Nitrogen along the Hydrological Gradient of Marsh Sediments in a Subtropical Estuary: Pools, Processes, and Fluxes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
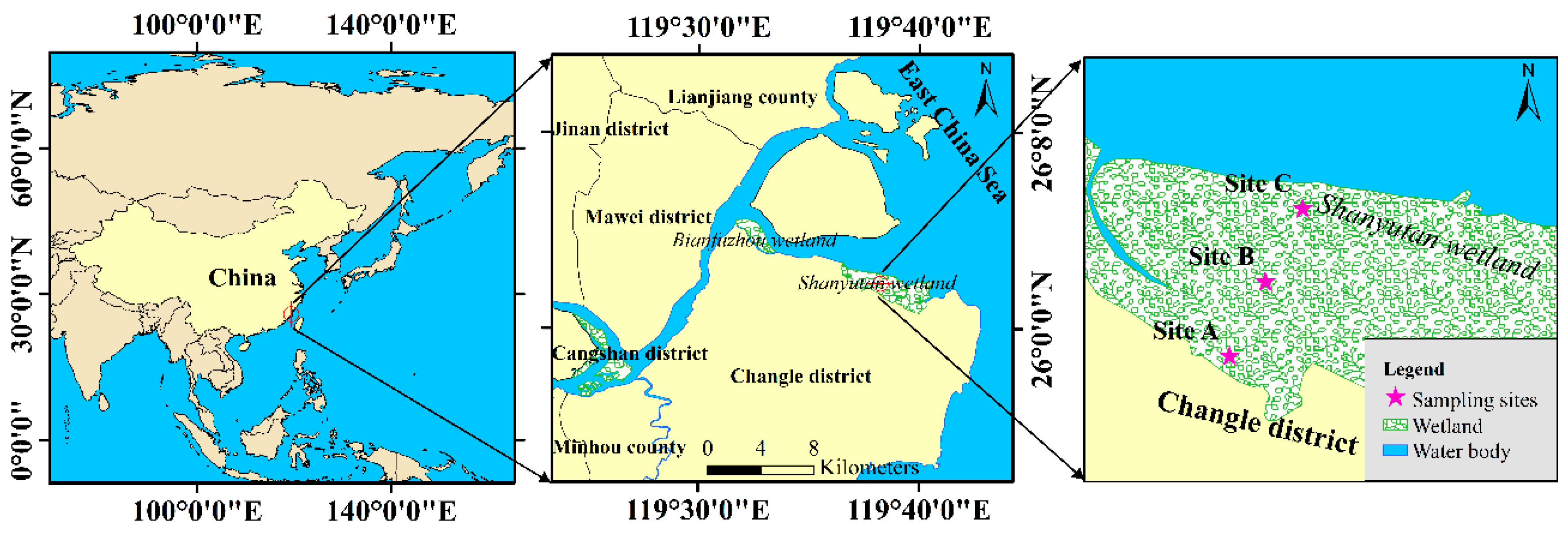
2.1. Study Site
2.2. Sampling and Analysis
2.3. Slurry Incubations
2.4. Dissolved Inorganic Nitrogen Flux Measurements
2.5. Statistical Analyses
3. Results
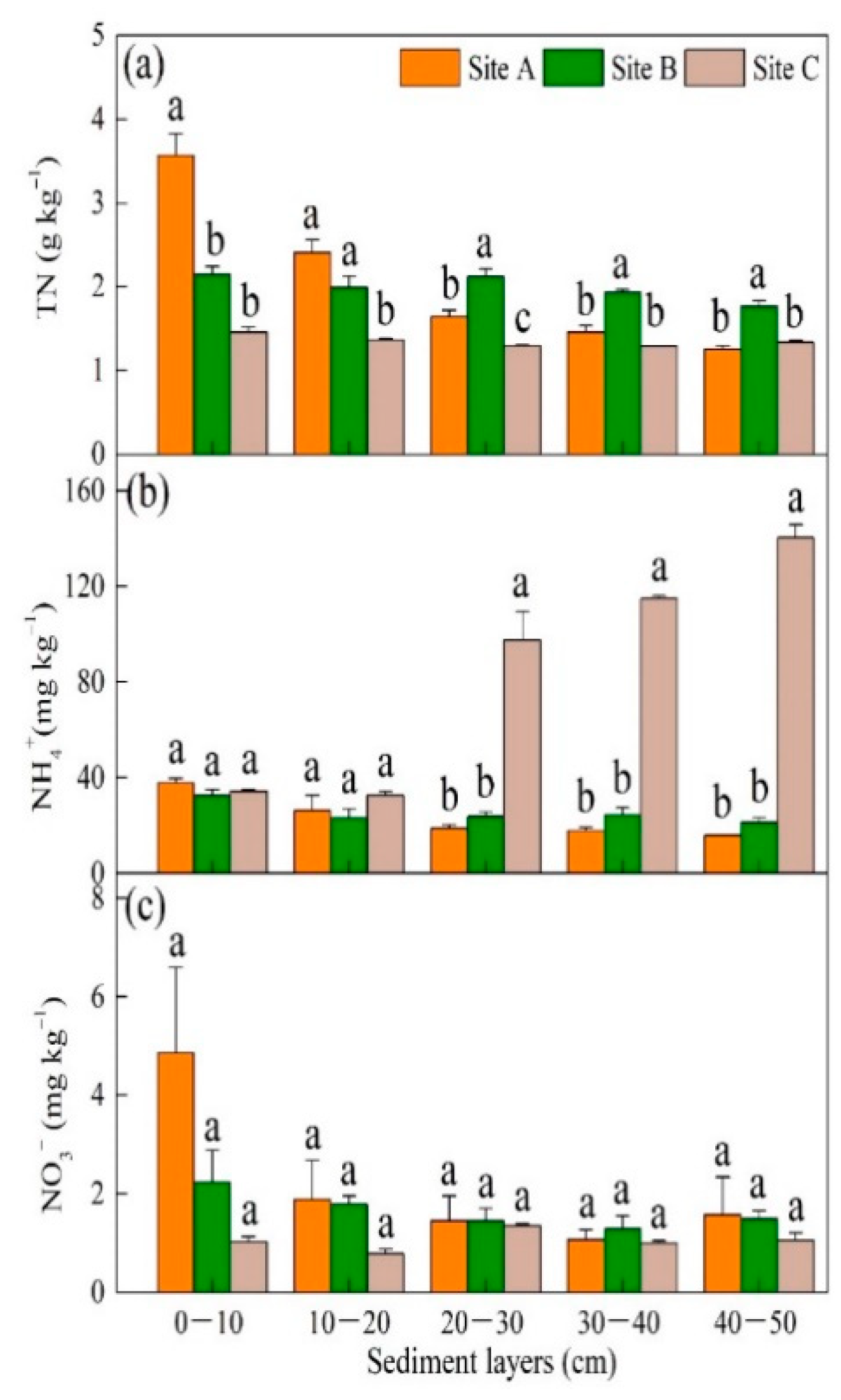
3.1. Plant Biomass and Sediment Geochemistry along a Hydrological Gradient
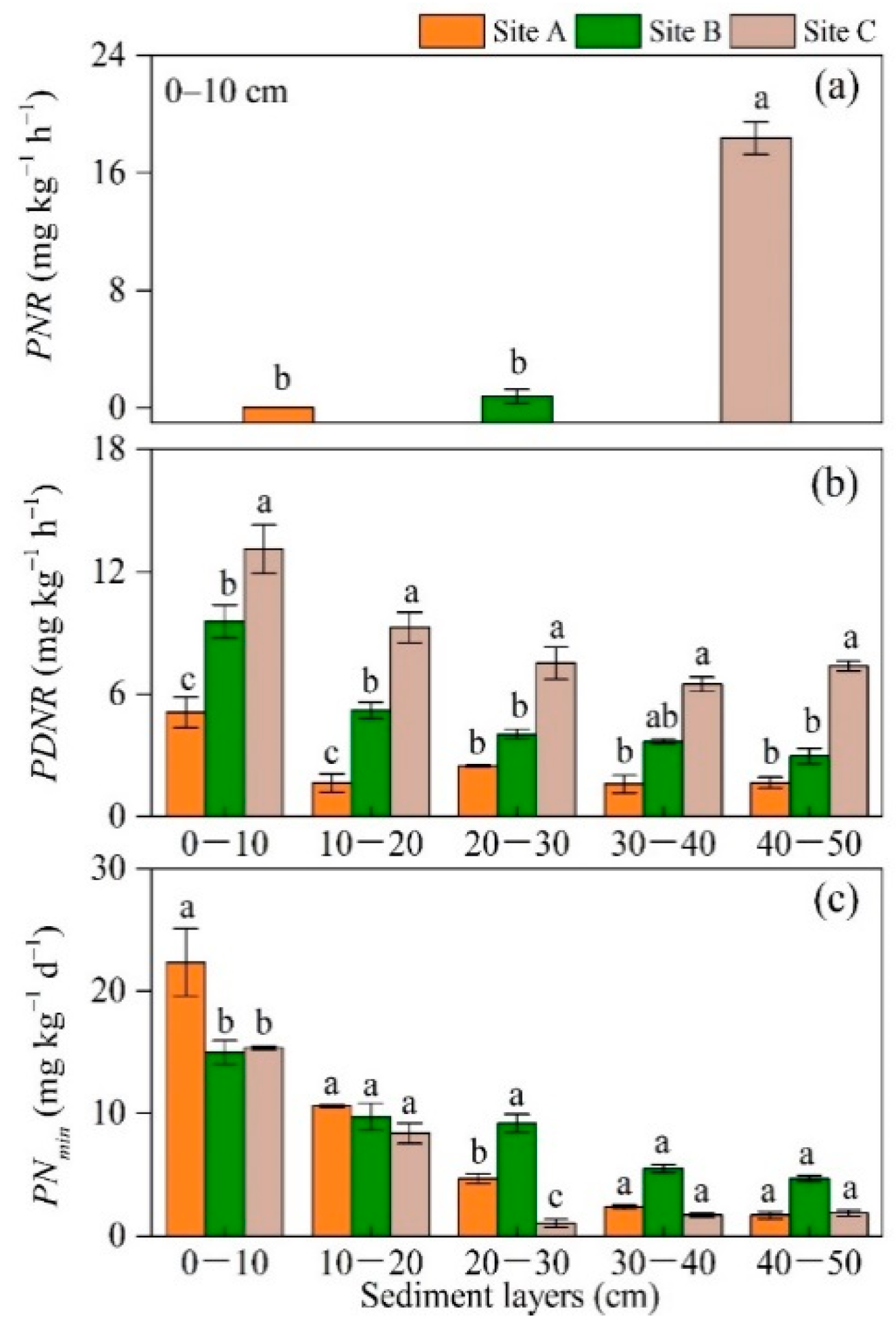
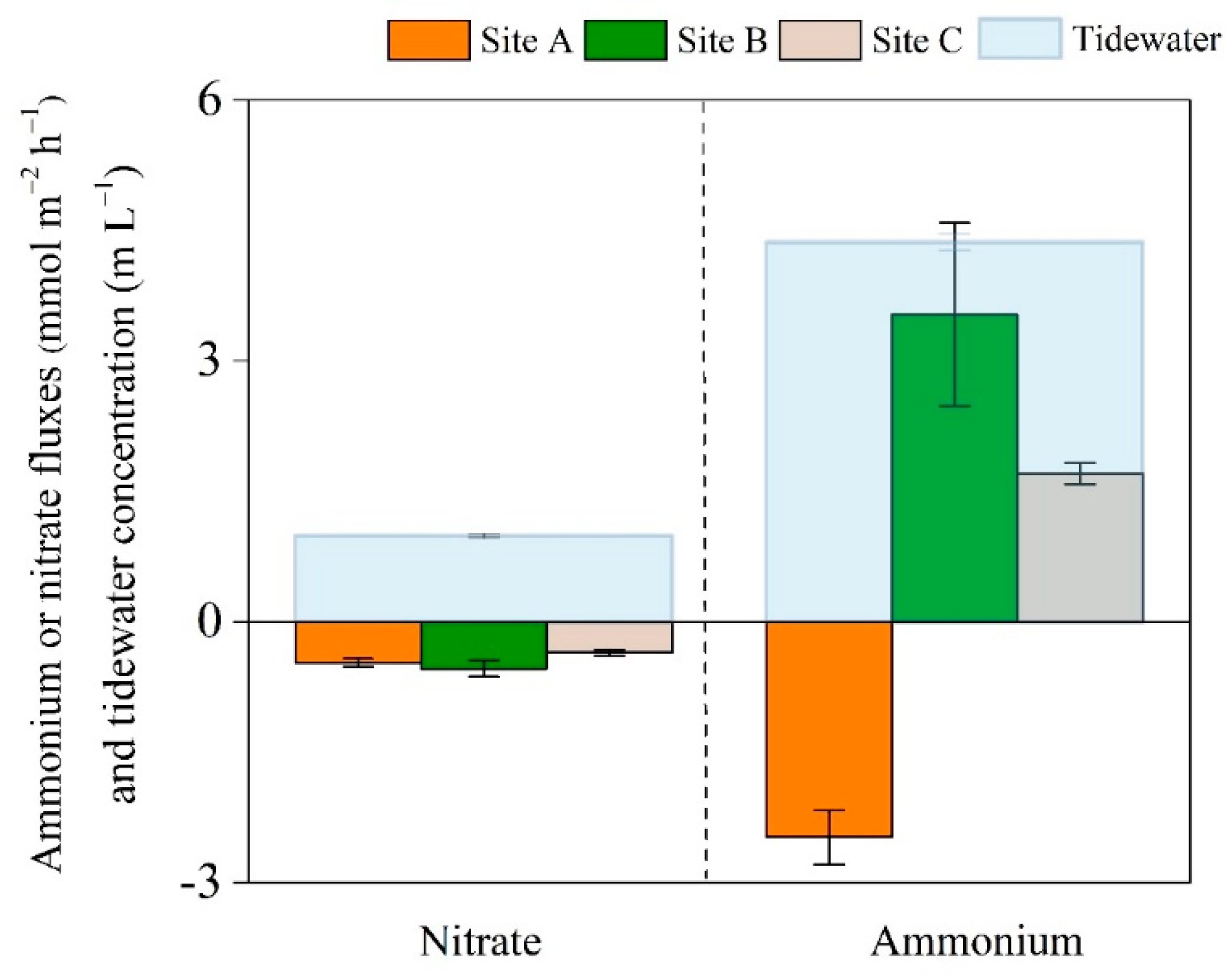
3.2. Distribution of Nitrogen Pools, Processes, and Fluxes
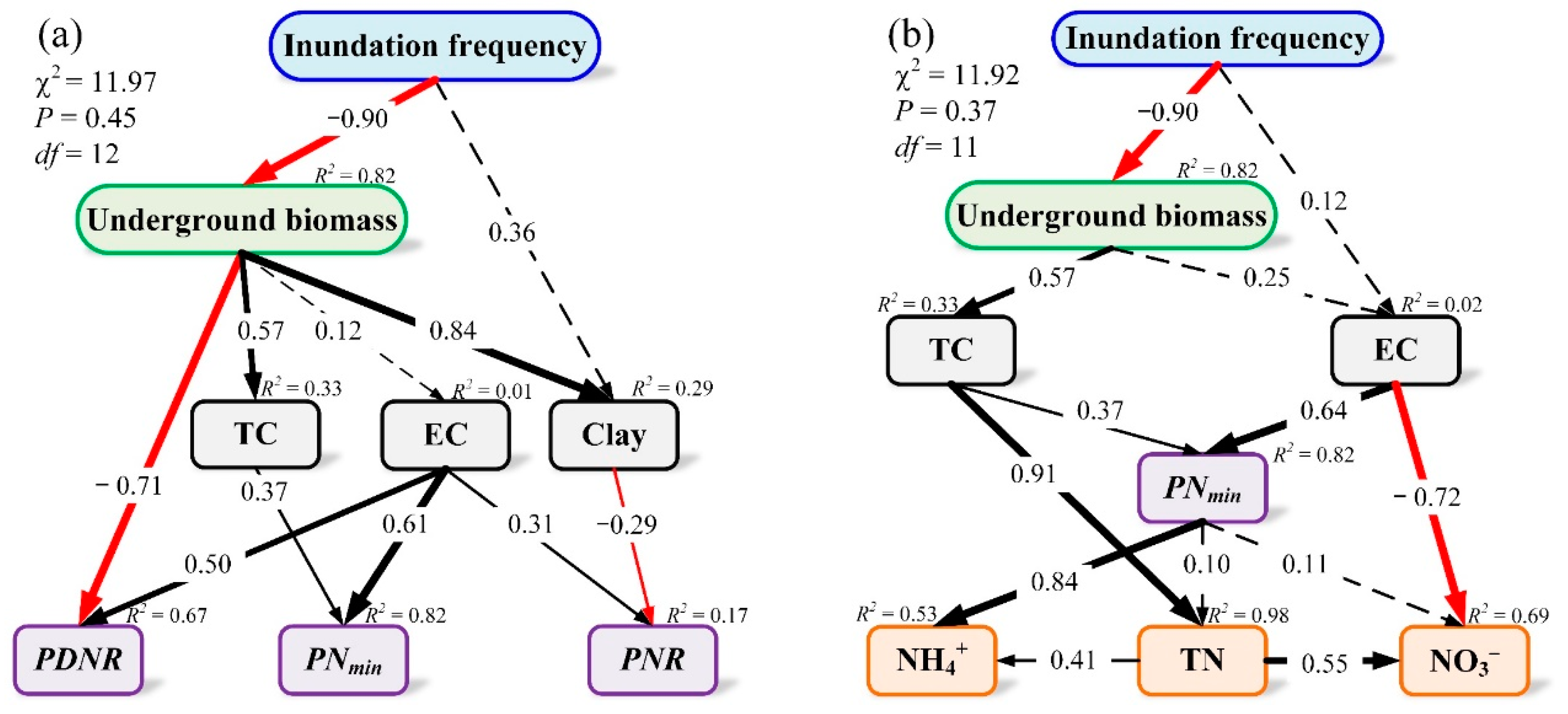
3.3. Structural Equation Modeling Analysis of the Drivers, Causal Links, and Contribution to the Priming Effect
4. Discussion
4.1. Spatial Distribution of Nitrogen Processes
4.2. Spatial Distribution of Nitrogen Pools
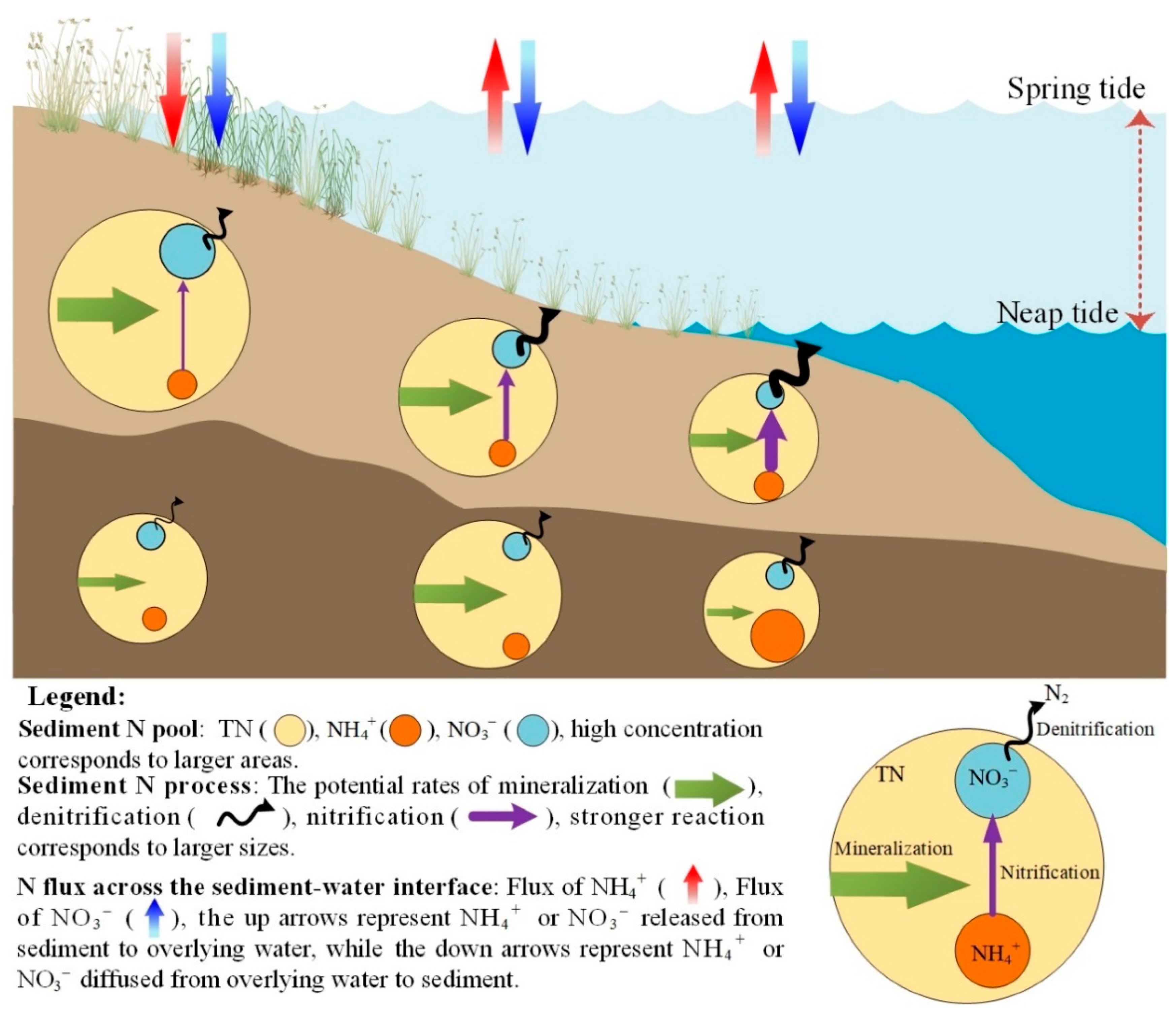
4.3. Spatial Distribution of Nitrogen Fluxes
4.4. Implications, Uncertainties, and Future Study
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hou, L.; Liu, M.; Xu, S.; Ou, D.; Yu, J.; Cheng, S.; Lin, X.; Yang, Y. The effects of semi-lunar spring and neap tidal change on nitrification, denitrification and N2O vertical distribution in the intertidal sediments of the Yangtze estuary, China. Estuar. Coast. Shelf Sci. 2007, 73, 607–616. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Megonigal, J.P. Tidal wetland stability in the face of human impacts and sea-level rise. Nature 2013, 504, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zeng, C.; Tong, C.; Zhai, S.; Lin, X.; Gao, D. Spatial distribution of phosphorus speciation in marsh sediments along a hydrologic gradient in a subtropical estuarine wetland, China. Estuar. Coast. Shelf Sci. 2015, 154, 30–38. [Google Scholar] [CrossRef]
- Wong, J.X.; Van Colen, C.; Airoldi, L. Nutrient levels modify saltmarsh responses to increased inundation in different soil types. Mar. Environ. Res. 2015, 104, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Zeng, C.S.; Tong, C.; Huang, J.F.; Yu, Q.; Guo, Y.B.; Wang, S.-H. Abundance and speciation of iron across a subtropical tidal marsh of the Min River Estuary in the East China Sea. Appl. Geochem. 2014, 45, 1–13. [Google Scholar] [CrossRef]
- Aelion, C.M.; Warttinger, U. Sulfide inhibition of nitrate removal in coastal sediments. Estuar. Coasts 2010, 33, 798–803. [Google Scholar] [CrossRef]
- Deng, F.; Hou, L.; Liu, M.; Zheng, Y.; Yin, G.; Li, X.; Lin, X.; Chen, F.; Gao, J.; Jiang, X. Dissimilatory nitrate reduction processes and associated contribution to nitrogen removal in sediments of the Yangtze Estuary. J. Geophys. Res.-Biogeosci. 2015, 120, 1521–1531. [Google Scholar] [CrossRef]
- Jicha, T.M.; Johnson, L.; Hill, B.; Regal, R.; Elonen, C.; Pearson, M. Spatial and temporal patterns of nitrification rates in forested floodplain wetland soils of upper Mississippi River Pool 8. River Res. Appl. 2013, 30, 650–662. [Google Scholar] [CrossRef]
- Roberts, K.L.; Kessler, A.J.; Grace, M.R.; Cook, P.L. Increased rates of dissimilatory nitrate reduction to ammonium (DNRA) under oxic conditions in a periodically hypoxic estuary. Geochim. Cosmochim. Acta 2014, 133, 313–324. [Google Scholar] [CrossRef]
- Wigand, C.; McKinney, R.A.; Chintala, M.M.; Charpentier, M.A.; Groffman, P.M. Denitrification enzyme activity of fringe salt marshes in New England (USA). J. Environ. Qual. 2004, 33, 1144–1151. [Google Scholar] [CrossRef]
- Cheng, X.; Hou, L.; Liu, M.; Zheng, Y.; Yin, G.; Li, X.; Li, X.; Gao, J.; Deng, F.; Jiang, X. Inorganic nitrogen exchange across the sediment-water interface in the eastern Chongming tidal flat of the Yangtze Estuary. Environ. Earth Sci. 2015, 74, 2173–2184. [Google Scholar] [CrossRef]
- Gao, H.; Bai, J.; Deng, X.; Lu, Q.; Ye, X. Short-term effects of tidal flooding on soil nitrogen mineralization in a Chinese tidal salt marsh. Phys. Chem. Earth 2018, 103, 3–10. [Google Scholar] [CrossRef]
- Salles, J.F.; e Silva, M.C.P.; Dini-Andreote, F.; Dias, A.C.; Guillaumaud, N.; Poly, F.; Van Elsas, J.D. Successional patterns of key genes and processes involved in the microbial nitrogen cycle in a salt marsh chronosequence. Biogeochemistry 2017, 132, 185–201. [Google Scholar] [CrossRef]
- Zhang, L.; Yao, X.; Tang, C.; Xu, H.; Jiang, X.; Zhang, Y. Influence of long-term inundation and nutrient addition on denitrification in sandy wetland sediments from Poyang Lake, a large shallow subtropical lake in China. Environ. Pollut. 2016, 219, 440–449. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Working Group I Contribution to the IPCC Fifth Assessment Report Climate 2013: The Physical Science Basis; IPCC: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Siddiqui, Z.A. Storm surge forecasting for the Arabian Sea. Mar. Geod. 2009, 32, 199–217. [Google Scholar] [CrossRef]
- Quan, W.; Shi, L.; Han, J.; Ping, X.; Shen, A.; Chen, Y. Spatial and temporal distributions of nitrogen, phosphorus and heavy metals in the intertidal sediment of the Chang jiang River Estuary in China. Acta Oceanol. Sin. 2010, 29, 108–115. [Google Scholar] [CrossRef]
- Zhou, J.; Wu, Y.; Kang, Q.; Zhang, J. Spatial variations of carbon, nitrogen, phosphorous and sulfur in the salt marsh sediments of the Yangtze Estuary in China. Estuar. Coast. Shelf Sci. 2007, 71, 47–59. [Google Scholar] [CrossRef]
- Hernandez, M.E.; Mitsch, W.J. Denitrification in created riverine wetlands: Influence of hydrology and season. Ecol. Eng. 2007, 30, 78–88. [Google Scholar] [CrossRef]
- Mchergui, C.; Besaury, L.; Langlois, E.; Aubert, M.; Akpa-Vinceslas, M.; Buatois, B.; Quillet, L.; Bureau, F. A comparison of permanent and fluctuating flooding on microbial properties in an ex-situ estuarine riparian system. Appl. Soil Ecol. 2014, 78, 1–10. [Google Scholar] [CrossRef]
- Song, K.; Lee, S.H.; Mitsch, W.J.; Kang, H. Different responses of denitrification rates and denitrifying bacterial communities to hydrologic pulsing in created wetlands. Soil Biol. Biochem. 2010, 42, 1721–1727. [Google Scholar] [CrossRef]
- Chen, Q.H.; Feng, Y.; Zhang, Y.; Zhang, Q.C.; Shamsi, I.H.; Zhang, Y.; Lin, X. Short-term responses of nitrogen mineralization and microbial community to moisture regimes in greenhouse vegetable soils. Pedosphere 2012, 22, 263–272. [Google Scholar] [CrossRef]
- Jia, J.; Bai, J.; Gao, H.; Wen, X.; Zhang, G.; Cui, B.; Liu, X. In situ soil net nitrogen mineralization in coastal salt marshes (Suaeda salsa) with different flooding periods in a Chinese estuary. Ecol. Indic. 2017, 73, 559–565. [Google Scholar] [CrossRef]
- Lewis, D.B.; Brown, J.A.; Jimenez, K. Effects of flooding and warming on soil organic matter mineralization in Avicennia germinans mangrove forests and Juncus roemerianus salt marshes. Estuar. Coast. Shelf Sci. 2014, 139, 11–19. [Google Scholar] [CrossRef]
- Hansen, J.I.; Henriksen, K.; Blackburn, T.H. Seasonal distribution of nitrifying bacteria and rates of nitrification in coastal marine sediments. Microb. Ecol. 1981, 7, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Domangue, R.J.; Mortazavi, B. Nitrate reduction pathways in the presence of excess nitrogen in a shallow eutrophic estuary. Environ. Pollut. 2018, 238, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Thornton, D.; Nedwell, D.; Underwood, G. Denitrification in sediments of the River Colne estuary, England. Mar. Ecol. Prog. Ser. 2000, 203, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Cabrita, M.T.; Brotas, V. Seasonal variation in denitrification and dissolved nitrogen fluxes in intertidal sediments of the Tagus estuary, Portugal. Mar. Ecol. Prog. Ser. 2000, 202, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Spohn, M.; Babka, B.; Giani, L. Changes in soil organic matter quality during sea-influenced marsh soil development at the North Sea coast. Catena 2013, 107, 110–117. [Google Scholar] [CrossRef]
- Ye, X.; Wang, A.; Chen, J. Distribution and deposition characteristics of carbon and nitrogen in sediments in a semi-closed bay area, southeast China. Cont. Shelf Res. 2014, 90, 133–141. [Google Scholar] [CrossRef]
- Huang, L.; Bai, J.; Xiao, R.; Shi, J.; Gao, H. The soil nitrogen dynamics in an inland salt marsh as affected by various experimental water levels. Hydrol. Process. 2014, 28, 4708–4717. [Google Scholar] [CrossRef]
- Laursen, A.E.; Seitzinger, S.P. The role of denitrification in nitrogen removal and carbon mineralization in Mid-Atlantic Bight sediments. Cont. Shelf Res. 2002, 22, 1397–1416. [Google Scholar] [CrossRef]
- Risgaard-Petersen, N.; Meyer, R.L.; Schmid, M.; Jetten, M.S.; Enrich-Prast, A.; Rysgaard, S.; Revsbech, N.P. Anaerobic ammonium oxidation in an estuarine sediment. Aquat. Microb. Ecol. 2004, 36, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Eyre, B.D.; Pont, D. Intra- and inter-annual variability in the different forms of diffuse nitrogen and phosphorus delivered to seven sub-tropical east Australian estuaries. Estuar. Coast. Shelf Sci. 2003, 57, 137–148. [Google Scholar] [CrossRef]
- Eyre, B.D. Regional evaluation of nutrient transformation and phytoplankton growth in nine river-dominated sub-tropical east Australian estuaries. Mar. Ecol. Prog. Ser. 2000, 205, 61–83. [Google Scholar] [CrossRef]
- Zhang, W.; Shi, F.; Hong, H.; Shang, S.; Kirby, J.T. Tide-surge interaction intensified by the Taiwan Strait. J. Geophys. Res.-Oceans 2010, 115, 1–17. [Google Scholar] [CrossRef]
- Hu, M.; Wilson, B.J.; Sun, Z.; Ren, P.; Tong, C. Effects of the addition of nitrogen and sulfate on CH4 and CO2 emissions, soil, and pore water chemistry in a high marsh of the Min River estuary in southeastern China. Sci. Total Environ. 2016, 579, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.F.; Nedwell, D.B.; Stott, A. Sources of nitrogen used for denitrification and nitrous oxide formation in sediments of the hypernutrified Colne, the nutrified Humber, and the oligotrophic Conwy estuaries, United Kingdom. Limnol. Oceanogr. 2006, 51, 545–557. [Google Scholar] [CrossRef]
- Luo, M.; Zhu, W.; Huang, J.; Liu, Y.; Duan, X.; Wu, J.; Tong, C. Anaerobic organic carbon mineralization in tidal wetlands along a low-level salinity gradient of a subtropical estuary: Rates, pathways, and controls. Geoderma 2019, 337, 1245–1257. [Google Scholar] [CrossRef]
- Bai, J.; Wang, Q.; Deng, W.; Gao, H.; Tao, W.; Xiao, R. Spatial and seasonal distribution of nitrogen in marsh soils of a typical floodplain wetland in Northeast China. Environ. Monit. Assess. 2012, 184, 1253–1263. [Google Scholar] [CrossRef]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Burger, M.; Jackson, L.E. Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems. Soil Biol. Biochem. 2003, 35, 29–36. [Google Scholar] [CrossRef]
- Barnes, J.; Owens, N. Denitrification and nitrous oxide concentrations in the Humber estuary, UK, and adjacent coastal zones. Mar. Pollut. Bull. 1999, 37, 247–260. [Google Scholar] [CrossRef]
- Magalhães, C.M.; Joye, S.B.; Moreira, R.M.; Wiebe, W.J.; Bordalo, A.A. Effect of salinity and inorganic nitrogen concentrations on nitrification and denitrification rates in intertidal sediments and rocky biofilms of the Douro River estuary, Portugal. Water Res. 2005, 39, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Fornara, D.A.; Tilman, D.; Hobbie, S.E. Linkages between plant functional composition, fine root processes and potential soil N mineralization rates. J. Ecol. 2009, 97, 48–56. [Google Scholar] [CrossRef]
- Clavero, V.; Izquierdo, J.J.; Fernandez, J.A.; Niell, F.X. Influence of bacterial density on the exchange of phosphate between sediment and overlying water. Hydrobiologia 1999, 392, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Ge, T.; Luo, Y.; Liu, S.; Xu, X.; Tong, C.; Shibistova, O.; Guggenberger, G.; Wu, J. Microbial stoichiometric flexibility regulates rice straw mineralization and its priming effect in paddy soil. Soil Biol. Biochem. 2018, 121, 67–76. [Google Scholar] [CrossRef]
- Xiao, K.; Wu, J.; Li, H.; Hong, Y.; Wilson, A.M.; Jiao, J.J.; Shananan, M. Nitrogen fate in a subtropical mangrove swamp: Potential association with seawater-groundwater exchange. Sci. Total Environ. 2018, 635, 586–597. [Google Scholar] [CrossRef]
- Marchant, H.K.; Holtappels, M.; Lavik, G.; Ahmerkamp, S.; Winter, C.; Kuypers, M.M. Coupled nitrification-denitrification leads to extensive N loss in subtidal permeable sediments. Limnol. Oceanogr. 2016, 61, 1033–1048. [Google Scholar] [CrossRef] [Green Version]
- Devol, A.H.; Christensen, J.P. Benthic fluxes and nitrogen cycling in sediments of the continental margin of the eastern North Pacific. J. Mar. Res. 1993, 51, 345–372. [Google Scholar] [CrossRef]
- Devol, A.H.; Codispoti, L.A.; Christensen, J.P. Summer and winter denitrification rates in western Arctic shelf sediments. Cont. Shelf Res. 1997, 17, 1029–1050. [Google Scholar] [CrossRef]
- Rysgaard, S.; Thamdrup, B.; Risgaard-Petersen, N.; Fossing, H.; Berg, P.; Christensen, P.B.; Dalsgaard, T. Seasonal carbon and nutrient mineralization in a high-Arctic coastal marine sediment, Young Sound, Northeast Greenland. Mar. Ecol. Prog. Ser. 1998, 175, 261–276. [Google Scholar] [CrossRef] [Green Version]
- Wrage, N.; Velthof, G.; Van Beusichem, M.; Oenema, O. Role of nitrifier denitrification in the production of nitrous oxide. Soil Biol. Biochem. 2001, 33, 1723–1732. [Google Scholar] [CrossRef]
- Watson, E.B.; Wigand, C.; Davey, E.W.; Andrews, H.M.; Bishop, J.; Raposa, K.B. Wetland loss patterns and inundation-productivity relationships prognosticate widespread salt marsh loss for southern New England. Estuar. Coasts 2017, 40, 662–681. [Google Scholar] [CrossRef] [PubMed]
- Holmer, M.; Gribsholt, B.; Kristensen, E. Effects of sea level rise on growth of Spartina anglica and oxygen dynamics in rhizosphere and salt marsh sediments. Mar. Ecol. Prog. Ser. 2002, 225, 197–204. [Google Scholar] [CrossRef]
- Duan, J.; Fang, H.; Su, B.; Chen, J.; Lin, J. Characterization of a halophilic heterotrophic nitrification-aerobic denitrification bacterium and its application on treatment of saline wastewater. Bioresour. Technol. 2015, 179, 421–428. [Google Scholar] [CrossRef]
- Sánchez-García, M.; Alburquerque, J.A.; Sánchez-Monedero, M.A.; Roig, A.; Cayuela, M.L. Biochar accelerates organic matter degradation and enhances N mineralisation during composting of poultry manure without a relevant impact on gas emissions. Bioresour. Technol. 2015, 192, 272–279. [Google Scholar] [CrossRef]
- Crews, T.E.; Kurina, L.M.; Vitousek, P.M. Organic matter and nitrogen accumulation and nitrogen fixation during early ecosystem development in Hawaii. Biogeochemistry 2001, 52, 259–279. [Google Scholar] [CrossRef]
- Tobias, C.R.; Anderson, I.C.; Canuel, E.A.; Macko, S.A. Nitrogen cycling through a fringing marsh-aquifer ecotone. Mar. Ecol. Prog. Ser. 2001, 210, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Yando, E.S.; Osland, M.J.; Willis, J.M.; Day, R.H.; Krauss, K.W.; Hester, M.W. Salt marsh-mangrove ecotones: Using structural gradients to investigate the effects of woody plant encroachment on plant-soil interactions and ecosystem carbon pools. J. Ecol. 2016, 104, 1020–1031. [Google Scholar] [CrossRef]
- Lin, X.; Hou, L.; Liu, M.; Li, X.; Zheng, Y.; Yin, G.; Gao, J.; Jiang, X. Nitrogen mineralization and immobilization in sediments of the East China Sea: Spatiotemporal variations and environmental implications. J. Geophys. Res.-Biogeosci. 2016, 121, 2842–2855. [Google Scholar] [CrossRef]
- Ardón, M.; Morse, J.L.; Colman, B.P.; Bernhardt, E.S. Drought-induced saltwater incursion leads to increased wetland nitrogen export. Glob. Chang. Biol. 2013, 19, 2976–2985. [Google Scholar] [CrossRef] [PubMed]
- Lamers, L.P.; Tomassen, H.B.; Roelofs, J.G. Sulfate-induced eutrophication and phytotoxicity in freshwater wetlands. Environ. Sci. Technol. 1998, 32, 199–205. [Google Scholar]
- Kroeger, K.; Charette, M. Nitrogen biogeochemistry of submarine groundwater discharge. Limnol. Oceanogr. 2008, 53, 1025–1039. [Google Scholar] [CrossRef]
- Weston, N.B.; Giblin, A.E.; Banta, G.T.; Hopkinson, C.S.; Tucker, J. The effects of varying salinity on ammonium exchange in estuarine sediments of the Parker River, Massachusetts. Estuar. Coasts 2010, 33, 985–1003. [Google Scholar]
- Hou, L.; Liu, M.; Jiang, H.; Xu, S.; Ou, D.; Liu, Q.; Zhang, B. Ammonium adsorption by tidal flat surface sediments from the Yangtze Estuary. Environ. Geol. 2003, 45, 72–78. [Google Scholar]
- Zhou, M.; Butterbach-Bahl, K.; Vereecken, H.; Brüggemann, N. A meta-analysis of soil salinization effects on nitrogen pools, cycles and fluxes in coastal ecosystems. Glob. Chang. Biol. 2017, 23, 1338–1352. [Google Scholar] [CrossRef]
- Cormier, N.; Krauss, K.W.; Conner, W.H. Periodicity in stem growth and litterfall in tidal freshwater forested wetlands: Influence of salinity and drought on nitrogen recycling. Estuar. Coasts 2013, 36, 533–546. [Google Scholar] [CrossRef]
- Helali, M.A.; Zaaboub, N.; Oueslati, W.; Added, A.; Aleya, L. Nutrient exchange and oxygen demand at the sediment-water interface during dry and wet seasons off the Medjerda River Delta (Tunis Gulf, Tunisia). Environ. Earth Sci. 2016, 75, 1–25. [Google Scholar] [CrossRef]
- Hanan, E.J.; Schimel, J.P.; Dowdy, K.; D’Antonio, C.M. Effects of substrate supply, pH, and char on net nitrogen mineralization and nitrification along a wildfire-structured age gradient in chaparral. Soil Biol. Biochem. 2016, 95, 87–99. [Google Scholar] [CrossRef]
- Zhao, Y.; Shan, B.; Tang, W.; Zhang, H. Nitrogen mineralization and geochemical characteristics of amino acids in surface sediments of a typical polluted area in the Haihe River Basin, China. Environ. Sci. Pollut. Res. 2015, 22, 17975–17986. [Google Scholar] [CrossRef]
- Lin, X.; McCarthy, M.J.; Carini, S.A.; Gardner, W.S. Net, actual, and potential sediment-water interface NH4+ fluxes in the northern Gulf of Mexico (NGOMEX): Evidence for NH4+ limitation of microbial dynamics. Cont. Shelf Res. 2011, 31, 120–128. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nitrogen Process | Location | Depth (cm) | Value | Distribution | References | |
|---|---|---|---|---|---|---|
| N pool | TN (g kg−1) | Yangtze Estuary, China | 0–10, 40–100 | 1.4–7.8 | H > M ≈ L | [18] |
| 10–40 | H ≈ M > L | |||||
| Yangtze Estuary, China | 0–15 | – | H ≥ M ≈ L | [17] | ||
| Petersgroden, Cäciliengroden, and Neβmersiel Germany | 0–15 | 2.6–6.8 | H > M > L | [29] | ||
| Nordschweiburg and Dangast, Germany | 0–15 | 3.1–4.2 | M > L ≥ H | [29] | ||
| Luoyuan Bay, China | 0–40 | 5.4–10.1 | H > M > L | [30] | ||
| 40–131 | M ≈ L > H | [30] | ||||
| NH4+ (mg kg−1) | Tagus Estuary, Portugal | 0–5 | 0.004–4.1 | L > H | [28] | |
| Yellow River Delta, China | 0–10 | 1.0–6.5 | H ≈ M ≈ L | [23] | ||
| NO3− (mg kg−1) | Tagus Estuary, Portugal | 0–5 | 0.03–2.5 | L > H | [28] | |
| Yellow River Delta, China | 0–10 | 0.7–8.4 | H ≥ M ≈ L | [23] | ||
| N process | PNR | East coast of Jutland, Denmark | 0–8 | – | H > L | [25] |
| PNmin (mg kg−1 d−1) | Yellow River Delta, China | 0–10 | −0.23–0.24 | L ≥ M ≥ H | [23] | |
| PDNR (μmol N m−2 h−1) | Colne Estuary, UK | 0–10 | 1.1–98.2 | H ≈ L | [27] | |
| Mid-Atlantic Bight, North Atlantic Ocean | 0–8 | 0.006–0.2 | W17m > W15m ≈ W11m | [32] | ||
| Randers Fjord and Norsminde Fjord, Denmark | 0–0.5 | – | W1m > W0.5m | [33] | ||
| Colne Estuary and Humber Estuary, UK | 0–2 | 0.1–421.7 | H > M > L | [38] | ||
| Conwy Estuary, UK | 0–2 | 0–108 | M > H > L | [38] | ||
| Weeks Bay Estuarine, USA | 0–19 | 21.6–33.6 | M ≈ L | [26] | ||
| Inorganic N fluxes at the sediment–water interface. | Flux of NO3− (mmol m−2 h−1) | Tagus Estuary, Portugal | – | −0.8–1.8 | H (–), L (+) | [28] |
| Mid-Atlantic Bight, North Atlantic Ocean | – | −0.01–0.02 | W11m (±), W15m (±), W17m (–) | [32] | ||
| Randers Fjord and Norsminde Fjord, Denmark | – | −120.0–47.0 | W1m (–) > W0.5m (–) | [33] | ||
| Yangtze Estuary, China | – | −0.8–0.4 | H (±), I (±), M (±) | [11] | ||
| Flux of NH4+ (mmol m−2 h−1) | Tagus Estuary, Portugal | – | 0.02–0.09 | H (+), L (+) | [28] | |
| Mid-Atlantic Bight, North Atlantic Ocean | – | −0.02–0.12 | W11m (±), W15m (+), W17m (+) | [32] | ||
| Yangtze Estuary, China | −0.2–0.6 | H (±), I (±), M (±) | [11] | |||
| Depth (cm) | Sites | BD (g cm−3) | pH (1:5, Soil:H2O) | EC (mS cm−1) | TC (mg g−1) | Grain Size (%) | ||
|---|---|---|---|---|---|---|---|---|
| Clay | Silt | Sand | ||||||
| 0–10 | Site A | 0.71 ab | 5.88 c | 3.58 a | 39.36 a | 26.94 a | 68.54 a | 4.53 a |
| Site B | 0.75 a | 6.45 b | 3.35 a | 24.56 b | 29.51 a | 69.59 a | 0.90 b | |
| Site C | 0.63 b | 7.32 a | 3.29 a | 15.70 c | 20.40 a | 75.28 a | 4.32 a | |
| 10–20 | Site A | 0.89 a | 6.24 a | 2.48 a | 27.89 a | 34.51 a | 63.61 b | 1.88 a |
| Site B | 0.70 b | 6.50 a | 3.08 a | 21.34 b | 31.52 ab | 66.73 ab | 1.75 a | |
| Site C | 0.75 ab | 7.17 a | 3.04 a | 15.37 c | 26.08 b | 71.12 a | 2.81 a | |
| 20–30 | Site A | 0.95 b | 6.60 ab | 2.13 b | 18.49 b | 34.98 a | 63.55 b | 1.46 a |
| Site B | 0.81 a | 6.41 b | 2.96 a | 22.56 a | 34.54 ab | 64.18 ab | 1.28 a | |
| Site C | 0.85 ab | 7.46 a | 2.07 b | 15.01 c | 26.66 b | 70.59 a | 2.76 a | |
| 30–40 | Site A | 1.01 a | 6.81 b | 1.89 b | 17.74 b | 30.98 a | 65.99 b | 3.04 a |
| Site B | 0.73 b | 6.44 b | 2.65 a | 20.81 a | 32.14 ab | 66.48 ab | 1.38 a | |
| Site C | 0.84 ab | 7.66 a | 1.90 b | 14.69 c | 26.96 b | 70.52 a | 2.52 a | |
| 40–50 | Site A | 1.03 a | 6.58 b | 2.00 b | 16.29 b | 25.92 a | 67.01 b | 7.08 a |
| Site B | 0.80 b | 6.65 b | 2.54 a | 20.93 a | 27.39 a | 69.88 ab | 2.73 a | |
| Site C | 0.82 b | 7.66 a | 2.00 b | 14.58 b | 23.95 a | 72.93 a | 3.12 a | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, W.; Zhang, W.; Zhang, L.; Tong, C.; Sun, Z.; Chen, Y.; Zeng, C. Nitrogen along the Hydrological Gradient of Marsh Sediments in a Subtropical Estuary: Pools, Processes, and Fluxes. Int. J. Environ. Res. Public Health 2019, 16, 2043. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16112043
Hu W, Zhang W, Zhang L, Tong C, Sun Z, Chen Y, Zeng C. Nitrogen along the Hydrological Gradient of Marsh Sediments in a Subtropical Estuary: Pools, Processes, and Fluxes. International Journal of Environmental Research and Public Health. 2019; 16(11):2043. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16112043
Chicago/Turabian StyleHu, Weifang, Wenlong Zhang, Linhai Zhang, Chuan Tong, Zhigao Sun, Yuehmin Chen, and Congsheng Zeng. 2019. "Nitrogen along the Hydrological Gradient of Marsh Sediments in a Subtropical Estuary: Pools, Processes, and Fluxes" International Journal of Environmental Research and Public Health 16, no. 11: 2043. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16112043