The Antialgal Mechanism of Luteolin-7-O-Glucuronide on Phaeocystis globosa by Metabolomics Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Cultures
2.2. Antialgal Assays
2.3. LC-MS/MS-Based Untargeted Metabolomic Analysis
2.3.1. Metabolites Extraction
2.3.2. LC-MS/MS Condition
2.3.3. Data Analysis
3. Results
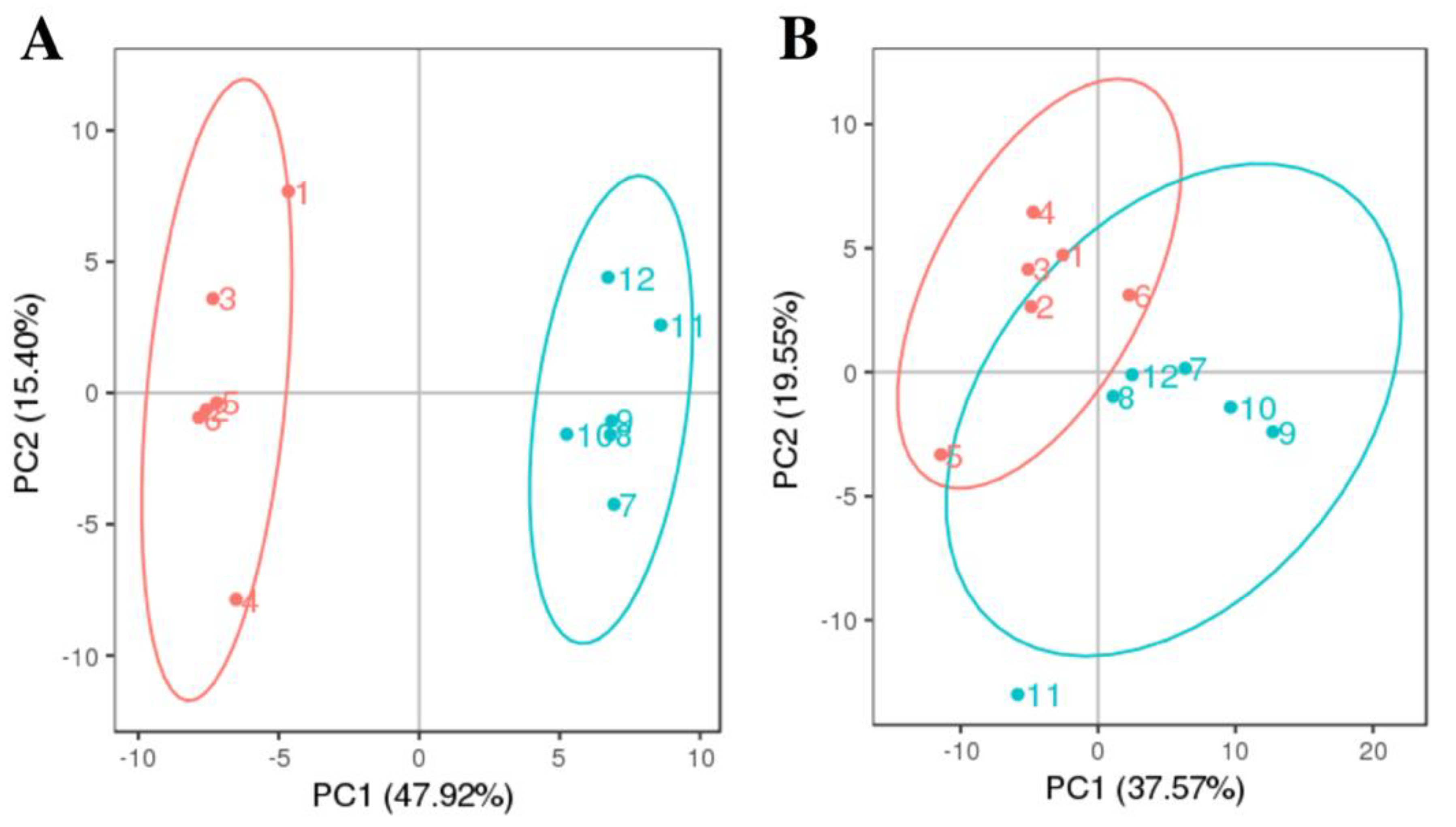
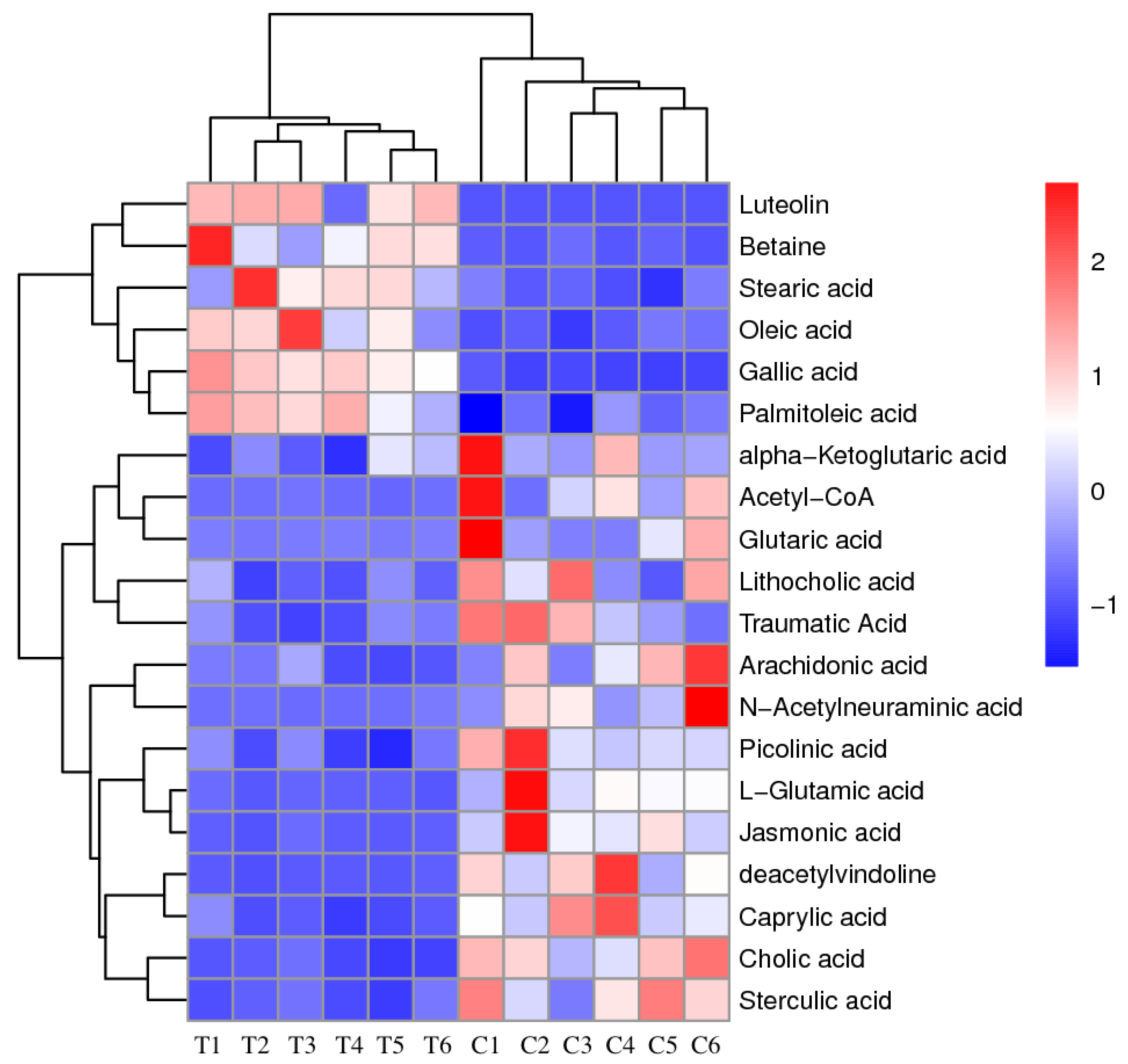
3.1. PCA Analysis and Clustering Analysis of LC-MS/MS Metabolomics Profiles of P. Globosa
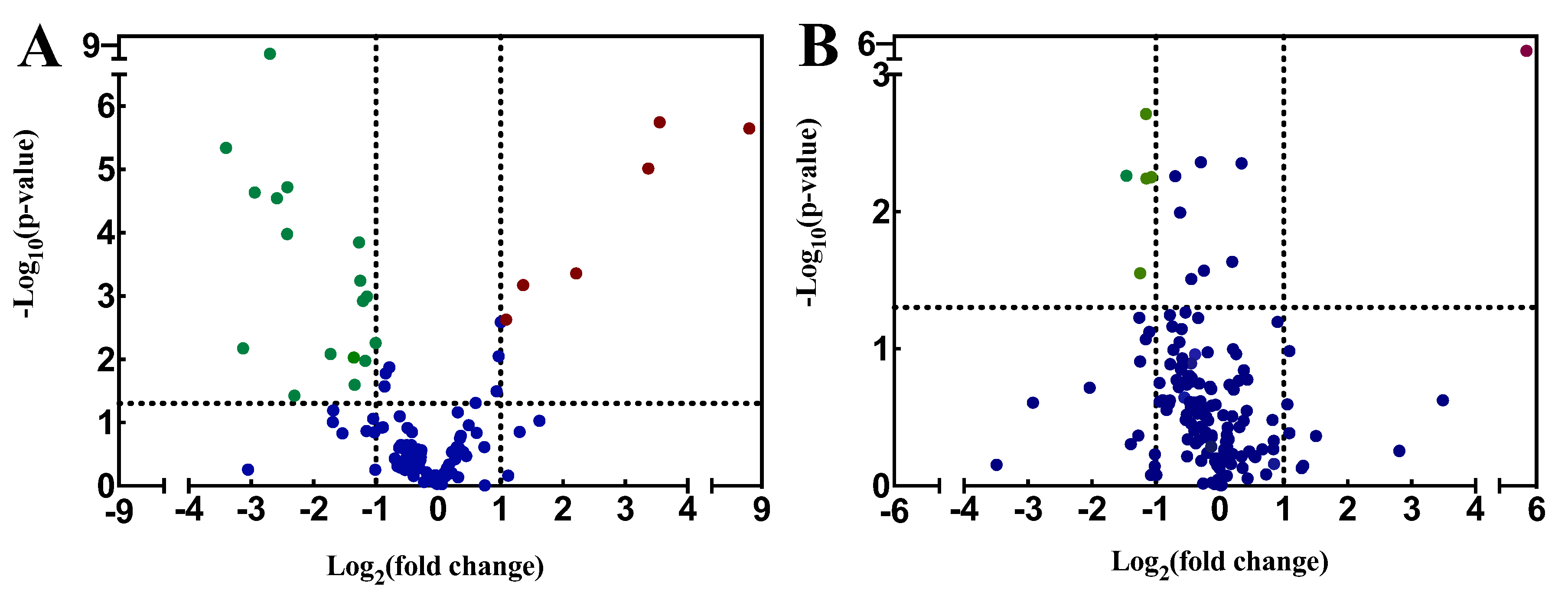
3.2. Volcano Plot Analysis of LC-MS/MS Metabolic Profiles of P. globosa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- De Cruz, A.; Logsdon, R.; Lye, D.; Guglielmi, S.; Rice, A.; Kannan, M.S. Harmful Algae Bloom Occurrence in Urban Ponds: Relationship of Toxin Levels with Cell Density and Species Composition. J. Earth Environ. Sci. 2017, 25, 704–726. [Google Scholar]
- Chai, Y.; Deng, W.J.; Qin, X.; Xu, X. Occurrence of four species of algae in the marine water of Hong Kong. Mar. Pollut. Bull. 2017, 124, 890–896. [Google Scholar] [CrossRef]
- Johnson, M.-V.V.; Scott, G.I.; Perkins, D.A.K.; Smith, S.A.; Reavie, E.D.; Brooks, B.W.; Lazorchak, J.M.; Morton, S.L.; Howard, M.D.A.; Steevens, J.A. Are harmful algal blooms becoming the greatest inland water quality threat to public health and aquatic ecosystems? Environ. Toxicol. Chem. 2016, 35, 6–13. [Google Scholar]
- Zohdi, E.; Abbaspour, M. Harmful algal blooms (red tide): A review of causes, impacts and approaches to monitoring and prediction. Int. J. Environ. Sci. Technol. 2019, 16, 1789–1806. [Google Scholar] [CrossRef]
- Schoemann, V.; Becquevort, S.; Stefels, J.; Rousseau, V.; Lancelot, C. Phaeocystis blooms in the global ocean and their controlling mechanisms: A review. J. Sea Res. 2005, 53, 43–66. [Google Scholar] [CrossRef]
- Li, L.; Lü, S.; Cen, J. Spatio-temporal variations of Harmful algal blooms along the coast of Guangdong, Southern China during 1980–2016. J. Oceanol. Limnol. 2019, 37, 535–551. [Google Scholar] [CrossRef]
- Tan, S.; Hu, X.; Yin, P.; Zhao, L. Photosynthetic inhibition and oxidative stress to the toxic Phaeocystis globosa caused by a diketopiperazine isolated from products of algicidal bacterium metabolism. J. Microbiol. 2016, 54, 364–375. [Google Scholar] [CrossRef]
- Zhuang, L.; Zhao, L.; Yin, P. Combined algicidal effect of urocanic acid, N-acetylhistamine and l-histidine to harmful alga Phaeocystis globosa. RSC Adv. 2018, 8, 12760–12766. [Google Scholar] [CrossRef]
- Park, J.; Church, J.; Son, Y.; Kim, K.T.; Lee, W.H. Recent advances in ultrasonic treatment: Challenges and field applications for controlling harmful algal blooms (HABs). Ultrason. Sonochem. 2017, 38, 326–334. [Google Scholar] [CrossRef]
- Kim, Z.-H.; Thanh, N.N.; Yang, J.-H.; Park, H.; Yoon, M.-Y.; Park, J.-K.; Lee, C.-G. Improving microalgae removal efficiency using chemically-processed clays. Biotechnol. Bioprocess Eng. 2016, 21, 787–793. [Google Scholar] [CrossRef]
- Song, L.Y.; Wang, Y.Q. Investigation of microbial community structure of a shallow lake after one season copper sulfate algaecide treatment. Microbiol. Res. 2015, 170, 105–113. [Google Scholar] [CrossRef]
- McComas, S. Lake and Pond Management Guidebook; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Sun, Y.; Zhou, W.; Wang, H.; Guo, G.; Su, Z.; Pu, Y. Antialgal compounds with antialgal activity against the common red tide microalgae from a green algae Ulva pertusa. Ecotoxicol. Environ. Safe 2018, 157, 61–66. [Google Scholar] [CrossRef]
- Chen, S.; Zheng, T.; Ye, C.; Huannixi, W.; Yakefu, Z.; Meng, Y.; Peng, X.; Tian, Z.; Wang, J.; Ma, Y.; et al. Algicidal properties of extracts from Cinnamomum camphora fresh leaves and their main compounds. Ecotoxicol. Environ. Safe 2018, 163, 594–603. [Google Scholar] [CrossRef]
- Patiño, R.; Rashel, R.H.; Rubio, A.; Longing, S. Growth-suppressing and algicidal properties of an extract from Arundo donax, an invasive riparian plant, against Prymnesium parvum, an invasive harmful alga. Harmful Algae 2018, 71, 1–9. [Google Scholar] [CrossRef]
- Shi, P.; Geng, S.; Feng, T.; Wu, H. Effects of Ascophyllum nodosum extract on growth and antioxidant defense systems of two freshwater microalgae. J. Appl. Phycol. 2018, 30, 851–859. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.; Zhang, P.; Zeng, G.; Cai, X.; Liu, S.; Yin, Y.; Hu, X.; Hu, X.; Tan, X. Growth inhibition and oxidative damage of Microcystis aeruginosa induced by crude extract of Sagittaria trifolia tubers. J. Environ. Sci. (China) 2016, 43, 40–47. [Google Scholar] [CrossRef]
- Mulderij, G.; Mooij, W.M.; Donk, E.V. Allelopathic growth inhibition and colony formation of the green alga Scenedesmus obliquus by the aquatic macrophyte Stratiotes aloides. Aquat. Ecol. 2005, 39, 11–21. [Google Scholar] [CrossRef]
- Pei, Y.; Liu, L.; Hilt, S.; Xu, R.; Wang, B.; Li, C.; Chang, X. Root exudated algicide of Eichhornia crassipes enhances allelopathic effects of cyanobacteria Microcystis aeruginosa on green algae. Hydrobiologia 2018, 823, 67–77. [Google Scholar] [CrossRef]
- Qian, H.; Xu, X.; Chen, W.; Jiang, H.; Jin, Y.; Liu, W.; Fu, Z. Allelochemical stress causes oxidative damage and inhibition of photosynthesis in Chlorella vulgaris. Chemosphere 2009, 75, 368–375. [Google Scholar] [CrossRef]
- Wu, X.; Wu, H.; Wang, S.; Wang, Y.; Zhang, R.; Hu, X.; Ye, J. Effect of propionamide on the growth of Microcystis flos-aquae colonies and the underlying physiological mechanisms. Sci. Total Environ. 2018, 630, 526–535. [Google Scholar] [CrossRef]
- Sanaei, Z.; Shahrabi, T.; Ramezanzadeh, B. Synthesis and characterization of an effective green corrosion inhibitive hybrid pigment based on zinc acetate-Cichorium intybus L leaves extract (ZnA-CIL.L): Electrochemical investigations on the synergistic corrosion inhibition of mild steel in aqueous. Dyes Pigments 2017, 139, 218–232. [Google Scholar] [CrossRef]
- Sareedenchai, V.; Zidorn, C. Flavonoids as chemosystematic markers in the tribe Cichorieae of the Asteraceae. Biochem. Syst. Ecol. 2010, 38, 935–957. [Google Scholar] [CrossRef]
- Lu, Y.; Yeap Foo, L. Flavonoid and phenolic glycosides from Salvia officinalis. Phytochemistry 2000, 55, 263–267. [Google Scholar] [CrossRef]
- Leyva-López, N.; Nair, V.; Bang, W.Y.; Cisneros-Zevallos, L.; Heredia, J.B. Protective role of terpenes and polyphenols from three species of Oregano (Lippia graveolens, Lippia palmeri and Hedeoma patens) on the suppression of lipopolysaccharide-induced inflammation in RAW 264.7 macrophage cells. J. Ethnopharmacol. 2016, 187, 302–312. [Google Scholar] [CrossRef]
- Zhu, J.; Xiao, H.; Chen, Q.; Zhao, M.; Sun, D.; Duan, S. Growth Inhibition of Phaeocystis Globosa Induced by Luteolin-7-O-glucuronide from Seagrass Enhalus acoroides. Int. J. Environ. Res. Public Health 2019, 16, 2615. [Google Scholar] [CrossRef]
- Petrović, S.; Ušjak, L.; Milenković, M.; Arsenijević, J.; Drobac, M.; Drndarević, A.; Niketić, M. Thymus dacicus as a new source of antioxidant and antimicrobial metabolites. J. Funct. Foods 2017, 28, 114–121. [Google Scholar] [CrossRef]
- Orhan, F.; Çeker, S.; Anar, M.; Agar, G.; Arasoglu, T. Protective effects of three luteolin derivatives on aflatoxin B 1-induced genotoxicity on human blood cells. Med. Chem. Res. 2016, 25, 2567–2577. [Google Scholar] [CrossRef]
- Jeyadevi, R.; Sivasudha, T.; Rameshkumar, A.; Dinesh Kumar, L. Anti-arthritic activity of the Indian leafy vegetable Cardiospermum halicacabum in Wistar rats and UPLC--QTOF--MS/MS identification of the putative active phenolic components. Inflamm. Res. 2013, 62, 115–126. [Google Scholar] [CrossRef]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought Responses of Leaf Tissues from Wheat Cultivars of Differing Drought Tolerance at the Metabolite Level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Diao, J.; Li, X.; Pei, G.; Liu, L.; Chen, L. Metabolomic analysis of Crypthecodinium cohnii in response to different dissolved oxygen levels during docosahexaenoic acid fermentation. Biochem. Biophys. Res. Commun. 2018, 499, 941–947. [Google Scholar] [CrossRef]
- Granucci, N.; Pinu, F.R.; Han, T.L.; Villas-Boas, S.G. Can we predict the intracellular metabolic state of a cell based on extracellular metabolite data? Mol. Biosyst. 2015, 11, 3297–3304. [Google Scholar] [CrossRef]
- Barrett, T.; Suresh, C.G.; Tolley, S.P.; Dodson, E.J.; Hughes, M.A. The crystal structure of a cyanogenic β-glucosidase from white clover, a family 1 glycosyl hydrolase. Structure 1995, 3, 951–960. [Google Scholar] [CrossRef]
- Xiao, X.; Li, C.; Huang, H.; Lee, Y.P. Inhibition effect of natural flavonoids on red tide alga Phaeocystis globosa and its quantitative structure-activity relationship. Environ. Sci. Pollut. Res. 2019, 26, 1–14. [Google Scholar] [CrossRef]
- Sui, N.; Wang, Y.; Liu, S.; Yang, Z.; Wang, F.; Wan, S. Transcriptomic and Physiological Evidence for the Relationship between Unsaturated Fatty Acid and Salt Stress in Peanut. Front. Plant Sci. 2018, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Graham, D.; Patterson, B.D. Responses of Plants to Low, Nonfreezing Temperatures: Proteins, Metabolism, and Acclimation. Annu. Rev. Plant Physiol. 1982, 33, 347–372. [Google Scholar] [CrossRef]
- Zhang, W.; Tan, N.G.J.; Li, S.F.Y. NMR-based metabolomics and LC-MS/MS quantification reveal metal-specific tolerance and redox homeostasis in Chlorella vulgaris. Mol. Biosyst. 2014, 10, 149–160. [Google Scholar] [CrossRef]
- Aurich, A.; Specht, R.; Müller, R.A.; Stottmeister, U.; Yovkova, V.; Otto, C.; Holz, M.; Barth, G.; Heretsch, P.; Thomas, F.A.; et al. Microbiologically Produced Carboxylic Acids Used as Building Blocks in Organic Synthesis. In Reprogramming Microbial Metabolic Pathways; Wang, X., Chen, J., Quinn, P., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 391–423. ISBN 978-94-007-5055-5. [Google Scholar]
- Wiskich, J.T.; Dry, I.B. The Tricarboxylic Acid Cycle in Plant Mitochondria: Its Operation and Regulation BT—Higher Plant Cell Respiration; Douce, R., Day, D.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 281–313. ISBN 978-3-642-70101-6. [Google Scholar]
- Bromke, M.A. Amino acid biosynthesis pathways in diatoms. Metabolites 2013, 3, 294–311. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Metabolites | Log2 (Fold Change) | −Log10 (p Value) | VIP | Up/Down |
|---|---|---|---|---|
| Acetyl-CoA | −3.13 | 2.17 | 1.95 | down |
| L-Glutamic acid | −2.94 | 4.64 | 1.62 | down |
| alpha-Ketoglutaric acid | −3.40 | 5.34 | 1.91 | down |
| Arachidonic acid | −1.36 | 2.03 | 0.73 | down |
| Deacetylvindoline | −2.42 | 3.98 | 1.33 | down |
| Glutaric acid | −2.31 | 1.43 | 1.25 | down |
| Jasmonic acid | −2.59 | 4.55 | 1.43 | down |
| Lithocholic acid | −1.34 | 1.60 | 0.72 | down |
| Caprylic acid | −1.27 | 3.85 | 0.71 | down |
| Cholic acid | −2.42 | 4.72 | 1.81 | down |
| N-Acetylneuraminic acid | −1.73 | 2.08 | 1.10 | down |
| Picolinic acid | −1.14 | 2.99 | 0.66 | down |
| Sterculic acid | −1.21 | 2.92 | 0.85 | down |
| Traumatic acid | −1.17 | 1.97 | 0.63 | down |
| Luteolin | 8.51 | 5.65 | 4.78 | up |
| Gallic acid | 3.55 | 5.74 | 2.08 | up |
| Oleic acid | 2.21 | 3.36 | 1.64 | up |
| Betaine | 3.37 | 5.01 | 1.91 | up |
| Palmitoleic acid | 1.09 | 2.63 | 0.66 | up |
| Stearic acid | 1.36 | 3.17 | 0.97 | up |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Yang, Y.; Duan, S.; Sun, D. The Antialgal Mechanism of Luteolin-7-O-Glucuronide on Phaeocystis globosa by Metabolomics Analysis. Int. J. Environ. Res. Public Health 2019, 16, 3222. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16173222
Zhu J, Yang Y, Duan S, Sun D. The Antialgal Mechanism of Luteolin-7-O-Glucuronide on Phaeocystis globosa by Metabolomics Analysis. International Journal of Environmental Research and Public Health. 2019; 16(17):3222. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16173222
Chicago/Turabian StyleZhu, Jingyi, Yeyin Yang, Shunshan Duan, and Dong Sun. 2019. "The Antialgal Mechanism of Luteolin-7-O-Glucuronide on Phaeocystis globosa by Metabolomics Analysis" International Journal of Environmental Research and Public Health 16, no. 17: 3222. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16173222