Emerging Mosquito-Borne Threats and the Response from European and Eastern Mediterranean Countries

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. National Mosquito-Borne Threats
2.1. United Kingdom
2.2. Sweden
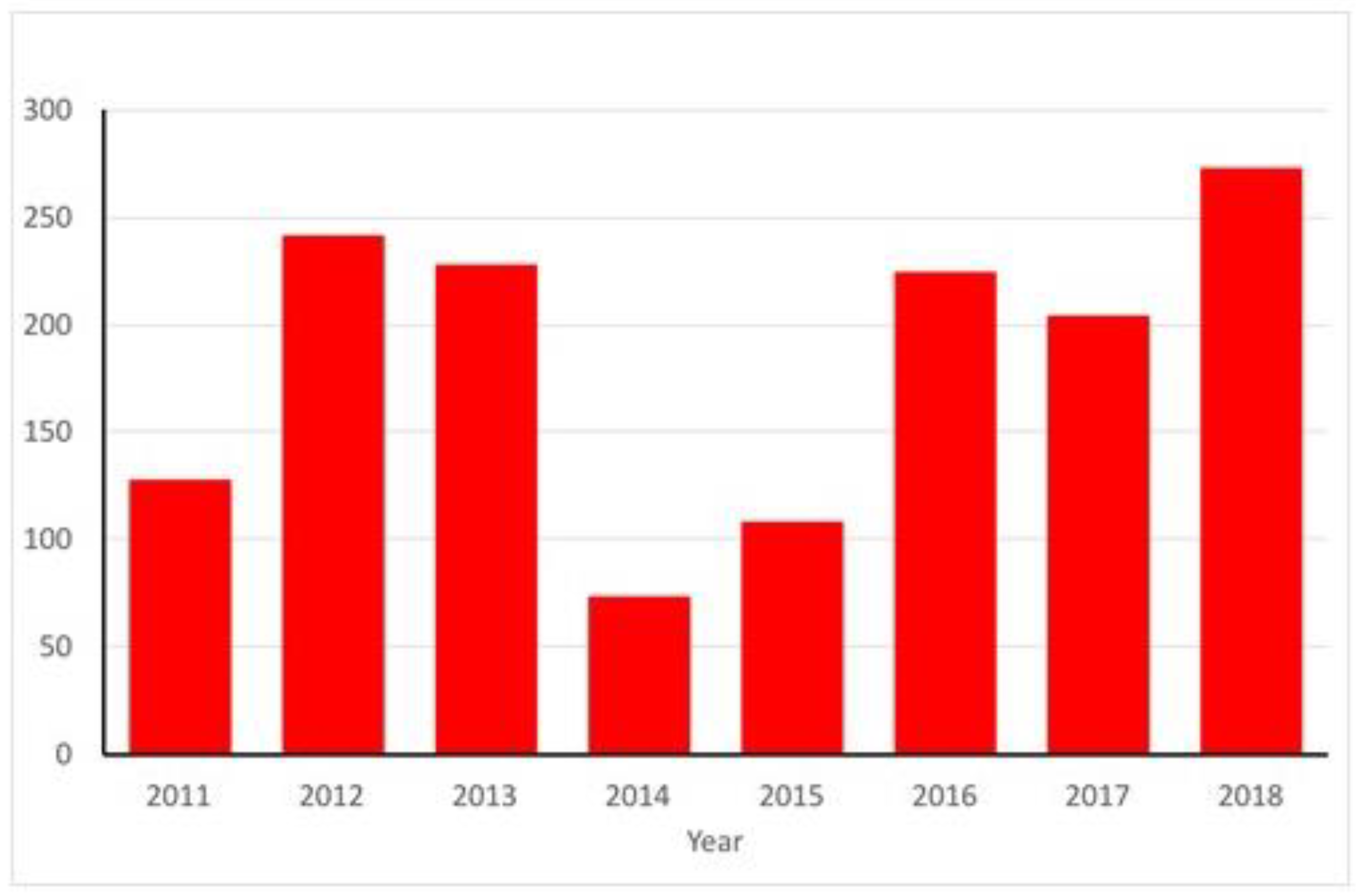
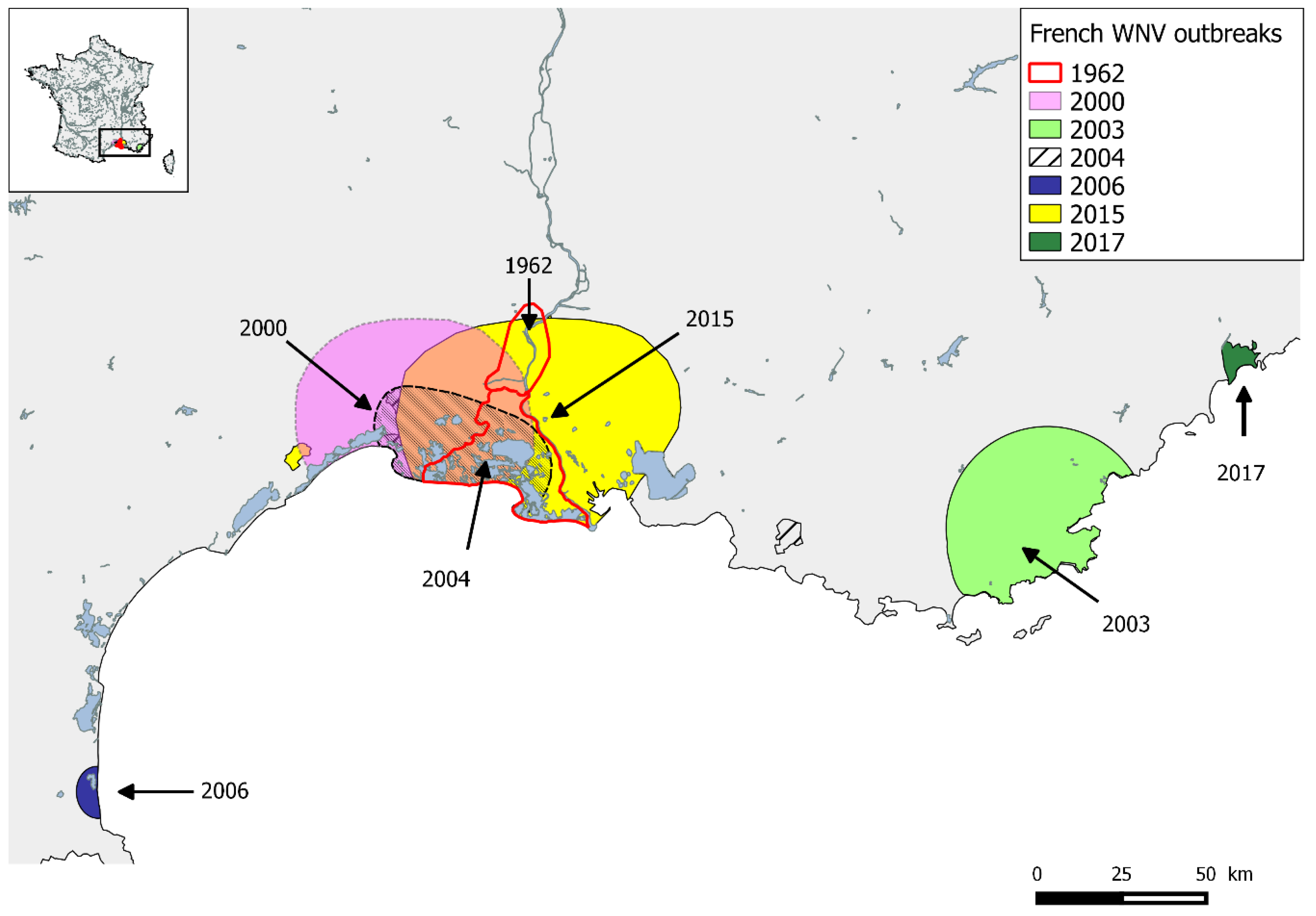
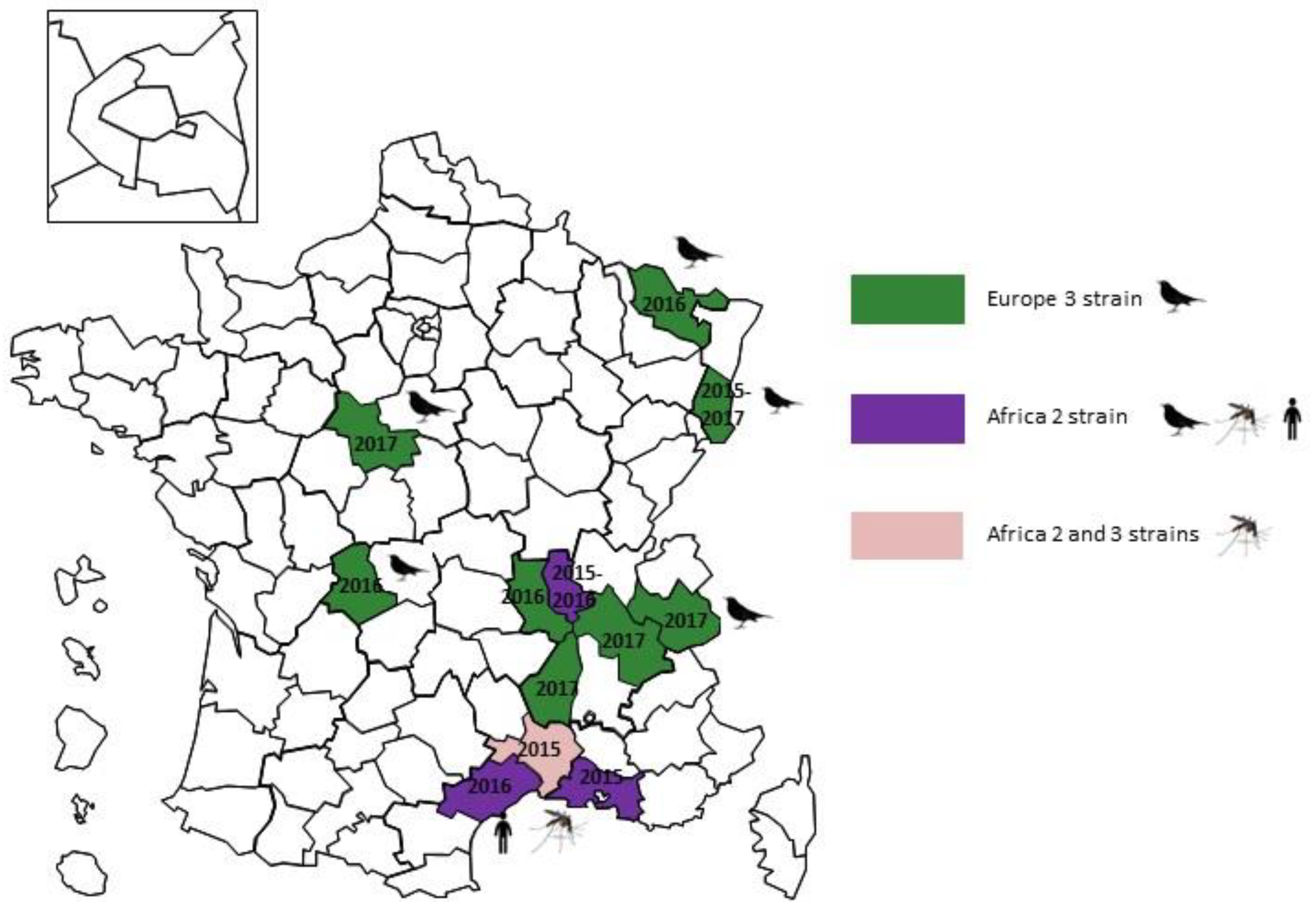
2.3. France
2.4. Italy
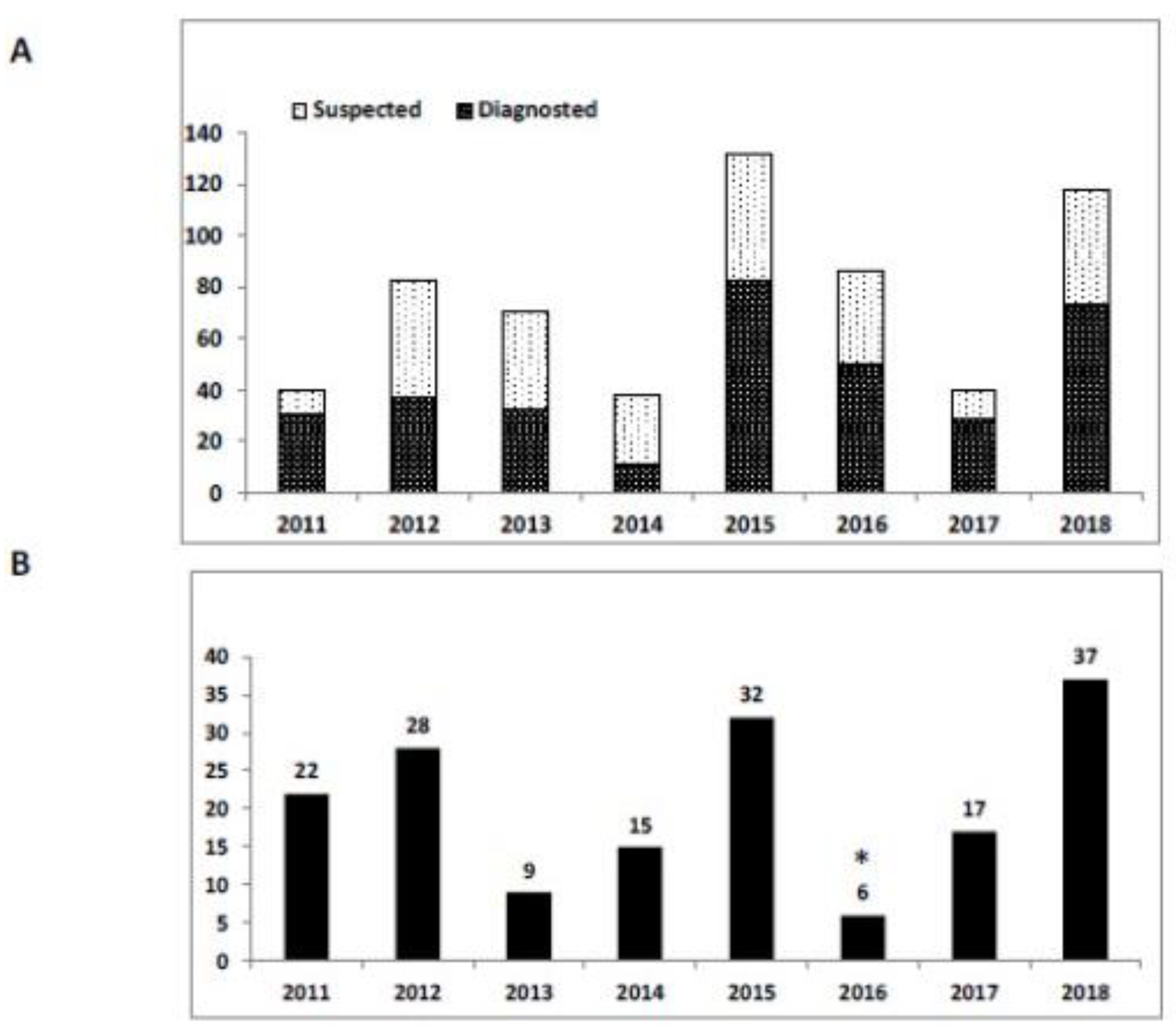
2.5. Israel
2.6. Germany
3. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Landscape effects on the presence, abundance and diversity of mosquitoes in Mediterranean wetlands. PLoS ONE 2015, 10, e0128112. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.; Vaux, A. Seasonal dynamics and habitat specificity of mosquitoes in an English wetland: Implications for UK wetland management and restoration. J. Vector Ecol. 2015, 40, 90–106. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.; Schaffner, F.; Verstiert, V.; Hendrickx, G.; Zeller, G.; Van Bortel, W. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffner, F.; Medlock, J.; Van Bortel, W. Public health significance of invasive mosquitoes in Europe. Clin. Microbiol. Infect. 2013, 19, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Eritja, R.; Palmer, J.; Roiz, D.; Sanpera-Calbet, I.; Bartumeus, F. Direct evidence of adult Aedes albopictus dispersal by car. Sci. Rep. 2017, 24, 14399. [Google Scholar] [CrossRef] [PubMed]
- Aranda, C.; Eritja, R.; Roiz, D. First record and establishment of the mosquito Aedes albopictus in Spain. Med. Vet. Entomol. 2006, 20, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Collantes, F.; Delacour, S.; Alarcón, P.; Ruiz-Arrondo, I.; Delgado, J.; Torrell-Sorio, A.; Bengoa, M.; Eritja, R.; Miranda, M.; Molina, R.; Lucientes, J. Review of ten-years presence of Aedes albopictus in Spain 2004-2014: Known distribution and public health concerns. Parasit. Vectors 2015, 8, 655. [Google Scholar] [CrossRef]
- Eritja, R.; Escosa, R.; Lucientes, J.; Marquès, E.; Molina, R.; Roiz, D.; Ruiz, S. Worldwide invasion of vector mosquitoes: Present European distribution and challenges for Spain. Biol. Invasions 2005, 7, 87–97. [Google Scholar] [CrossRef]
- Engler, O.; Savini, G.; Papa, A.; Figuerola, J.; Groschup, M.; Kampen, H.; Medlock, J.; Vaux, A.; Wilson, A.; Werner, D.; et al. European surveillance for West Nile virus in mosquito populations. Int. J. Environ. Res. Public Health 2013, 10, 4869–4895. [Google Scholar] [CrossRef] [Green Version]
- Delbue, S.; Ferrante, P.; Mariotto, S.; Zanusso, G.; Pavone, A.; Chinaglia, M.; L’Erario, R.; Monaco, S.; Ferrari, S. Review of West Nile virus epidemiology in Italy and report of a case of West Nile virus encephalitis. J. Neurovirol. 2014, 20, 437–441. [Google Scholar] [CrossRef]
- Hernández-Triana, L.; Jeffries, C.; Mansfield, K.; Carnell, G.; Fooks, A.; Johnson, N. Emergence of West Nile virus lineage 2 in Europe: A review on the introduction and spread of a mosquito-borne disease. Front. Public Health 2014, 2, 271. [Google Scholar] [CrossRef] [PubMed]
- ECDC, European Centre for Disease Prevention and Control. Weekly Updates: 2018 West Nile Fever Transmission Season. Available online: https://ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/disease-data-ecdc (accessed on 2 October 2018).
- ECDC, European Centre for Disease Prevention and Control. Historical Data by Year—West Nile Fever Seasonal Surveillance. Available online: https://ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/historical (accessed on 2 October 2018).
- Vogels, C.B.; Möhlemann, T.W.; Melsen, D.; Favia, G.; Wennergren, U.; Koenraadt, C.J. Latitudinal diversity of Culex pipiens biotypes and hybrids in farm, peri-urban, and wetland habitats in Europe. PLoS ONE 2016, 11, e0166959. [Google Scholar] [CrossRef] [PubMed]
- Balenghien, T.; Vazeille, M.; Grandadam, M.; Schaffner, F.; Zeller, H.; Reiter, P.; Sabatier, P.; Fouque, F.; Bicout, D. Vector competence of some French Culex and Aedes mosquitoes for West Nile virus. Vector Borne Zoonotic Dis. 2008, 8, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Succo, T.; Leparc-Goffart, I.; Ferré, J.; Roiz, D.; Broche, B.; Maquart, M.; Noel, H.; Catelinois, O.; Entezam, F.; Caire, D.; Jourdain, F.; et al. Autochthonous dengue outbreak in Nimes, South of France, July to September 2015. Euro Surveill. 2016, 21, 30240. [Google Scholar] [CrossRef] [PubMed]
- Bordi, L.; Carletti, F.; Lalle, E.; Colavita, F.; Meschi, S.; Di Caro, A.; Nicastri, W.; Scognamidglio, P.; Vairo, F.; Di Lallo, D.; et al. Molecular characterization of autochthonous Chikungunya cluster in Latium Region, Italy. Emerg. Infect. Dis. 2018, 24, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Engel, D.; Jöst, H.; Wink, M.; Börstler, J.; Bosch, S.; Gangliany, M.; Jöst, A.; Czajka, C.; Lühken, R.; Ziegler, U.; et al. Reconstruction of the evolutionary history and dispersal of Usutu virus, a neglected emerging arbovirus in Europe and Africa. MBio 2016, 7, e01938. [Google Scholar] [CrossRef] [PubMed]
- Lühken, R.; Jöst, H.; Cadar, D.; Thomas, S.M.; Bosch, S.; Tannich, E.; Becker, N.; Ziegler, U.; lachmann, L.; Schmidt-Chanasit, J. Distribution of Usutu virus in Germany and its effects on breeding bird populations. Emerg. Infect. Dis. 2017, 23, 1991–1998. [Google Scholar] [CrossRef]
- Gossner, C.; Marrama, L.; Carson, M.; Allerberger, F.; Calistri, P.; Dilaveris, D.; Lecollinet, S.; Morgan, D.; Morgan, D.; Nowotny, N.; et al. West Nile virus surveillance in Europe: Moving towards an integrated animal-human-vector approach. Euro Surveill. 2016, 22, 30526. [Google Scholar] [CrossRef]
- Venter, M. Assessing the zoonotic potential of arboviruses of African origin. Curr. Opin. Virol. 2018, 28, 74–84. [Google Scholar] [CrossRef]
- Jeffries, C.L.; Mansfield, K.L.; Phipps, L.P.; Wakeley, P.R.; Mearns, R.; Schock, A.; Bell, S.; Breed, A.C.; Fooks, A.R.; Johnson, N. Louping ill virus: An endemic tick-borne disease of Great Britain. J. Gen. Virol. 2014, 90, 1005–1014. [Google Scholar] [CrossRef]
- Mackenzie-Impoinvil, L.; Impoinvil, D.E.; Galbraith, S.E.; Dillon, R.J.; Ranson, H.; Johnson, N.; Fooks, A.R.; Solomon, T.; Baylis, M. Evaluation of a temperate climate mosquito, Ochlerotatus detritus (=Aedes detritus), as a potential vector of Japanese encephalitis virus. Med. Vet. Entomol. 2015, 29, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Blagrove, M.S.; Sherlock, K.; Chapman, G.E.; Impoinvil, D.E.; McCall, P.J.; Medlock, J.M.; Lycett, G.; Solomon, T.; Baylis, M. Evaluation of the vector competence of a native UK mosquito Ochlerotatus detritus (Aedes detritus) for dengue, chikungunya and West Nile viruses. Parasit. Vectors 2016, 9, 452. [Google Scholar] [CrossRef] [PubMed]
- Lumley, S.; Hernández-Triana, L.M.; Horton, D.L.; Fernández de Marco, M.D.M.; Medlock, J.M.; Hewson, R.; Fooks, A.R.; Johnson, N. Competence of mosquitoes native to the United Kingdom to support replication and transmission of Rift Valley fever virus. Parasit. Vectors 2018, 11, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Triana, L.M.; Fernández de Marco, M.; Mansfield, K.L.; Thorne, L.; Lumley, S.; Marston, D.A.; Fooks, A.R.; Johnson, N. Assessment of vector competence of UK mosquitoes for Usutu virus of African origin. Parasit. Vectors 2018, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.; Dawson, A.; Moss, S.R.; Hinsley, S.A.; Bellamy, P.E.; Gould, E.A. Serological evidence of West Nile virus, Usutu virus and Sindbis virus infection of birds in the UK. J. Gen. Virol. 2003, 84, 2807–2817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckley, A.; Dawson, A.; Gould, E.A. Detection of seroconversion to West Nile virus, Usutu virus and Sindbis virus in UK sentinel chickens. Virol. J. 2006, 3, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, D.L.; Lawson, B.; Egbetade, A.; Jeffries, C.; Johnson, N.; Cunningham, A.A.; Fooks, A.R. Targeted surveillance for Usutu virus in British birds (2005–2011). Vet. Rec. 2013, 172, 17. [Google Scholar] [CrossRef]
- Brugman, V.A.; Horton, D.L.; Phipps, L.P.; Johnson, N.; Cook, A.J.C.; Fooks, A.R.; Breed, A.C. Epidemiological perspectives on West Nile virus surveillance in wild birds in Great Britain. Epidemiol. Infect. 2013, 141, 1134–1142. [Google Scholar] [CrossRef]
- Vaux, G.C.V.; Gibson, G.; Hernandez-Triana, L.M.; Cheke, R.A.; McCracken, F.; Jeffries, C.L.; Horton, D.L.; Springate, S.; Johnson, N.; Fooks, A.R.; Leach, S.; Medlock, J.M. Enhanced West Nile surveillance in the North Kent marshes, UK. Parasit. Vectors 2015, 8, 91. [Google Scholar] [CrossRef]
- Rijks, J.M.; Kik, M.L.; Slaterus, R.; Foppen, R.; Stroo, A.; Ijzer, J.; Stahl, J.; Gröne, A.; Koopmans, M.; van der Jeugd, H.P.; Reusken, C. Widespread Usutu virus outbreak in birds in the Netherlands, 2016. Euro Surveill. 2016, 21, 30391. [Google Scholar] [CrossRef]
- Golding, N.; Nunn, M.A.; Medlock, J.M.; Purse, B.V.; Vaux, A.G.; Schäfer, S.M. West Nile virus vector Culex modestus established in southern England. Parasit. Vectors. 2012, 5, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cull, B.; Vaux, A.G.C.; Medlock, J.M.; Abbot, A.; Gibson, G. Expansion of the range of the West Nile virus vector in Essex. Vet. Rec. 2016, 179, 363–364. [Google Scholar] [CrossRef] [PubMed]
- Mayor, S. More than 50 UK Zika cases are reported in people returning from abroad so far this year. BMJ 2016, 354, i4227. [Google Scholar] [CrossRef] [PubMed]
- Connell, J.; McKeown, P.; Garvey, P.; Cotter, S.; Conway, A.; O’Flanagan, D.; O’Herleihy, B.P.; Morgan, D.; Nicoll, A.; Lloyd, G. Two linked cases of West Nile virus (WNV) acquired by Irish tourists in the Algarve, Portugal. Euro Surveill. 2004, 8, 2517. [Google Scholar]
- Gabriel, M.; Emmerich, P.; Frank, C.; Fiedler, M.; Rashidi-Alavijeh, J.; Jochum, C.; Günther, S.; Auerhammer, K.; Rupprecht, H.-J.; Blank, R.-T.; et al. Increase in West Nile virus infections imported to Germany in 2012. J. Clin. Virol. 2013, 58, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Fooks, A.R.; Horton, D.L.; Phipps, L.P.; Mansfield, K.L.; McCracken, F.; Jefffries, C.L.; Johnson, N.; Wrigley, S.; Deuchande, R.; Toth, B.; et al. Suspect West Nile encephalitis in a horse imported into the UK from Europe. Vet. Rec. 2014, 2, e000104. [Google Scholar] [CrossRef]
- Murphy, G.; Vaux, A.; Medlock, J. Challenges in undertaking mosquito surveillance at UK seaports and airports to prevent the entry and establishment of invasive vector species. Int. J. Environ. Health Res. 2013, 23, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Vaux, A.G.; Cull, B.; Schaffner, F.; Gillingham, E.; Pfluger, V.; Leach, S. Detection of the invasive mosquito species Aedes albopictus in southern England. Lancet. Infect. Dis. 2017, 17, 140. [Google Scholar] [CrossRef]
- Bessell, P.R.; Robinson, R.A.; Golding, N.; Searle, K.R.; Handel, I.G.; Boden, L.A.; Purse, B.V.; Bronsvoort, B.M.D.C. Quantifying the risk of introduction of West Nile virus into Great Britain by migrating passerine birds. Transbound. Emerg. Dis. 2016, 63, e347–e359. [Google Scholar] [CrossRef]
- Lundström, J.O. Mosquito-borne viruses in western Europe: A review. J. Vector. Ecol. 1999, 24, 1–39. [Google Scholar]
- Bergquist, J.; Forsman, O.; Larsson, P.; Näslund, J.; Lilja, T.; Engdahl, C.; Linström, A.; Gylfe, Á.; Ahlm, C.; Evander, M.; Bucht, G. Detection and isolation of Sindbis virus from mosquitoes captured during an outbreak in Sweden, 2013. Vector Borne Zoonotic Dis. 2015, 15, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Putkuri, N.; Kurkela, S.; Levanov, L.; Huhtamo, E.; Vaheri, A.; Sironen, T.; Vapalahti, O. Isolation and characterization of a California encephalitis serogroup orthobunyavirus from Finnish mosquitoes. Infect. Genet. Evol. 2014, 22, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Tingström, O.; Lwande, O.W.; Näslund, J.L.; Spyckerelle, I.; Engdahl, C.; Von Schoenberg, P.; Alhm, C.; Evander, M.; Bucht, G. Detection of Sindbis and Inkoo virus RNA in Genetically typed mosquito larvae sampled in Northern Sweden. Vector Borne Zoonotic Dis. 2016, 16, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Murgue, B.; Murri, S.; Triki, H.; Deubel, V.; Zeller, H.G. West Nile in the Mediterranean basin: 1950–2000. Ann. N. Y. Acad. Sci. 2001, 951, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Bahoun, C.; Marcillaud-Pitel, C.; Bournez, L.; Leblond, A.; Beck, C.; Hars, J.; Leparc-Goffart, I.; L’Ambert, G.; Paty, M.C.; Daix, C.; et al. West Nile virus epizootics in the Camargue (France) in 2015 and reinforcement of surveillance and control networks. Rev. Sci. Tech. 2016, 35, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Lecollinet, S.; Blanchard, Y.; Manson, C.; Lowenski, S.; Laloy, E.; Quenault, H.; Touzain, F.; Lucas, P.; Eraud, C.; Bahoun, C.; et al. Dual emergence of Usutu virus in common blackbirds, Eastern France, 2015. Emerg. Infect. Dis. 2016, 22, 2225. [Google Scholar] [CrossRef] [PubMed]
- Eiden, M.; Gil, P.; Ziegler, U.; Rakotoarizony, I.; Marie, A.; Frances, B.; L’Ambert, G.; Simonin, Y.; Foulongne, V.; Groschup, M.H.; Gurierrez, S.; Eloit, M. Emergence of two Usutu virus lineages in Culex pipiens mosquitoes in the Camargue, France, 2015. Infect. Genet. Evol. 2018, 61, 151–154. [Google Scholar] [CrossRef]
- Simonin, Y.; Sillam, O.; Carles, M.J.; Gutierrez, Z.; Gil, P.; Constant, O.; Martin, M.F.; Girard, G.; Van de Perre, P.; Salinas, S.; Leparc-Goffart, I.; Foulongne, V. Human Usutu virus infection with atypical neurological presentation, Montpellier, France, 2016. Emerg. Infect. Dis. 2018, 24, 875–878. [Google Scholar] [CrossRef]
- Delaunay, P.; Mathieu, B.; Marty, P.; Fauran, P.; Schaffner, F. Chronology of the development of Aedes albopictus in the Alpes-Maritimes Department of France, from 2002–2005. Med. Trop. 2007, 67, 310–311. [Google Scholar]
- Sardelis, M.R.; Turell, B.; O’Guinn, M.L.; Andre, R.G.; Roberts, D.R. Vector competence of three North American strains of Aedes albopictus for West Nile virus. J. Am. Mosq. Control Assoc. 2002, 18, 284–289. [Google Scholar]
- Marchande, E.; Prat, C.; Jeannin, C.; Lafont, E.; Bergmann, T.; Flusin, O.; Rizzi, J.; Roux, N.; Busso, V.; Deniau, J.; Noel, H.; et al. Autochthonous case of dengue in France, October 2013. Euro Surveill. 2013, 18, 20661. [Google Scholar] [CrossRef] [Green Version]
- Paty, M.C.; Six, C.; Charlet, F.; Heuzé, G.; Cochet, A.; Wiegandt, A.; Chappert, J.L.; Dejour-Salamanca, D.; Guinard, A.; Soler, P.; et al. Large number of imported chikungunya cases in mainland France, 2014: A challenge for surveillance and response. Euro Surveill. 2014, 19, 20856. [Google Scholar] [CrossRef] [PubMed]
- Circulaire Interministérielle DGS/R11/DFALN/DGAL no 2009-233 du 24 Juillet 2009 Relative Aux Measures Visant à Limiter la Circulation du Virus West Nile en France Métropolitaine. Available online: https://solidarites-sante.gouv.fr/fichiers/bo/2009/09-08/ste_20090008_0100_0125.pdf (accessed on 3 October 2018).
- Delise, E.; Rousseau, C.; Broche, B.; Leparc-Goffart, I.; L’Ambert, G.; Cochet, A.; Prat, C.; Foulongne, V.; Ferre, J.B.; Catelinois, O.; et al. Chikungunya outbreak in Montpellier, France, September to October 2014. Euro Surveill. 2015, 20, 21108. [Google Scholar] [CrossRef] [Green Version]
- Mancini, G.; Montarsi, F.; Calzolari, M.; Capelli, G.; Dottori, M.; Ravagnan, S.; Lelli, D.; Santilli, A.; Quagliam, M.; Federici, V.; Monaco, F.; et al. Mosquito species involved in the circulation of West Nile and Usutu virus in Italy. Vet. Ital. 2017, 53, 97–110. [Google Scholar] [PubMed]
- Brunetti, M. Climate Monitoring for Italy. Institute of Atmospheric Sciences and Climate. Available online: http://www.isac.cnr.it/climstor/climate_news.html#top (accessed on 26 September 2018).
- Kirkland, P.D. Akabane and bovine ephemeral fever virus infections. Vet. Clin. North Am. Food Anim. Pract. 2002, 18, 501–514. [Google Scholar] [CrossRef]
- Schwartz, E. Study of dengue fever among Israeli travelers to Thailand. Dengue Bull. 2002, 26, 162. [Google Scholar]
- Meltzer, E.; Lustig, Y.; Leshem, E.; Levy, R.; Gottesman, G.; Weissmann, R.; Rabi, D.H.; Hindiyeh, M.; Koren, R.; Mendelson, E.; et al. Zika virus disease in traveler returning from Vietnam to Israel. Emerg. Infect. Dis. 2016, 22, 1521–1522. [Google Scholar] [CrossRef]
- Avni, C.; Stienlauf, S.; Meltzer, E.; Sidi, Y.; Schwartz, E.; Leshem, E. Region-Specific, life-threatening diseases among international travelers from Israel, 2004–2015. Emerg. Infect. Dis. 2018, 24, 790–793. [Google Scholar] [CrossRef]
- Mannasse, B.; Mendelson, E.; Orshan, L.; Mor, O.; Shalom, U.; Yeger, T.; Lustig, Y. Usutu virus RNA in mosquitoes, Israel, 2014–2015. Emerg. Infect. Dis. 2017, 23, 1699–1702. [Google Scholar] [CrossRef]
- Bernkopf, H.; Levine, S.; Nerson, R. Isolation of West Nile virus in Israel. J. Infect. Dis. 1953, 93, 207–218. [Google Scholar] [CrossRef]
- Lustig, Y.; Hindiyeh, M.; Orshan, L.; Weiss, L.; Koren, R.; Katz-Likvornik, S.; Zadka, H.; Glatman-Freedman, A.; Mendelson, E.; Shulman, L.M. Mosquito surveillance for 15 years reveals high genetic diversity among West Nile viruses in Israel. J. Infect. Dis. 2016, 213, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Steinman, A.; Banet, C.; Sutton, G.A.; Yadin, H.; Hadar, S.; Brill, A. Clinical signs of West Nile virus encephalomyelitis in horses during the outbreak in Israel in 2000. Vet. Rec. 2002, 151, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Banet-Noach, C.; Malkinson, M.; Brill, A.; Samina, I.; Yadin, H.; Weisman, Y.; Pokamunski, S.; King, R.; Deubel, V.; Stram, Y. Phylogenetic relationships of West Nile viruses isolated from birds and horses in Israel from 1997 to 2001. Virus Genes 2003, 26, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Israeli Ministry of Health. West Nile virus Morbidity Status Report 2016. Available online: https://www.health.gov.il/English/Topics/Diseases/WNF/Pages/StatusReport2016.aspx (accessed on 19 August 2018).
- Malkinson, M.; Banet, C.; Khinich, Y.; Samina, I.; Pokamunski, S.; Weisman, Y. Use of live and inactivated vaccines in the control of West Nile fever in domestic geese. Ann. N. Y. Acad. Sci. 2001, 951, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Ahronson-Ras, K.; Lichter-Peled, A.; Tal, S.; Gelman, B.; Cohen, D.; Klement, E.; Steinman, A. Spatial and temporal distribution of West Nile virus in horses in Israel (1997–2013)—From endemic to epidemics. PLoS ONE 2014, 9, e113149. [Google Scholar]
- Yeruham, I.; Sharir, B.; Yadin, H.; Tiomkin, D.; Chai, D. Bovine ephemeral fever in beef cattle herds in the Jordan Valley, Israel. Vet. Rec. 2003, 152, 86–88. [Google Scholar] [CrossRef] [PubMed]
- Yeruham, I.; Van Ham, M.; Bar, D.; Yadin, H.; Tiomkin, D. Economic aspects of the 1999 outbreak of bovine ephemeral fever in dairy cattle herds in the Jordan Valley in Israel. Vet. Rec. 2003, 153, 180–182. [Google Scholar] [CrossRef]
- Yeruham, I.; Van Ham, M.; Stram, Y.; Friedgut, O.; Yadin, H.; Mumcuoglu, K.Y.; Braverman, Y. Epidemiological investigation of bovine ephemeral Fever outbreaks in Israel. Vet. Med. Int. 2010, 2010, 290541. [Google Scholar] [CrossRef]
- Erster, O.; Stram, R.; Menasherow, S.; Rubistein-Giuni, M.; Sharir, B.; Kchinich, E.; Stram, Y. High-resolution melting (HRM) for genotyping bovine ephemeral fever virus (BEFV). Virus Res. 2017, 229, 1–8. [Google Scholar] [CrossRef]
- Leshem, E.; Bin, H.; Shalom, U.; Perkin, M.; Schwartz, E. Risk for emergence of dengue and chikungunya virus in Israel. Emerg. Infect. Dis. 2012, 18, 345–347. [Google Scholar] [CrossRef]
- Kenawy, M.A.; Abdel-Hamid, Y.M.; Beier, J.C. Rift Valley Fever in Egypt and other African countries: Historical review, recent outbreaks and possibility of disease occurrence in Egypt. Acta Trop. 2018, 181, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Kampen, H.; Medlock, J.; Vaux, A.; Moenraadt, C.; van Vliet, A.; Bartumeus, F.; Oltra, A.; Sousa, C.; Chouin, S.; Werner, D. Approaches to passive mosquito surveillance in the EU. Parasit. Vectors 2015, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, F.; Fischer, D.; Eiden, M.; Reuschel, M.; Müller, K.; Rinder, M.; Urbaniak, S.; Brandes, F.; Schwehn, R.; Lühken, R.; et al. West Nile virus and Usutu virus monitoring of wild birds in Germany. Int. J. Environ. Res. Public Health 2018, 15, E171. [Google Scholar] [CrossRef] [PubMed]
- Scheuch, D.E.; Schäfer, M.; Eiden, M.; Heym, E.C.; Ziegler, U.; Walther, D.; Schmidt-Chanasit, J.; Keller, M.; Groschup, M.H.; Kampen, H. Detection of Usutu, Sindbis, and Batai viruses in mosquitoes (Diptera: Culicidae) collected in Germany, 2011–2016. Viruses 2018, 10, E389. [Google Scholar] [CrossRef] [PubMed]
- Eiden, M.; Ziegler, U.; Keller, M.; Müller, K.; Granzow, H.; Jöst, H.; Schmidt-Chanasit, J.; Groschup, M.H. Isolation of Sindbis virus from a hooded crow in Germany. Vector Borne Zoonotic Dis. 2014, 14, 220–222. [Google Scholar] [CrossRef] [PubMed]
- Cadar, D.; Lühken, R.; Van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Euro Surveill. 2017, 22, 30452. [Google Scholar] [CrossRef] [PubMed]
- ProMed-Mail, Archive Number 20181006.6074497. Available online: https://www.promedmail.org/ (accessed on 6 October 2018).
- Suk, J.E.; Semenza, J.C. Future infectious disease threats to Europe. Am. J. Public Health 2011, 101, 2068–2079. [Google Scholar] [CrossRef] [PubMed]
- Findlater, A.; Bogoch, I.I. Human mobility and the global spread of infectious diseases: A focus on air travel. Trends Parasitol. 2018, 34, 772–783. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Semenza, J.C.; Suk, J.E. Vector-borne diseases and climate change: A European perspective. FEMS Microbiol. Lett. 2018, 365, fnx244. [Google Scholar] [CrossRef]
- Palmer, J.R.B.; Oltra, A.; Collantes, F.; Delgado, J.A.; Lucientes, J.; Delacour, S.; Bengoa, M.; Eritja, R.; Bartumeus, D. Citizen science provides a reliable and scalable tool to track disease-carrying mosquitoes. Nat. Commun. 2017, 8, 916. [Google Scholar] [CrossRef] [PubMed]
- Walther, D.; Kampen, H. The citizen science project ‘Mueckenatlas’ helps monitor the distribution and spread of invasive mosquito species in Germany. J. Med. Entomol. 2017, 54, 1790–1794. [Google Scholar] [CrossRef] [PubMed]
- Cleton, N.B.; Van Maanen, K.; Bergvervoet, S.A.; Bon, N.; Beck, C.; Godeke, G.J.; Lecollinet, S.; Bowen, R.; Lelli, D.; Nowotny, N.; Koopmans, M.P.G.; Reusken, C.B.E.M. A serological protein microarray for detection of multiple cross-reactive flavivirus infections in horses for veterinary and public health surveillance. Transbound. Emerg. Dis. 2017, 64, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.; Desprès, P.; Paulous, S.; Vanhomwegen, J.; Lowenski, S.; Nowotny, N.; Durand, B.; Garnier, A.; Blaise-Boisseau, S.; Guitton, E.; et al. A High-Performance Multiplex Immunoassay for Serodiagnosis of Flavivirus-Associated Neurological Diseases in Horses. Biomed. Res. Int. 2015, 2015, 678084. [Google Scholar] [CrossRef] [PubMed]
- Fros, J.J.; Miesen, P.; Vogels, C.B.; Gaibani, P.; Sambri, V.; Martina, B.E.; Koenraadt, C.J.; Van Rij, R.P.; Vlak, J.M.; Takken, W.; et al. Comparative Usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Health 2015, 1, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Haussig, J.M.; Young, J.J.; Gossner, C.M.; Mezei, E.; Belle, A.; Sirbu, A.; Pervanidou, D.; Drakulovic, M.B.; Sudre, B. Early start of the West Nile fever transmission season 2018 in Europe. Euro Surveill. 2018, 23, 1800428. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | Common Name | Origin |
|---|---|---|
| Aedes albopictus | Asian tiger mosquito | Southeast Asia |
| Aedes aegypti | Yellow fever mosquito | Tropical and subtropical regions |
| Aedes japonicus | Asian bush mosquito | Eastern Asia |
| Aedes atropalpus | American rock pool mosquito | North America |
| Aedes koreicus | None | Korea, Japan, northeast China |
| Virus Family | Genome Structure | Genus | Species |
|---|---|---|---|
| Flaviviridae | Positive strand RNA | Flavivirus | Dengue virus Japanese encephalitis virus |
| Usutu virus | |||
| West Nile virus | |||
| Zika virus | |||
| Peribunyaviridae | Segmented negative strand RNA | Orthobunyavirus | Batai orthobunyavirus |
| Inkoo virus | |||
| Phenuiviridae | Segmented negative strand RNA | Phlebovirus | Rift Valley fever virus |
| Rhabdoviridae | Non-segmented negative strand RNA | Vesiculovirus | Bovine ephemeral fever virus |
| Togaviridae | Positive strand RNA | Alphavirus | Sindbis virus Chikungunya virus |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, N.; Fernández de Marco, M.; Giovannini, A.; Ippoliti, C.; Danzetta, M.L.; Svartz, G.; Erster, O.; Groschup, M.H.; Ziegler, U.; Mirazimi, A.; et al. Emerging Mosquito-Borne Threats and the Response from European and Eastern Mediterranean Countries. Int. J. Environ. Res. Public Health 2018, 15, 2775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15122775
Johnson N, Fernández de Marco M, Giovannini A, Ippoliti C, Danzetta ML, Svartz G, Erster O, Groschup MH, Ziegler U, Mirazimi A, et al. Emerging Mosquito-Borne Threats and the Response from European and Eastern Mediterranean Countries. International Journal of Environmental Research and Public Health. 2018; 15(12):2775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15122775
Chicago/Turabian StyleJohnson, Nicholas, Mar Fernández de Marco, Armando Giovannini, Carla Ippoliti, Maria Luisa Danzetta, Gili Svartz, Oran Erster, Martin H. Groschup, Ute Ziegler, Ali Mirazimi, and et al. 2018. "Emerging Mosquito-Borne Threats and the Response from European and Eastern Mediterranean Countries" International Journal of Environmental Research and Public Health 15, no. 12: 2775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15122775