Bouquet, J.; Greninger, A.L.; Luk, K.C.; Enge, B.; et al. A cloud-compatible bioinformatics pipeline for ultrarapid pathogen identification from next-generation sequencing of clinical samples. Genome Res. 2014, 24, 1180–1192. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, R.; Mishra, A.K.; Raoult, D.; Fournier, P.E. Genomics and metagenomics in medical microbiology. J. Microbiol. Meth. 2013, 95, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16s ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Boon, N.; Top, E.M.; Verstraete, W.; Siciliano, S.D. Bioaugmentation as a tool to protect the structure and function of an activated-sludge microbial community against a 3-chloroaniline shock load. Appl. Environ. Microbiol. 2003, 69, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Cantwell, H.I.; Reid, M.S. Postharvest physiology and handling of fresh culinary herbs. J. Herbs Spices Med. Plants 1993, 1, 93–125. [Google Scholar] [CrossRef]
- Kampfer, P.; Young, C.C.; Busse, H.J.; Lin, S.Y.; Rekha, P.D.; Arun, A.B.; Chen, W.M.; Shen, F.T.; Wu, Y.H. Novosphingobium soli sp. nov., isolated from soil. Int. J. Syst. Evol. Micr. 2011, 61, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, H.; Kamesaki, T.; Sasaki, R.; Minami, H.; Yumoto, I. Production of two types of exopolysaccharide by Novosphingobium rosa. J. Biosci. Bioeng. 2003, 95, 152–156. [Google Scholar] [CrossRef]
- Addison, S.L.; Foote, S.M.; Reid, N.M.; Lloyd-Jones, G. Novosphingobium nitrogenifigens sp nov., a polyhydroxyalkanoate-accumulating diazotroph isolated from a New Zealand pulp and paper wastewater. Int. J. Syst. Evol. Micr. 2007, 57, 2467–2471. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.P.; Wang, B.J.; Liu, Y.H.; Liu, S.J. Novosphingobium taihuense sp nov., a novel aromatic-compound-degrading bacterium isolated from Taihu lake, China. Int. J. Syst. Evol. Micr. 2005, 55, 1229–1232. [Google Scholar] [CrossRef] [PubMed]
- Kampfer, P.; Witzenberger, R.; Denner, E.B.M.; Busse, H.J.; Neef, A. Novosphingobium hassiacum sp nov., a new species isolated from an aerated sewage pond. Syst. Appl. Microbiol. 2002, 25, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Tiirola, M.A.; Busse, H.J.; Kampfer, P.; Mannisto, M.K. Novosphingobium lentum sp nov., a psychrotolerant bacterium from a polychlorophenol bioremediation process. Int. J. Syst. Evol. Micr. 2005, 55, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Aylward, F.O.; McDonald, B.R.; Adams, S.M.; Valenzuela, A.; Schmidt, R.A.; Goodwin, L.A.; Woyke, T.; Currie, C.R.; Suen, G.; Poulsen, M. Comparison of 26 sphingomonad genomes reveals diverse environmental adaptations and biodegradative capabilities. Appl. Environ. Microb. 2013, 79, 3724–3733. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Chai, L.Y.; Zhu, Y.H.; Yang, Z.H.; Zheng, Y.; Zhang, H. Biodegradation of kraft lignin by a bacterial strain Comamonas sp b-9 isolated from eroded bamboo slips. J. Appl. Microbiol. 2012, 112, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Sultana, T.; Joe, M.M.; Yim, W.; Cho, J.C.; Sa, T. Nitrogen-fixing bacteria with multiple plant growth-promoting activities enhance growth of tomato and red pepper. J. Basic Microb. 2013, 53, 1004–1015. [Google Scholar] [CrossRef] [PubMed]
- Opalchenova, G.; Obreshkova, D. Comparative studies on the activity of basil—an essential oil from Ocimum basilicum L.—against multidrug resistant clinical isolates of the genera Staphylococcus, Enterococcus and Pseudomonas by using different test methods. J. Microbiol. Meth. 2003, 54, 105–110. [Google Scholar] [CrossRef]
- Pezzoli, L.; Elson, R.; Little, C.L.; Yip, H.; Fisher, I.; Yishai, R.; Anis, E.; Valinsky, L.; Biggerstaff, M.; Patel, N.; et al. Packed with salmonella - investigation of an international outbreak of Salmonella Senftenberg infection linked to contamination of prepacked basil in 2007. Foodborne Pathog. Dis. 2008, 5, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Kisluk, G.; Kalily, E.; Yaron, S. Resistance to essential oils affects survival of Salmonella enterica serovars in growing and harvested basil. Environ. Microbiol. 2013, 15, 2787–2798. [Google Scholar] [PubMed]
- Settanni, L.; Miceli, A.; Francesca, N.; Moschetti, G. Investigation of the hygienic safety of aromatic plants cultivated in soil contaminated with Listeria monocytogenes. Food Control. 2012, 26, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Herrador, B.R.; Nilsen, E.; Cudjoe, K.S.; Jensvoll, L.; Kvamme, J.M.; Aanstad, A.L.; Lindstedt, B.A.; Nygard, K.; Severinsen, G.; Werner-Johansen, O.; Wester, A.L.; Wiklund, M.; Vold, L. A Shigella sonnei outbreak traced to imported basil—the importance of good typing tools and produce traceability systems, Norway, 2011. Eurosurveillance 2013, 18, 15–21. [Google Scholar] [CrossRef]
- Pezzoli, L.; Elson, R.; Little, C.L.; Yip, H.; Fisher, I.; Yishai, R.; Anis, E.; Valinsky, L.; Biggerstaff, M.; Patel, N.; et al. Packed with Salmonella—Investigation of an international outbreak of Salmonella Senftenberg infection linked to contamination of prepacked basil in 2007. Foodborne Pathog. Dis. 2008, 5, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Pakalniskiene, J.; Falkenhorst, G.; Lisby, M.; Madsen, S.B.; Olsen, K.E.P.; Nielsen, E.M.; Mygh, A.; Boel, J.; Molbak, K. A foodborne outbreak of enterotoxigenic E. coli and Salmonella Anatum infection after a high-school dinner in Denmark, November 2006. Epidemiol Infect. 2009, 137, 396–401. [Google Scholar] [CrossRef] [PubMed]
- The Proteobacteria. In Bergey’s Manual of Systematic Bacteriology; Garrity, G.M. (Ed.) Springer-Verlag: New York, NY, USA, 2005; Volume 2.
- Wetzel, K.; Lee, J.; Lee, C.S.; Binkley, M. Comparison of microbial diversity of edible flowers and basil grown with organic versus conventional methods. Can. J. Microbiol. 2010, 56, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, H.; Mossel, D.A.A.; Edberg, S.C.; Struijk, C.B. Advances in the bacteriology of the coliform group: Their suitability as markers of microbial water safety. Ann. Rev. Microbiol. 2001, 55, 201–234. [Google Scholar] [CrossRef] [PubMed]
- Brandl, M.T.; Huynh, S. Effect of the surfactant tween 80 on the detachment and dispersal of Salmonella enterica serovar Thompson single cells and aggregates from cilantro leaves as revealed by image analysis. Appl. Environ. Microbiol. 2014, 80, 6. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.E.; Monier, J.M. The ecological significance of biofilm formation by plant-associated bacteria. Ann. Rev. Phytopathol. 2003, 41, 429–453. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Zhang, T. Biases during DNA extraction of activated sludge samples revealed by high throughput sequencing. Appl. Microbiol. Biotechnol. 2013, 97, 4607–4616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceuppens, S.; Boon, N.; Rajkovic, A.; Heyndrickx, M.; van de Wiele, T.; Uyttendaele, M. Quantification methods for Bacillus cereus vegetative cells and spores in the gastrointestinal environment. J. Microbiol. Meth. 2010, 83, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Maropola, M.K.A.; Ramond, J.B.; Trindade, M. Impact of metagenomic DNA extraction procedures on the identifiable endophytic bacterial diversity in Sorghum bicolor (l. Moench). J. Microbiol. Meth. 2015, 112, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Yu, K.Q.; Deng, G.H.; Jiang, Y.X.; Wang, Y.; Zhang, G.X.; Zhou, H.W. Comparison of direct boiling method with commercial kits for extracting fecal microbiome DNA by Illumina sequencing of 16S rRNA tags. J. Microbiol. Meth. 2013, 95, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Chelius, M.K.; Triplett, E.W. The diversity of Archaea and Bacteria in association with the roots of Zea Mays L. Microbial Ecol. 2001, 41, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Dorn-In, S.; Bassitta, R.; Schwaiger, K.; Bauer, J.; Holzel, C.S. Specific amplification of bacterial DNA by optimized so-called universal bacterial primers in samples rich of plant DNA. J. Microbiol. Meth. 2015, 113, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Ruegger, P.M.; Clark, R.T.; Weger, J.R.; Braun, J.; Borneman, J. Improved resolution of bacteria by high throughput sequence analysis of the rRNA internal transcribed spacer. J. Microbiol. Meth. 2014, 105, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Engelbrektson, A.; Kunin, V.; Wrighton, K.C.; Zvenigorodsky, N.; Chen, F.; Ochman, H.; Hugenholtz, P. Experimental factors affecting PCR-based estimates of microbial species richness and evenness. ISME J. 2010, 4, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Chakravorty, S.; Helb, D.; Burday, M.; Connell, N.; Alland, D. A detailed analysis of 16S ribosomal RNA gene segments for the diagnosis of pathogenic bacteria. J. Microbiol. Meth. 2007, 69, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Ju, F.; Cai, L.; Zhang, T. Taxonomic precision of different hypervariable regions of 16S rRNA gene and annotation methods for functional bacterial groups in biological wastewater treatment. PLoS ONE 2013, 8, e76185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claesson, M.J.; Wang, Q.O.; O'Sullivan, O.; Greene-Diniz, R.; Cole, J.R.; Ross, R.P.; O’Toole, P.W. Comparison of two next-generation sequencing technologies for resolving highly complex microbiota composition using tandem variable 16S rRNA gene regions. Nucl. Acids Res. 2010. [Google Scholar] [CrossRef] [PubMed]
- Mosher, J.J.; Bernberg, E.L.; Shevchenko, O.; Kan, J.; Kaplan, L.A. Efficacy of a 3rd generation high-throughput sequencing platform for analyses of 16S rRNA genes from environmental samples. J. Microbiol. Meth. 2013, 95, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Ercolini, D. High-throughput sequencing and metagenomics: Moving forward in the culture-independent analysis of food microbial ecology. Appl. Environ. Microbiol. 2013, 79, 3148–3155. [Google Scholar] [CrossRef] [PubMed]
- Pylro, V.S.; Roesch, L.F.W.; Morais, D.K.; Clark, I.M.; Hirsch, P.R.; Totola, M.R. Data analysis for 16s microbial profiling from different benchtop sequencing platforms. J. Microbiol. Meth. 2014, 107, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The ribosomal database project: Improved alignments and new tools for rRNA analysis. Nucl. Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved GreenGenes taxonomy with explicit ranks for ecological and evolutionary analyses of Bacteria and Archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The Silva ribosomal RNA gene database project: Improved data processing and web-based tools. Nucl. Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceuppens, S.; Delbeke, S.; De Coninck, D.; Boussemaere, J.; Boon, N.; Uyttendaele, M. Characterization of the Bacterial Community Naturally Present on Commercially Grown Basil Leaves: Evaluation of Sample Preparation Prior to Culture-Independent Techniques. Int. J. Environ. Res. Public Health 2015, 12, 10171-10197. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph120810171
Ceuppens S, Delbeke S, De Coninck D, Boussemaere J, Boon N, Uyttendaele M. Characterization of the Bacterial Community Naturally Present on Commercially Grown Basil Leaves: Evaluation of Sample Preparation Prior to Culture-Independent Techniques. International Journal of Environmental Research and Public Health. 2015; 12(8):10171-10197. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph120810171
Chicago/Turabian StyleCeuppens, Siele, Stefanie Delbeke, Dieter De Coninck, Jolien Boussemaere, Nico Boon, and Mieke Uyttendaele. 2015. "Characterization of the Bacterial Community Naturally Present on Commercially Grown Basil Leaves: Evaluation of Sample Preparation Prior to Culture-Independent Techniques" International Journal of Environmental Research and Public Health 12, no. 8: 10171-10197. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph120810171