Degradation Characteristics of Phosphorus in Phytoplankton-Derived Particulate Organic Matter and Its Effects on the Growth of Phosphorus-Deficient Microcystis aeruginosa in Lake Taihu


Abstract
:1. Introduction
2. Materials and Method
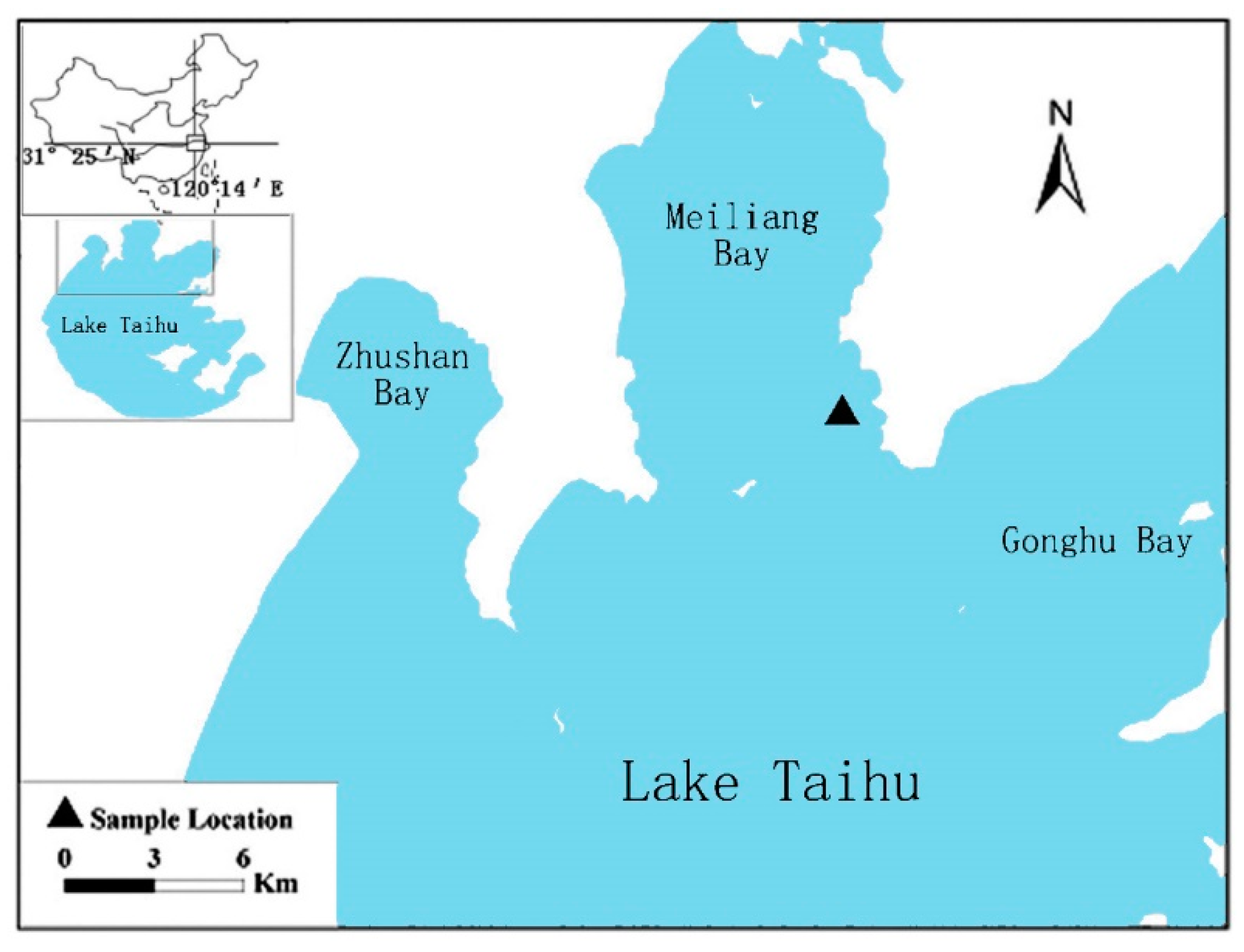
2.1. Study Area
2.2. Experimental Design
2.2.1. Simulation Experiment on Degradation of PPOM
2.2.2. Effects of PPOM Degradation on the Growth of Microcystis Aeruginosa
2.3. Sample Analysis
2.3.1. Physical and Chemical Index Analysis
2.3.2. Molecular Particulate P Analysis by 31P Nuclear Magnetic Resonance
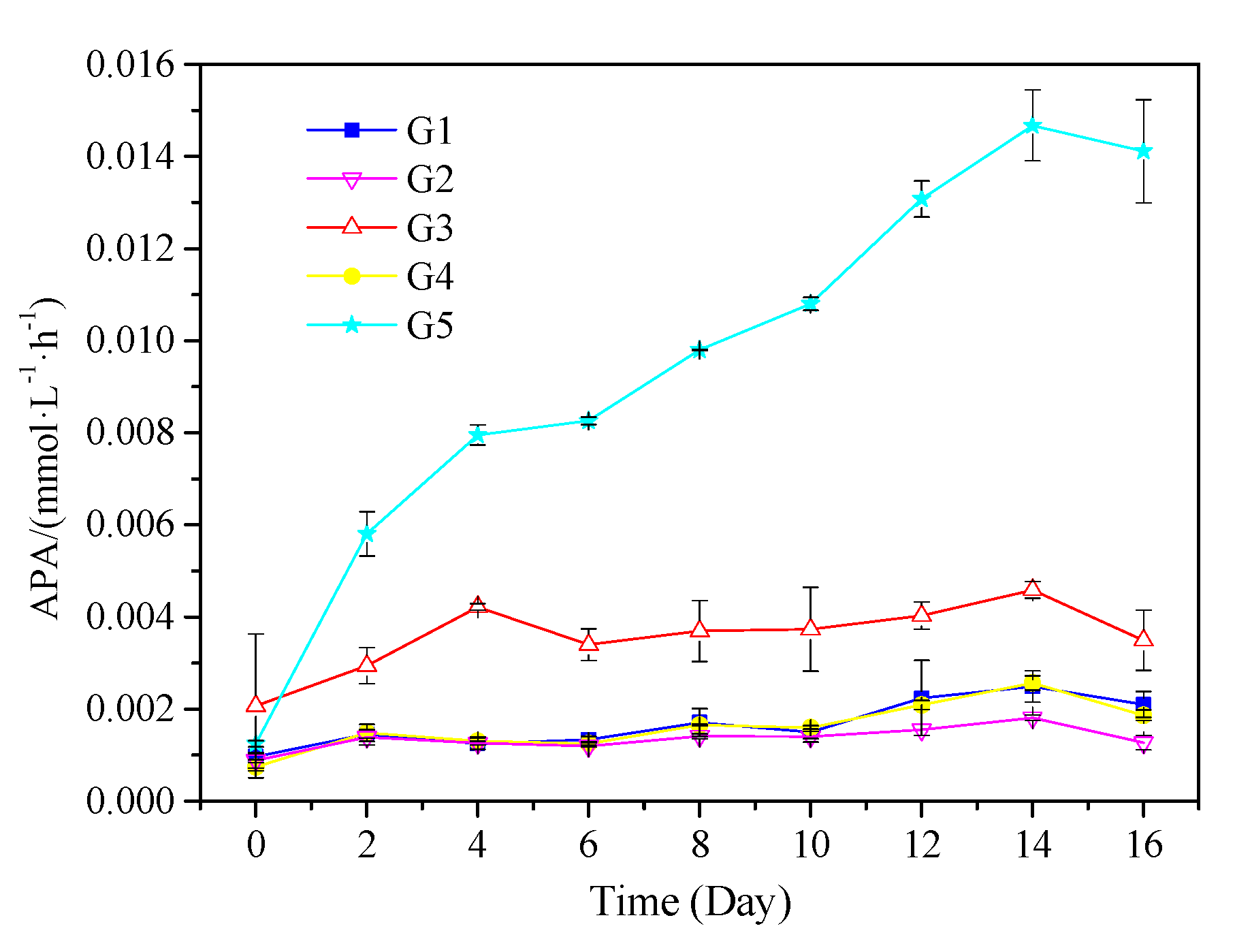
2.3.3. Alkaline Phosphatase Activity in Particulate Matter and Water
2.4. Statistical Analysis
3. Results
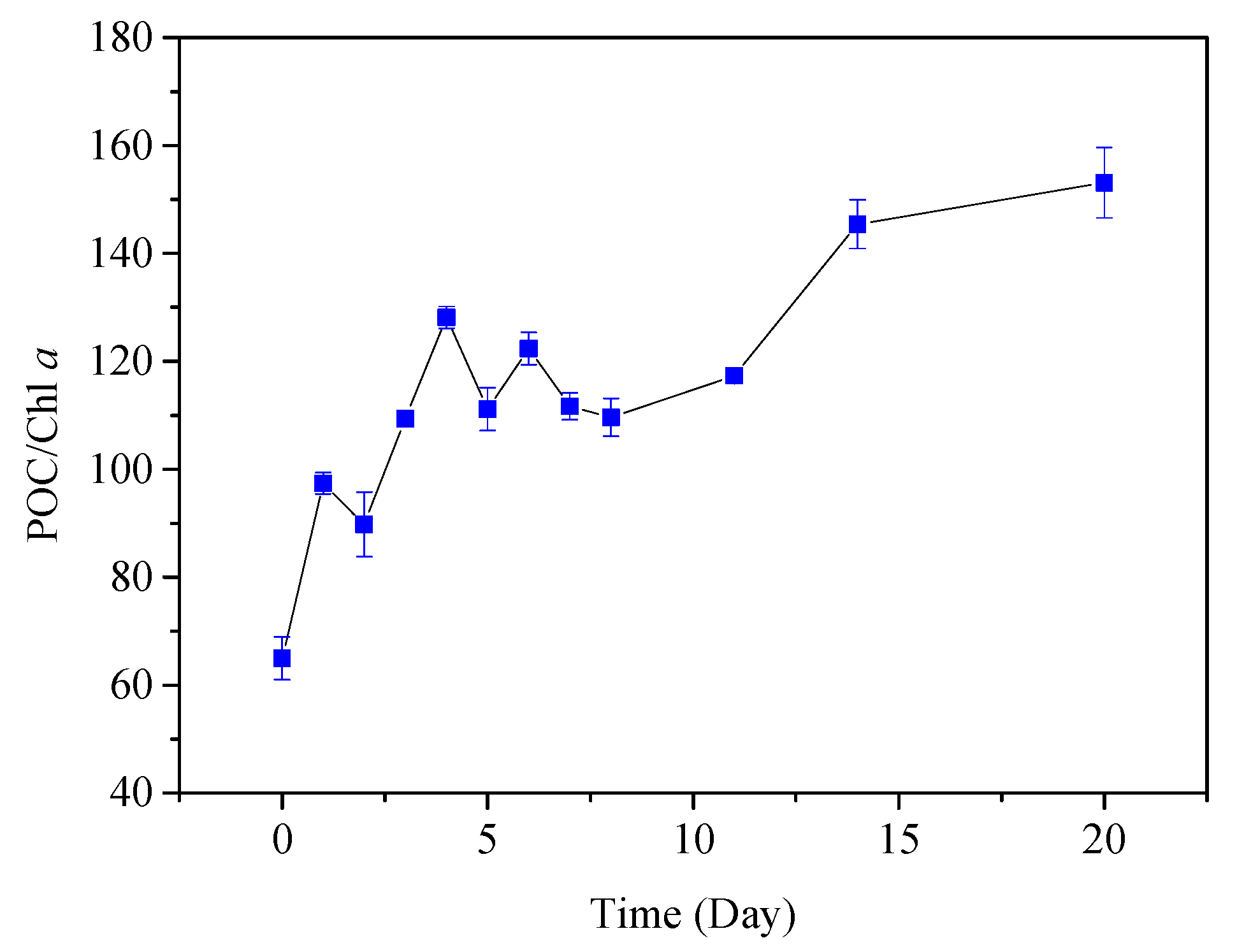
3.1. General Characteristics of Water and PPOM
3.2. Degradation Characteristics of PPOM
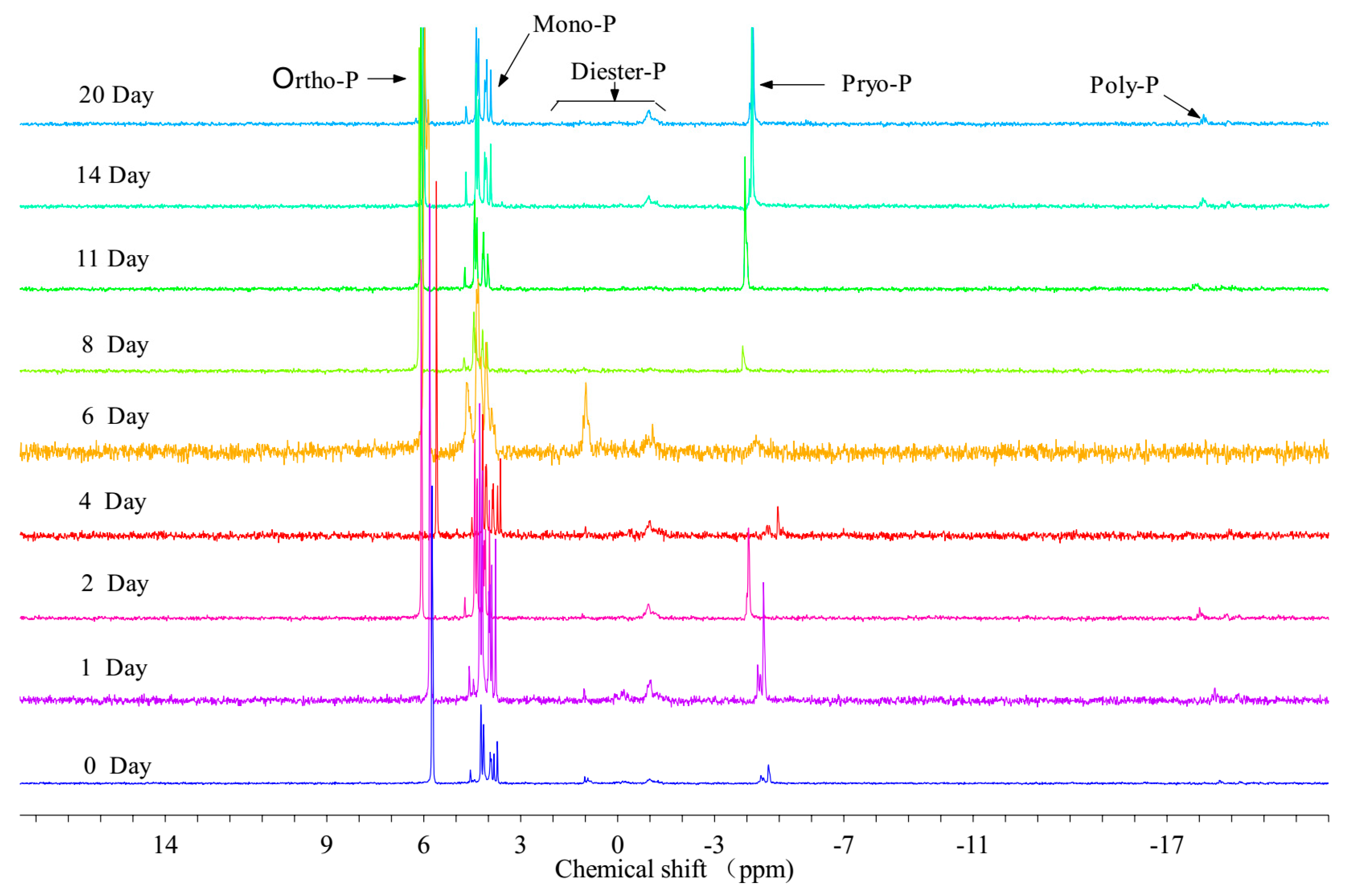
3.2.1. Analysis of P Species in PPOM by 31P NMR
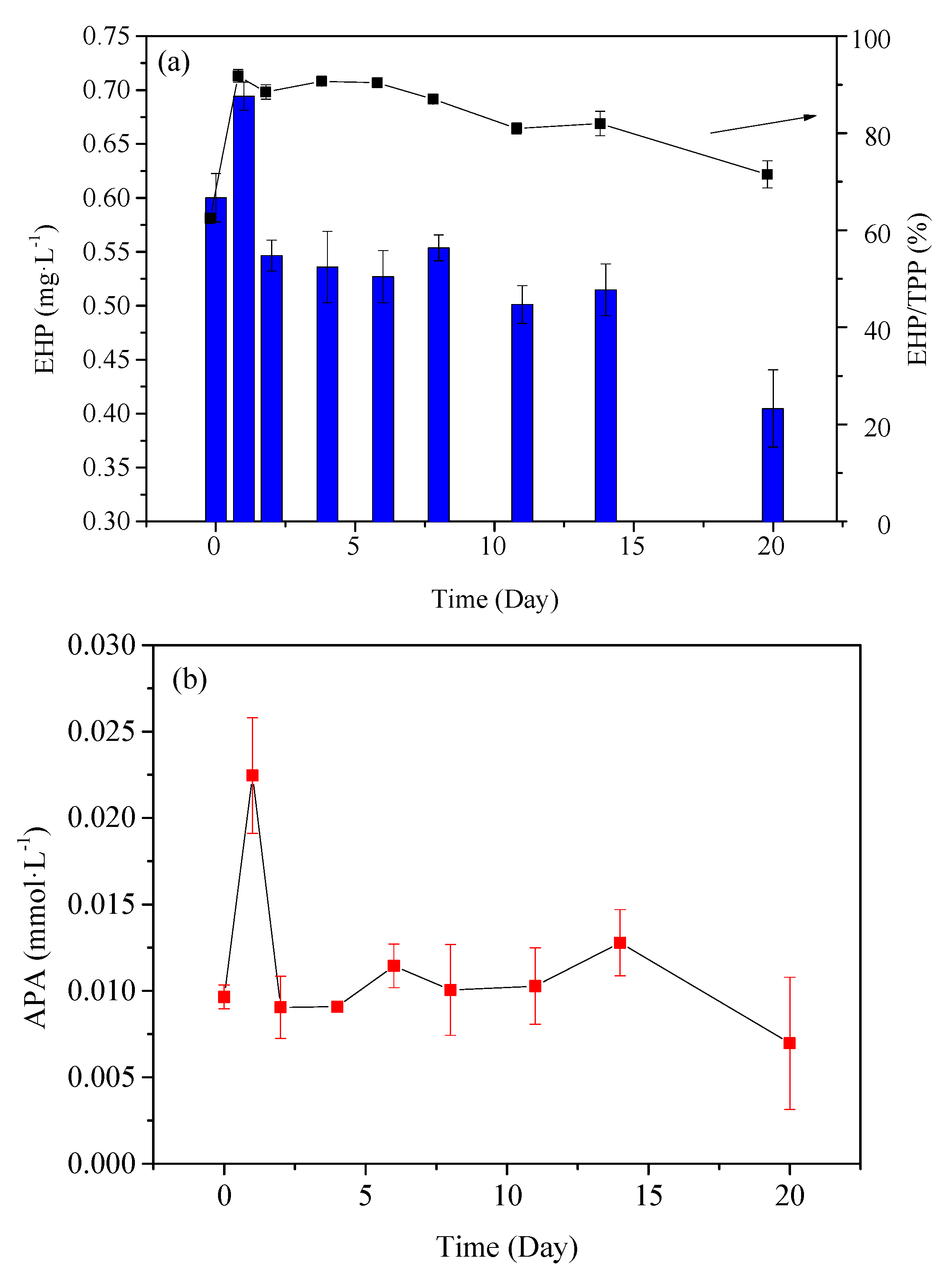
3.2.2. Analysis of P Species by Enzymatic Hydrolysis
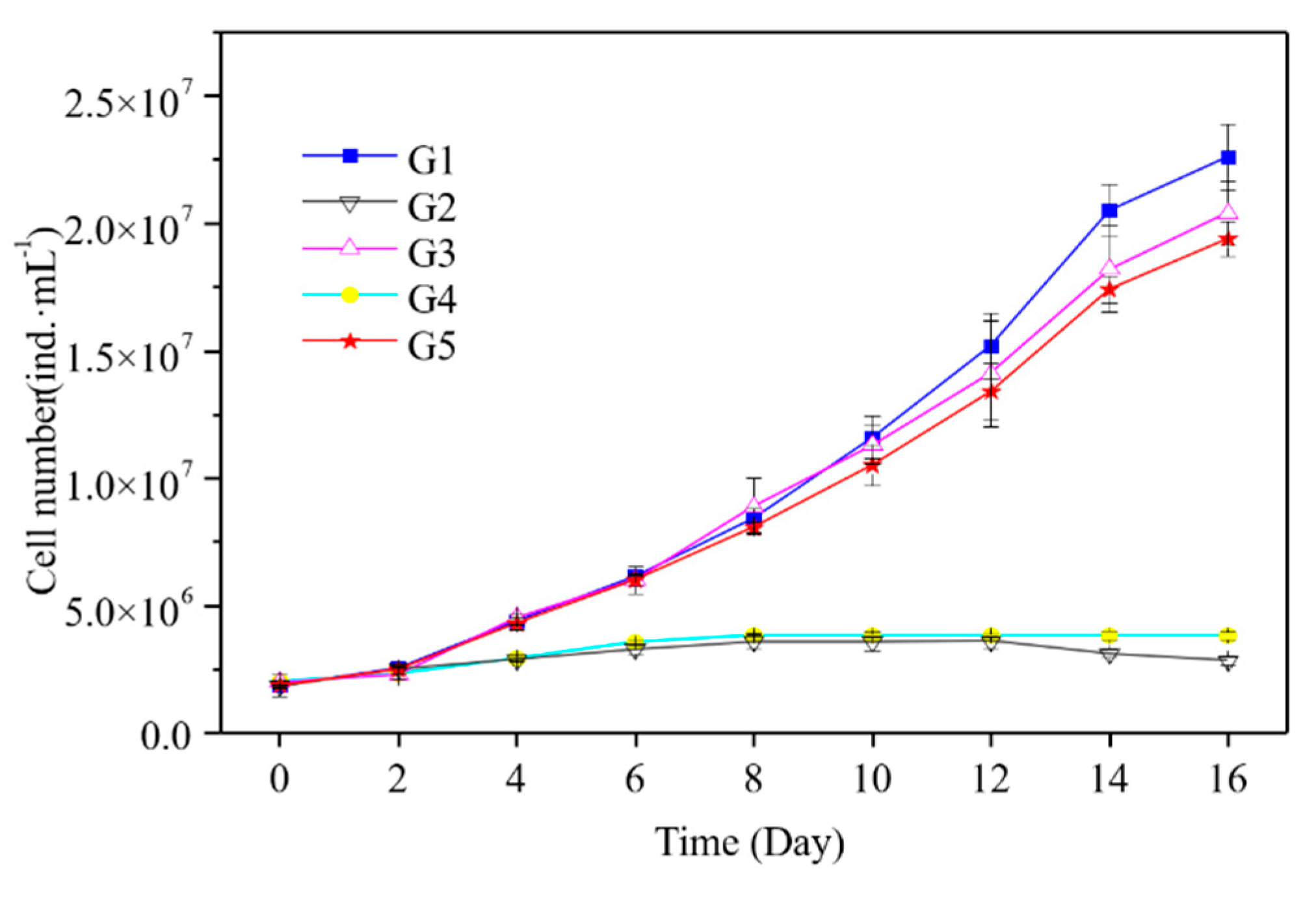
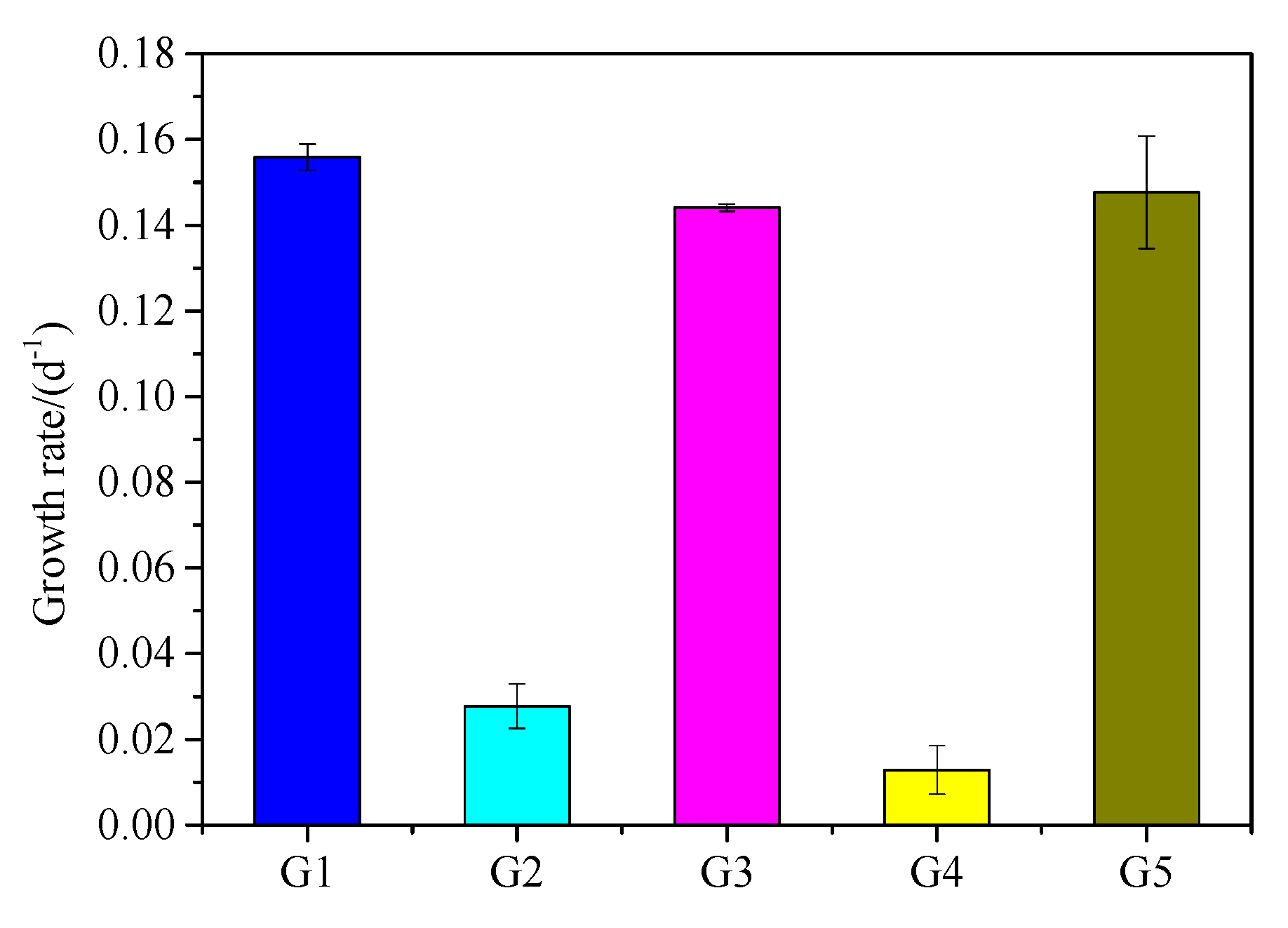
3.3. Cell density Changes of M. Aeruginosa in Culture Systems with Different P Sources
4. Discussion
4.1. Physicochemical Properties of PPOM
4.2. Degradation Characteristics of P in PPOM
4.2.1. Variation Characteristics of Organic P in PPOM
4.2.2. Variation Characteristics of Enzymatically Hydrolysable P in PPOM
4.3. Effect of PPOM on the Growth of M. Aeruginosa
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.W.; Qin, B.Q.; Li, Y.P.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hypereutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar] [PubMed]
- Qin, B.Q.; Gao, G.; Zhu, G.W.; Zhang, Y.L.; Song, Y.Z.; Tang, X.M.; Xu, H.; Deng, J.M. Lake eutrophication and its ecosystem response. Chin. Sci. Bull. 2013, 58, 961–970. [Google Scholar] [CrossRef]
- Álvarez-Rogel, J.; Jiménez-Cárceles, F.J.; Roca, M.J.; Ortiz, R. Changes in soils and vegetation in a Mediterranean coastal salt marsh impacted by human activities. Estuar. Coast. Shelf Sci. 2007, 73, 510–526. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jensen, J.P.; Jeppesen, E. Role of sediment and internal loading of P in shallow lakes. Hydrobiologia 2003, 506–509, 135–145. [Google Scholar] [CrossRef]
- Van der Grift, B.; Osté, L.; Schot, P.; Kratz, A.; van Popta, E.; Wassen, M.; Griffioen, J. Forms of P in suspended particulate matter in agriculture-dominated lowland catchments: Iron as P carrier. Sci. Total Environ. 2018, 631, 115–129. [Google Scholar] [CrossRef]
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef]
- Gu, B. Variations and controls of nitrogen stable isotopes in particulate organic matter of lakes. Oecologia 2009, 160, 421–431. [Google Scholar] [CrossRef]
- Shanks, A.L.; Edmondson, E.W. The vertical flux of metazoans (Holoplankton, Meiofauna, and Larval Invertebrates) due to their association with marine snow. Limnol. Oceanogr. 1990, 35, 455–463. [Google Scholar] [CrossRef]
- Goñi, M.A.; Voulgaris, G.; Kim, Y.H. Composition and fluxes of particulate organic matter in a temperate estuary (Winyah Bay, South Carolina, USA) under contrasting physical forcings. Estuar. Coast. Shelf Sci. 2009, 85, 273–291. [Google Scholar] [CrossRef]
- Middelburg, J.J.; Herman, P.M. Organic matter processing in tidal estuaries. Mar. Chem. 2007, 106, 127–147. [Google Scholar] [CrossRef]
- Morgan, B.; Rate, A.W.; Burton, E.D. Water chemistry and nutrient release during the resuspension of FeS-rich sediments in a eutrophic estuarine system. Sci. Total Environ. 2012, 432, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Bristow, L.A.; Jickells, T.D.; Weston, K.; Marca-Bell, A.; Parker, R.; Andrews, J.E. Tracing estuarine organic matter sources into the Southern North Sea using C and N isotopic signatures. Biogeochemistry 2013, 113, 9–22. [Google Scholar] [CrossRef]
- Kaiser, D.; Unger, D.; Qiu, G. Particulate organic matter dynamics in coastal systems of the Northern Beibu Gulf. Cont. Shelf Res. 2014, 82, 99–118. [Google Scholar] [CrossRef]
- Wetz, M.S.; Wheeler, P.A. Production and partitioning of organic matter during simulated phytoplankton blooms. Limnol. Oceanogr. 2003, 48, 1808–1817. [Google Scholar] [CrossRef] [Green Version]
- Countway, R.E.; Canuel, E.A.; Dickhut, R.M. Sources of particulate organic matter in surface waters of the York River, VA estuary. Org. Geochem. 2007, 38, 365–379. [Google Scholar] [CrossRef]
- Riemann, L.; Steward, G.F.; Azam, F. Dynamics of bacterial community composition and activity during a mesocosm diatom bloom. Appl. Environ. Microbiol. 2000, 66, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Chao, J.; Gong, Y.; Wang, Y.; Wilhelm, S.W.; Gao, G. Spatiotemporal dynamics of bacterial community composition in large shallow eutrophic Lake Taihu: High overlap between free-living and particle-attached assemblages. Limnol. Oceanogr. 2017, 62, 1366–1382. [Google Scholar] [CrossRef]
- Kong, M.; Chao, J.; Zhuang, W.; Wang, P.; Wang, C.; Hou, J.; Wu, Z.; Wang, L.; Gao, G.; Wang, Y. Spatial and temporal distribution of particulate P and their correlation with environmental factors in a shallow eutrophic Chinese lake (Lake Taihu). Int. J. Environ. Res. Public Health 2018, 15, 2355. [Google Scholar] [CrossRef]
- Jančula, D.; Maršálek, B. Critical review of actually available chemical compounds for prevention and management of cyanobacterial blooms. Chemosphere 2011, 85, 1415–1422. [Google Scholar] [CrossRef]
- Li, D.P.; Huang, Y. Synergistic effect of disturbance from wind-waves and benthic organisms on release of internal P from sediments. China Water Wastewater 2013, 29, 17–20. (In Chinese) [Google Scholar]
- Frka, S.; Gašparović, B.; Marić, D.; Godrijan, J.; Djakovac, T.; Vojvodić, V.; Dautović, J.; Kozarac, K. Phytoplankton driven distribution of dissolved and particulate lipids in a semi-enclosed temperate sea (mediterranean): Spring to summer situation. Estuar. Coast. Shelf Sci. 2011, 93, 290–304. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Kanemata, K.; Nakase, G.; Eguchi, M. Microbial decomposition process of organic matter in sinking particles, resuspendable particles, and bottom sediments at a coastal fish farming area. Fish. Sci. 2017, 83, 1–13. [Google Scholar]
- Dokulil, M.T.; Teubner, K. Cyanobacterial dominance in lakes. Hydrobiologia 2000, 438, 1–12. [Google Scholar] [CrossRef]
- Jiang, X.; Jin, X.; Yao, Y.; Li, L.; Wu, F. Effects of biological activity, light, temperature and oxygen on P release processes at the sediment and water interface of Taihu Lake, China. Water Res. 2008, 42, 2251–2259. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Paerl, H.W.; Qin, B.; Zhu, G.; Hall, N.S.; Wu, Y. Determining critical nutrient thresholds needed to control harmful cyanobacterial blooms in eutrophic Lake Taihu, China. Environ. Sci. Technol. 2014, 49, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Paerl, H.W.; Brookes, J.D.; Liu, J.; Jeppesen, E.; Zhu, G.; Zhang, Y.; Xu, H.; Shi, K.; Deng, J.; et al. Why Lake Taihu continues to be plagued with cyanobacterial blooms through 10 years (2007–2017) efforts. Sci. Bull. 2019, 64, 354–356. [Google Scholar] [CrossRef]
- Chen, Y.; Qin, B.; Teubner, K.; Dokulil, M.T. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China. J. Plankton Res. 2003, 25, 445–453. [Google Scholar] [CrossRef]
- Jensen, H.S.; Kristensen, P.; Jeppesen, E.; Skytthe, A. Iron: P ratio in surface sediment as an indicator of phosphate release from aerobic sediments in shallow lakes. Hydrobiologia 1992, 235, 731–743. [Google Scholar] [CrossRef]
- Hatten, J.A.; Goñi, M.A.; Wheatcroft, R.A. Chemical characteristics of particulate organic matter from a small, mountainous river system in the Oregon Coast Range, USA. Biogeochemistry 2012, 107, 43–66. [Google Scholar] [CrossRef]
- Solórzano, L.; Sharp, J.H. Determination of total dissolved P and particulate P in natural waters1. Limnol. Oceanogr. 1980, 25, 754–758. [Google Scholar] [CrossRef]
- Chen, Y.W.; Gao, X.Y. Comparison of two methods for phytoplankton chlorophyll a concentration measurement. J. Lake Sci. 2000, 12, 185–188. (In Chinese) [Google Scholar]
- Read, E.K.; Ivancic, M.; Hanson, P.; Cade-Menun, B.J.; McMahon, K.D. P speciation in a eutrophic lake by 31P NMR spectroscopy. Water Res. 2014, 62, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Sun, J.; Gu, L.; Zhao, H.; Wang, J. Variations of different dissolved and particulate P classes during an algae bloom in a eutrophic lake by 31P NMR spectroscopy. Chemosphere 2017, 169, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Ding, S.; Fan, C.; Liu, T.; Shi, D.; Zhang, L. Organic P species in surface sediments of a large, shallow, eutrophic lake, Lake Taihu, China. Environ. Pollut. 2009, 157, 2507–2513. [Google Scholar] [CrossRef]
- Ding, S.; Xu, D.; Bai, X.; Yao, S.; Fan, C.; Zhang, C. Speciation of organic P in a sediment profile of Lake Taihu II. Molecular species and their depth attenuation. J. Environ. Sci. 2013, 25, 925–932. [Google Scholar] [CrossRef]
- Gao, G.; Zhu, G.; Qin, B.; Chen, J.; Wang, K. Alkaline phosphatase activity and the phosphorus mineralization rate of Lake Taihu. Sci. China Ser. D 2006, 49, 176–185. (In Chinese) [Google Scholar] [CrossRef]
- Chrost, R.J.; Siuda, W.; Albrecht, D.; Overbeck, J. A method for determining enzymatically hydrolyzable phosphate (EHP) in natural waters. Limnol. Oceanogr. 1986, 31, 662–667. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, Z.; Wang, D.; Ma, M.; Wang, C. Distribution and origin of biologically available P in the water of the Meiliang Bay in summer. Sci. China Ser. D 2006, 49, 146–153. (In Chinese) [Google Scholar] [CrossRef]
- Chuai, X.; Ding, W.; Chen, X.; Wang, X.; Miao, A.; Xi, B.; He, L.; Yang, L. P release from cyanobacterial blooms in Meiliang Bay of Lake Taihu, China. Ecol. Eng. 2011, 37, 842–849. [Google Scholar] [CrossRef]
- Chao, J.Y.; Gao, G.; Tang, X.M.; Shen, Y.; Zhu, L.P. Effects of ecosystem structure on the organic aggregates physical and chemical parameters of Lake Taihu. Huan Jing Ke Xue 2009, 30, 3196–3202. (In Chinese) [Google Scholar]
- Bi, L.L.; Bai, J.; Zhao, Z.S.; Li, Y.G.; Yun, Z. Characteristics of suspended particles in summer in Jiaozhou Bay. Mar. Environ. Sci. 2007, 26, 518–522. [Google Scholar]
- Cifuentes, L.A.; Sharp, J.H.; Fogel, M.L. Stable carbon and nitrogen isotope biogeochemistry in the Delaware estuary. Limnol. Oceanogr. 1988, 33, 1102–1115. [Google Scholar] [CrossRef]
- Shi, L.; Huang, Y.; Lu, Y.; Chen, F.; Zhang, M.; Yu, Y.; Kong, F. Stocks and dynamics of particulate and dissolved organic matter in a large, shallow eutrophic lake (Taihu, China) with dense cyanobacterial blooms. J. Oceanol. Limnol. 2018, 36, 738–749. [Google Scholar] [CrossRef]
- Ahlgren, J.; Reitzel, K.; Tranvik, L.; Gogoll, A.; Rydin, E. Degradation of organic P compounds in anoxic Baltic Sea sediments: A 31P nuclear magnetic resonance study. Limnol. Oceanogr. 2006, 51, 2341–2348. [Google Scholar] [CrossRef]
- Makarov, M.; Haumaier, L.; Zech, W. The nature and origins of diester phosphates in soils: A 31 P-NMR study. Biol. Fertil. Soils 2002, 35, 136–146. [Google Scholar]
- Hakulinen, R.; Kähkönen, M.A.; Salkinoja-Salonen, M. Vertical distribution of sediment enzyme activities involved in the cycling of carbon, nitrogen, P and sulphur in three boreal rural lakes. Water Res. 2005, 39, 2319–2326. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, F.; He, Z.; Guo, J.; Qu, X.; Xie, F.; Giesy, J.P.; Liao, H.; Guo, F. Characterization of organic P in lake sediments by sequential fractionation and enzymatic hydrolysis. Environ. Sci. Technol. 2013, 47, 7679–7687. [Google Scholar] [CrossRef]
- Magen, C.; Chaillou, G.; Crowe, S.A.; Mucci, A.; Sundby, B.; Gao, A.; Makabe, R.; Sasaki, H. Origin and fate of particulate organic matter in the southern Beaufort Sea–Amundsen Gulf region, Canadian Arctic. Estuar. Coast. Shelf Sci. 2010, 86, 31–41. [Google Scholar] [CrossRef]
- Smith, D.C.; Simon, M.; Alldredge, A.L.; Azam, F. Intense hydrolytic enzyme activity on marine aggregates and implications for rapid particle dissolution. Nature 1992, 359, 139–142. [Google Scholar] [CrossRef]
- Peacock, M.B.; Kudela, R.M. Alkaline phosphatase activity detected in distinct phytoplankton communities in the northern Gulf of Alaska. Mar. Ecol. Prog. Ser. 2013, 473, 79–90. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date (d) | T (°C) | Turbidity (NTU) | DO (mg·L−1) | pH | SS mg·L−1 | POM mg·L−1 | POC mg·L−1 | POP mg·L−1 | TPP mg·L−1 | TP mg·L−1 | Chl a μg·L−1 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 29.9 ± 0.1 | 291.65 ± 10.33 | 7.12 ± 0.50 | 7.86 ± 0.06 | 135.32 ± 12.38 | 130.12 ± 2.17 | 56.43 ± 2.18 | 0.97 ± 0.35 | 0.99 ± 0.34 | 1.12 ± 0.26 | 837.45 ± 112.38 |
| 2 | 30.1 ± 0.1 | 156.62 ± 10.68 | 3.42 ± 0.61 | 7.60 ± 0.09 | 117.65 ± 11.31 | 127.54 ± 13.43 | 74.05 ± 5.33 | 0.56 ± 0.11 | 0.59 ± 0.10 | 0.65 ± 0.17 | 740.22 ± 94.03 |
| 6 | 30.2 ± 0.1 | 108.81 ± 8.32 | 5.75 ± 0.62 | 7.58 ± 0.08 | 100.89 ± 18.08 | 89.33 ± 7.79 | 57.26 ± 3.49 | 0.53 ± 0.21 | 0.54 ± 0.21 | 0.59 ± 0.16 | 461.53 ± 58.57 |
| 11 | 30.3 ± 0.1 | 197.42 ± 18.87 | 5.94 ± 0.34 | 7.51 ± 0.04 | 115.52 ± 12.12 | 114.46 ± 15.78 | 60.38 ± 3.98 | 0.57 ± 0.18 | 0.58 ± 0.17 | 0.63 ± 0.22 | 470.65 ± 43.31 |
| 14 | 30.2 ± 0.1 | 230.07 ± 25.34 | 6.15 ± 0.27 | 7.72 ± 0.12 | 134.26 ± 24.56 | 123.35 ± 20.02 | 61.47 ± 5.45 | 0.52 ± 0.12 | 0.53 ± 0.12 | 0.66 ± 0.15 | 412.28 ± 83.02 |
| 20 | 30.5 ± 0.1 | 332.95 ± 20.27 | 6.48 ± 0.18 | 7.85 ± 0.10 | 142.75 ± 19.53 | 118.85 ± 20.66 | 57.57 ± 9.25 | 0.49 ± 0.28 | 0.50 ± 0.28 | 0.62 ± 0.15 | 417.45 ± 64.39 |
| Species | 0d % | 1d % | 2d % | 4d % | 6d % | 8d % | 11d % | 14d % | 20d % |
|---|---|---|---|---|---|---|---|---|---|
| Ortho-P | 46.68 | 25.63 | 29.00 | 33.40 | 46.04 | 48.24 | 42.83 | 25.71 | 32.14 |
| Mono-P | 38.75 | 51.96 | 48.35 | 46.48 | 45.2 | 45.90 | 34.43 | 37.62 | 37.26 |
| Diester-P | 6.66 | 8.29 | 7.90 | 10.28 | 7.09 | 0.00 | 0.81 | 5.27 | 2.38 |
| Pyro-P | 6.53 | 12.63 | 12.92 | 9.85 | 1.65 | 5.86 | 20.32 | 27.55 | 23.43 |
| Poly-P | 1.37 | 1.48 | 1.84 | 0.00 | 0.00 | 0.00 | 1.61 | 3.85 | 4.79 |
| Pi | 54.58 | 39.74 | 43.76 | 43.25 | 47.69 | 54.10 | 64.76 | 57.11 | 60.36 |
| Po | 45.41 | 60.25 | 56.25 | 56.76 | 52.31 | 45.90 | 35.24 | 42.89 | 39.64 |
| Bio-P | 53.31 | 74.36 | 71.01 | 66.61 | 53.96 | 51.76 | 57.17 | 74.29 | 67.86 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, M.; Chao, J.; Han, W.; Ye, C.; Li, C.-H.; Tian, W. Degradation Characteristics of Phosphorus in Phytoplankton-Derived Particulate Organic Matter and Its Effects on the Growth of Phosphorus-Deficient Microcystis aeruginosa in Lake Taihu. Int. J. Environ. Res. Public Health 2019, 16, 2155. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16122155
Kong M, Chao J, Han W, Ye C, Li C-H, Tian W. Degradation Characteristics of Phosphorus in Phytoplankton-Derived Particulate Organic Matter and Its Effects on the Growth of Phosphorus-Deficient Microcystis aeruginosa in Lake Taihu. International Journal of Environmental Research and Public Health. 2019; 16(12):2155. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16122155
Chicago/Turabian StyleKong, Ming, Jianying Chao, Wei Han, Chun Ye, Chun-Hua Li, and Wei Tian. 2019. "Degradation Characteristics of Phosphorus in Phytoplankton-Derived Particulate Organic Matter and Its Effects on the Growth of Phosphorus-Deficient Microcystis aeruginosa in Lake Taihu" International Journal of Environmental Research and Public Health 16, no. 12: 2155. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16122155