Bioaccumulation and Biomagnification of 2-Ethylhexyl-4-dimethylaminobenzoate in Aquatic Animals


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Testing Organisms and Exposure
2.3. Sample Extraction and Chemical Analysis
2.4. Determination of Enzyme Activities
2.5. Statistical Analysis
3. Results and Discussion
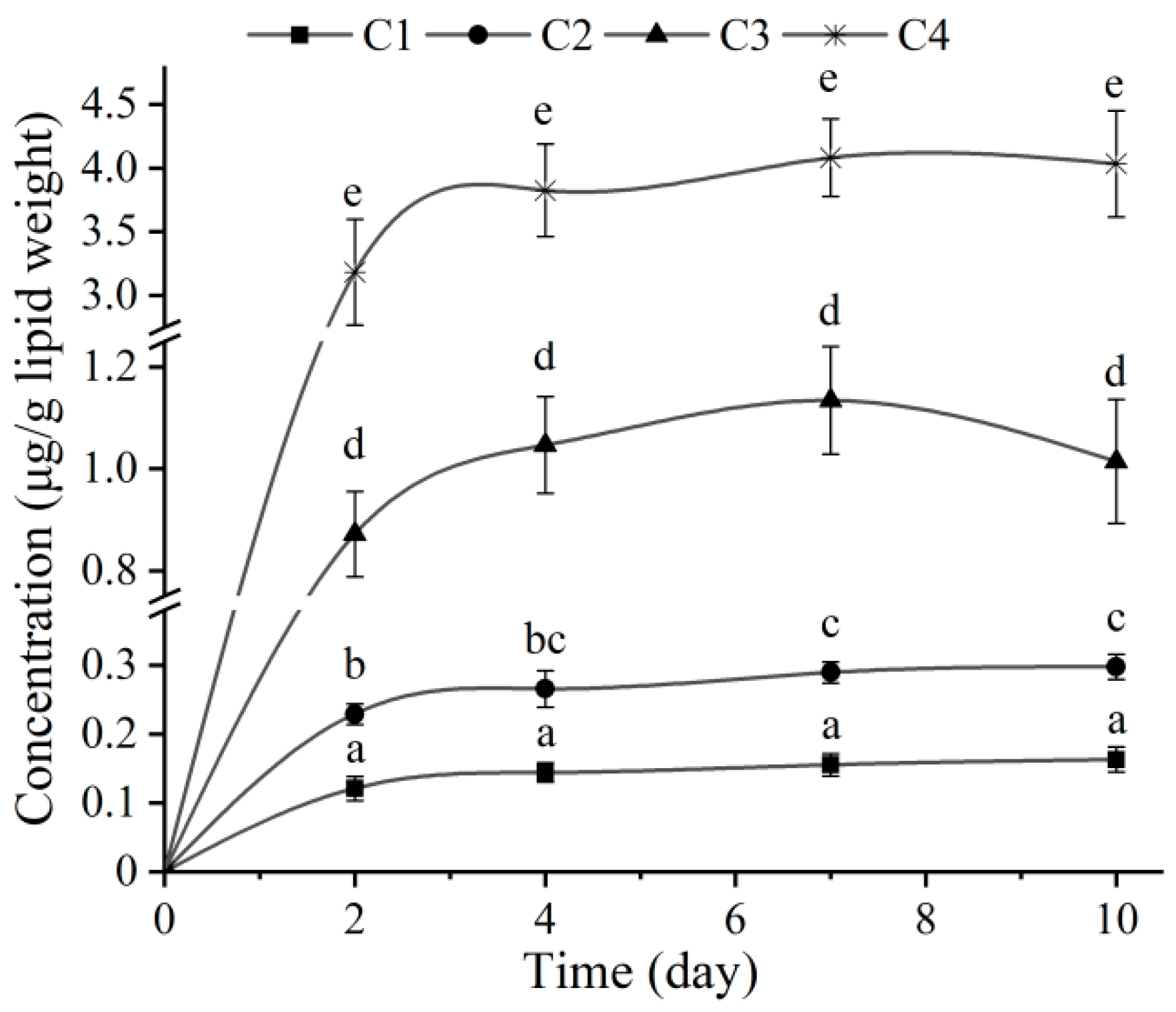
3.1. Bioaccumulation of EHDAB in Midge Larvae
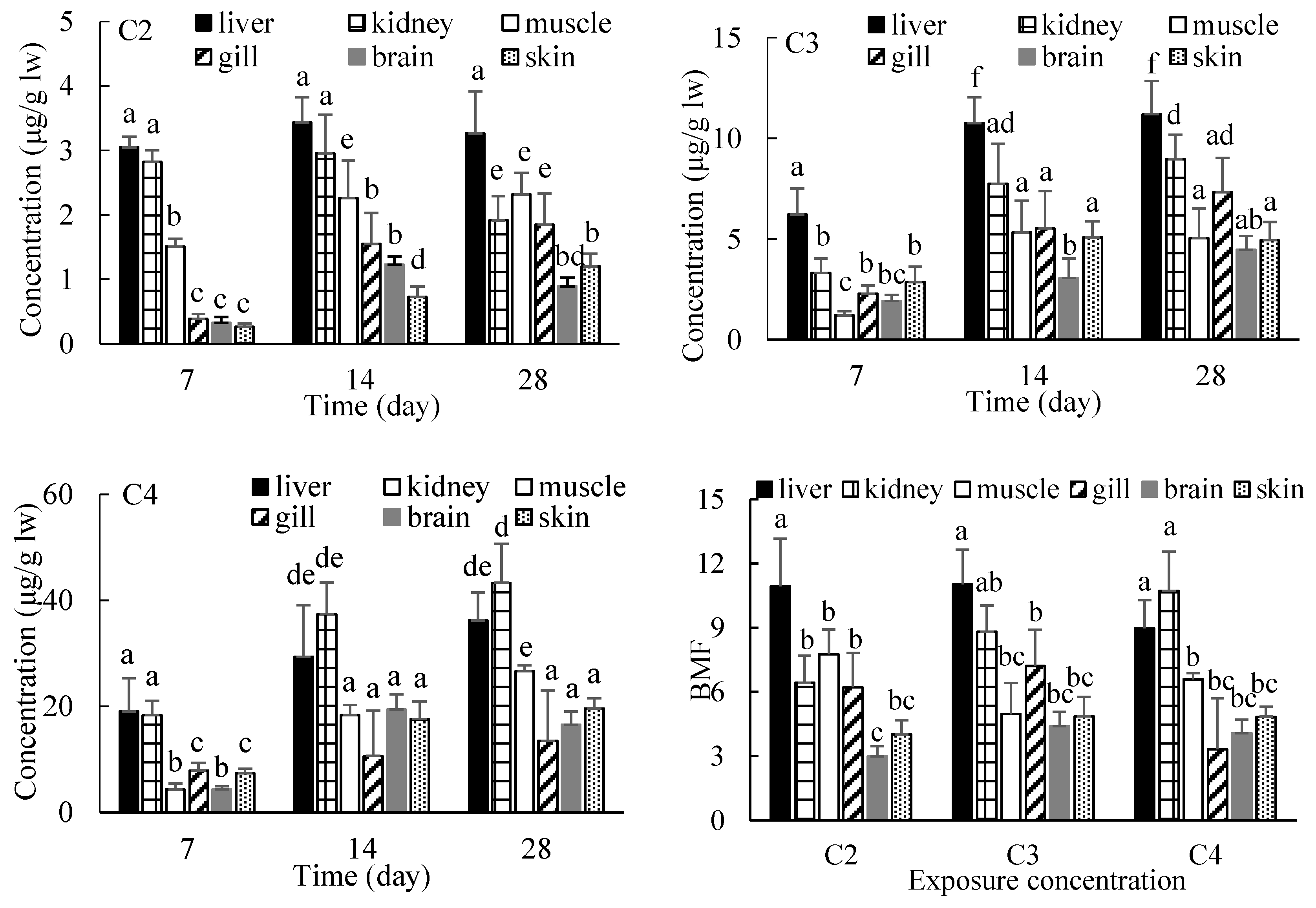
3.2. Biomagnification of EHDAB in Crucian Carp
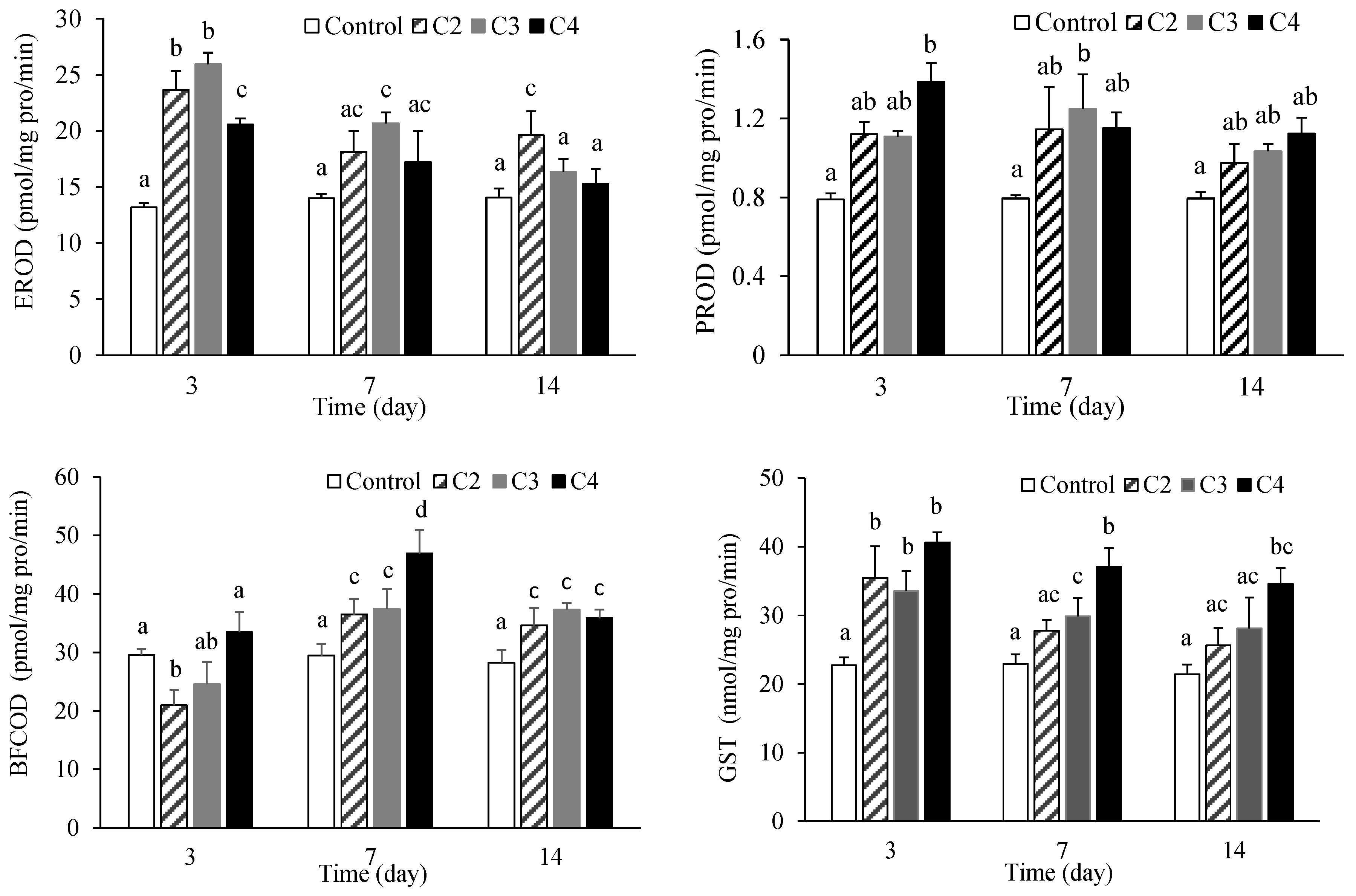
3.3. Effects of EHDAB on Metabolic Enzymes in Fish
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liao, C.; Kannan, K. Widespread occurrence of benzophenone-type UV light filters in personal care products from China and the United States: An assessment of human exposure. Environ. Sci. Technol. 2014, 48, 4103–4109. [Google Scholar] [CrossRef] [PubMed]
- Balmer, M.E.; Buser, H.R.; Müller, M.D.; Muller, M.D.; Poiger, T. Occurrence of some organic UV filters in wastewater, in surface waters, and in fish from Swiss lakes. Environ. Sci. Technol. 2005, 3, 953–962. [Google Scholar] [CrossRef]
- Langford, K.H.; Reid, M.J.; Fjeld, E.; Ãxnevad, S.; Thomas, K.V. Environmental occurrence and risk of organic UV filters and stabilizers in multiple matrices in Norway. Environ. Int. 2015, 80, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rainieri, S.; Barranco, A.; Primec, M.; Langerholc, T. Occurrence and toxicity of musks and UV filters in the marine environment. Food Chem. Toxicol. 2017, 104, 57–66. [Google Scholar]
- Tsui, M.M.; Lam, J.C.; Ng, T.Y.; Ang, P.O.; Murphy, M.B.; Lam, P.K.S. Occurrence, distribution and fate of organic UV filters in coral communities. Environ. Sci. Technol. 2017, 51, 4182–4190. [Google Scholar] [CrossRef] [PubMed]
- Zenker, A.; Schmutz, H.; Fent, K. Simultaneous trace determination of nine organic UV-absorbing compounds (UV filters) in environmental samples. J. Chromatogr. A 2008, 1202, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Molins-Delgado, D.; Máñez, M.; Andreu, A.; Hiraldo, F.; Eljarrat, E.; Barceló, D.; Díaz-Cruz, M.S. A potential new threat to wild life: Presence of UV filters in bird eggs from a preserved area. Environ. Sci. Technol. 2017, 51, 10983–10990. [Google Scholar] [CrossRef] [PubMed]
- Molins-Delgado, D.; Muñoz, R.; Nogueira, S.; Alonso, M.B.; Torres, J.P.; Malm, O.; Ziolli, R.L.; Hauser-Davis, R.A.; Eljarrat, E.; Barceló, D.; et al. Occurrence of organic UV filters and metabolites in lebranche mullet (Mugil liza) from Brazil. Sci. Total Environ. 2018, 618, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Fan, Y.; Jin, J.; Xiong, S.; Liu, J.; Tang, C. Bioaccumulation and biomagnification of ultraviolet absorbents in marine wildlife of the Pearl River Estuarine, South China Sea. Environ. Pollut. 2017, 225, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Liñán, L.; Villaverde-de-Sáa, E.; Rodil, R.; Quintana, J.B.; Beiras, R. Bioaccumulation of UV filters in Mytilus galloprovincialis mussel. Chemosphere 2018, 190, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.; Gravato, C.; Quintaneiro, C.; Golovko, O.; Žlábek, V.; Soares, A.M.V.M.; Pestana, J.L.T. Toxicity of organic UV-filters to the aquatic midge Chironomus riparius. Ecotoxcol. Environ. Saf. 2017, 143, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Wu, Y.T.; Ding, W.H. UV-filter benzophenone-3 inhibits agonistic behavior in male Siamese fighting fish (Betta splendens). Ecotoxicology 2016, 25, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Kunz, P.Y.; Gries, T.; Fent, K. The ultraviolet filter 3-benzylidene camphor adversely affects reproduction in fathead minnow (Pimephales promelas). Toxicol. Sci. 2006, 93, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Li, V.W.T.; Tsui, M.P.M.; Chen, X.; Hui, M.N.Y.; Jin, L.; Lam, R.H.W.; Yu, R.M.K.; Murphy, M.B.; Cheng, J.; Lam, P.K.S.; et al. Effects of 4-methylbenzylidene camphor (4-MBC) on neuronal and muscular development in zebrafish (Danio rerio) embryos. Environ. Sci. Pollut. Res. 2016, 23, 8275–8285. [Google Scholar] [CrossRef] [PubMed]
- Park, C.B.; Jang, J.; Kim, S.; Kim, Y.J. Single- and mixture toxicity of three organic UV-filters, ethylhexyl methoxycinnamate, octocrylene, and avobenzone on Daphnia magna. Ecotoxcol. Environ. Saf. 2017, 137, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ma, X.; Dzakpasu, M.; Wang, X.C. Evaluation of ecotoxicological effects of benzophenone UV filters: Luminescent bacteria toxicity, genotoxicity and hormonal activity. Ecotoxicol. Environ. Saf. 2017, 142, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Lu, G.; Liu, F.; Nie, Y.; Zhang, Z.; Li, Y. Organic UV Filters in the surface water of Nanjing, China: Occurrence, distribution and ecological risk assessment. Bull. Environ. Contam. Toxicol. 2016, 96, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Cruz, M.S.; Gago-Ferrero, P.; Llorca, M.; Barceló, D. Analysis of UV filters in tap water and other clean waters in Spain. Anal. Bioanal. Chem. 2012, 402, 2325–2333. [Google Scholar] [CrossRef] [PubMed]
- Tsui, M.M.; Leung, H.W.; Kwan, B.K.; Ng, K.Y.; Yamashita, N.; Taniyasu, S.; Lam, P.K.; Murphy, M.B. Occurrence, distribution and ecological risk assessment of multiple classes of UV filters in marine sediments in Hong Kong and Japan. J. Hazard. Mater. 2015, 292, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Sang, Z.; Leung, K.S.Y. Environmental occurrence and ecological risk assessment of organic UV filters in marine organisms from Hong Kong coastal waters. Sci. Total Environ. 2016, 566, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lu, G.; Xie, Z.; Ding, J.; Liu, J.; Li, Y. Sorption and degradation of selected organic UV filters (BM-DBM, 4-MBC, and OD-PABA) in laboratory water-sediment systems. Environ. Sci. Pollut. Res. 2016, 23, 9679–9689. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Lu, G.; Liu, J.; Yan, Z.; Yang, H.; Pan, T. Bioconcentration and multi-biomarkers of organic UV filters (BM-DBM and OD-PABA) in crucian carp. Ecotoxicol. Environ. Saf. 2017, 141, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Xia, X.; Wang, F.; Zhang, P.; Zhao, X. Influences of multiwalled carbon nanotubes and plant residue chars on bioaccumulation of polycyclic aromatic hydrocarbons by Chironomus plumosus larvae in sediment. Environ. Toxicol. Chem. 2012, 31, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Frasco, M.F.; Guilhermino, L. Effects of dimethoate and beta-naphthoflavone on selected biomarkers of Poecilia reticulata. Fish Physiol. Biochem. 2002, 26, 149–156. [Google Scholar] [CrossRef]
- Hahn, M.E.; Lamb, T.M.; Schultz, M.E.; Smolowitz, R.M.; Stegeman, J.J. Cytochrome P4501A induction and inhibition by 3,3′,4,4′-tetrachlorobiphenyl in an Ah receptor-containing fish hepatoma cell line (PLHC-1). Aquat. Toxicol. 1993, 26, 185–208. [Google Scholar] [CrossRef]
- Smith, E.M.; Wilson, J.Y. Assessment of cytochrome P450 fluorometric substrates with rainbow trout and killifish exposed to dexamethasone, pregnenolone-16α-carbonitrile, rifampicin, and β-naphthoflavone. Aquat. Toxicol. 2010, 97, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Santos, A.J.; Crista, D.M.A.; Miranda, M.S.; Almeida, I.F.; Sousa e Silva, J.P.; Costa, P.C.; Amaral, M.H.; Lobao, P.A.L.; Sousa Lobo, J.M.; Esteves da Silva, J.C.G. Degradation of UV filters 2-ethylhexyl-4-methoxycinnamate and 4-tert-butyl-4′-methoxydibenzoylmethane in chlorinated water. Environ. Chem. 2013, 10, 127–134. [Google Scholar] [CrossRef]
- Calza, P.; Vione, D.; Galli, F.; Fabbri, D.; Bello, F.D.; Medana, C. Study of the photochemical transformation of 2-ethylhexyl 4-(dimethylamino) benzoate (OD-PABA) under conditions relevant to surface waters. Water Res. 2016, 88, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Contardo-Jara, C.; Lorenz, C.; Pflugmacher, S.; Nutzmann, G.; Kloas, W.; Wiegand, C. Exposure to human pharmaceuticals carbamazepine, ibuprofen and bezafibrate causes molecular effects in Dreissena polymorpha. Aquat. Toxicol. 2011, 105, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lu, G.; Yang, H.; Yan, Z.; Wang, Y.; Wang, P. Bioconcentration and metabolism of ketoconazole and effects on multi-biomarkers in crucian carp (Carassius auratus). Chemosphere 2016, 150, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Mehinto, A.C.; Hill, E.M.; Tyler, C.R. Uptake and biological effects of environmentally relevant concentrations of the nonsteroidal anti-inflammatory pharmaceutical diclofenac in rainbow trout (Oncorhynchus mykiss). Environ. Sci. Technol. 2010, 44, 2176–2182. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lu, G.; Ding, J.; Zhang, Z.; Wang, Y. Tissue distribution, bioconcentration, metabolism, and effects of erythromycin in crucian carp (Carassius auratus). Sci. Total Environ. 2014, 490, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.L.; Oris, J.T. CYP1A expression in liver and gill of rainbow trout following waterborne exposure: Implications for biomarker determination. Aquat. Toxicol. 1999, 46, 279–287. [Google Scholar] [CrossRef]
- Fent, K.; Zenker, A.; Rapp, M. Widespread occurrence of estrogenic UV-filters in aquatic ecosystems in Switzerland. Environ. Pollut. 2010, 158, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Ribalta, C.; Solé, M. In vitro interaction of emerging contaminants with the cytochrome P450 system of Mediterranean deep-sea fish. Environ. Sci. Technol. 2014, 48, 12327–12335. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lu, G.; Wang, Y.; Yan, Z.; Yang, X.; Ding, J.; Jiang, Z. Bioconcentration, metabolism, and biomarker responses in freshwater fish Carassius auratus exposed to roxithromycin. Chemosphere 2014, 99, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Lu, G.; Li, S.; Nie, Y.; Liu, J. Biological fate and effects of propranolol in an experimental aquatic food chain. Sci. Total Environ. 2015, 532, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Lu, G.; Hou, K.; Qin, D.; Yan, Z.; Chen, W. Bioconcentration, metabolism and effects of diphenhydramine on behavioral and biochemical markers in crucian carp (Carassius auratus). Sci. Total Environ. 2016, 544, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, B.M.; Berg, K.; Goksoyr, A. Induction of cytochrome P4501A (CYP1A) in fish a biomarker for environmental pollution. Methods Mol. Biol. 1998, 107, 23–438. [Google Scholar]
- Ding, J.; Lu, G.; Li, Y. Interactive effects of selected pharmaceutical mixtures on bioaccumulation and biochemical status in crucian carp (Carassius auratus). Chemosphere 2016, 148, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Grabicova, K.; Fedorova, G.; Burkina, V.; Steinbach, C.; Schmidt-Posthaus, H.; Zlabek, V.; Kroupova, H.K.; Graic, R.; Randak, T. Presence of UV filters in surface water and the effects of phenylbenzimidazole sulfonic acid on rainbow trout (Oncorhynchus mykiss) following a chronic toxicity test. Ecotoxicol. Environ. Saf. 2013, 96, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.B.; Fisk, A.T.; Brown, M.; Villella, M.; Muir, D.C.G.; Evans, R.E.; Lockhart, W.L.; Metner, D.A.; Cooley, H.M. Dietary accumulation and biochemical responses of juvenile rainbow trout (Oncorhynchus mykiss) to 3,3′,4,4′,5-pentachlorobiphenyl (PCB 126). Aquat. Toxicol. 2002, 59, 139–152. [Google Scholar] [CrossRef]
- Danis, B.; Goriely, S.; Dubois, P.; Fowler, S.W.; Flamand, V.; Warnau, M. Contrasting effects of coplanar versus non-coplanar PCB congeners on immunomodulation and CYP1A levels (determined using an adapted ELISA method) in the common sea star Asterias rubens L. Aquat. Toxicol. 2004, 69, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Leon-Gonzalez, Z.; Ferreiro-Vera, C.; Priego-Capote, F.; Luque de Castro, M.D. Targeting metabolomics analysis of the sunscreen agent 2-ethylhexyl 4-(N,N-dimethylamino) benzoate in human urine by automated on-line solid-phase extraction-liquid chromatography-tandem mass spectrometry with liquid chromatography-time-of-flight/mass spectrometry confirmation. J. Chromatogr. A 2011, 1218, 3013–3021. [Google Scholar] [PubMed]
- Liang, Y.; Zhan, J.; Liu, X.; Zhou, Z.; Zhu, W.; Liu, D.; Wang, P. Stereoselective metabolism of the UV-filter 2-ethylhexyl 4-dimethylaminobenzoate and its metabolites in rabbits in vivo and vitro. RSC Adv. 2017, 7, 16991–16996. [Google Scholar] [CrossRef] [Green Version]
- Richardson, B.J.; Mak, E.; De Luca-Abbott, S.B.; Martin, M.; McClellan, K.; Lam, P.K.S. Antioxidant responses to polycyclic aromatic hydrocarbons and organochlorine pesticides in green-lipped mussels (Perna viridis): Do mussels “integrate” biomarker responses? Mar. Pollut. Bull. 2008, 57, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, S.; Praskova, E.; Chromcova, L.; Plhalova, L.; Prokes, M.; Blahova, J.; Svobodova, Z. The effects of diclofenac on early life stages of common carp (Cyprinus carpio). Environ. Toxicol. Pharmacol. 2013, 35, 454–460. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Concentration (ng/g dw) | 0 d | 2 d | 4 d | 7 d | 10 d |
|---|---|---|---|---|---|
| C1 | 3.9 ± 0.7 | 3.9 ± 0.8 | 3.8 ± 0.6 | 3.6 ± 0.7 | 3.3 ± 0.5 |
| C2 | 19.2 ± 3.6 | 18.8 ± 2.8 | 18.7 ± 2.3 | 18.1 ± 1.9 | 17.2 ± 2.1 |
| C3 | 95.6 ± 12.8 | 94.7 ± 11.5 | 92.5 ± 10.6 | 89.5 ± 11.9 | 84.6 ± 9.4 |
| C4 | 486.6 ± 24.2 | 482.1 ± 37.8 | 479.2 ± 28.2 | 467.7 ± 36.4 | 445.5 ± 32.9 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, G.; Zhou, R.; Li, S.; Dang, T.; Liu, J. Bioaccumulation and Biomagnification of 2-Ethylhexyl-4-dimethylaminobenzoate in Aquatic Animals. Int. J. Environ. Res. Public Health 2018, 15, 2395. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15112395
Lu G, Zhou R, Li S, Dang T, Liu J. Bioaccumulation and Biomagnification of 2-Ethylhexyl-4-dimethylaminobenzoate in Aquatic Animals. International Journal of Environmental Research and Public Health. 2018; 15(11):2395. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15112395
Chicago/Turabian StyleLu, Guanghua, Ranran Zhou, Sheng Li, Tianjian Dang, and Jianchao Liu. 2018. "Bioaccumulation and Biomagnification of 2-Ethylhexyl-4-dimethylaminobenzoate in Aquatic Animals" International Journal of Environmental Research and Public Health 15, no. 11: 2395. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15112395