Effect of Specific Retinoic Acid Receptor Agonists on Noise-Induced Hearing Loss


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Noise Generation
2.3. All-Trans Retinoic Acid (ATRA) and Selective RAR Agonist Treatment
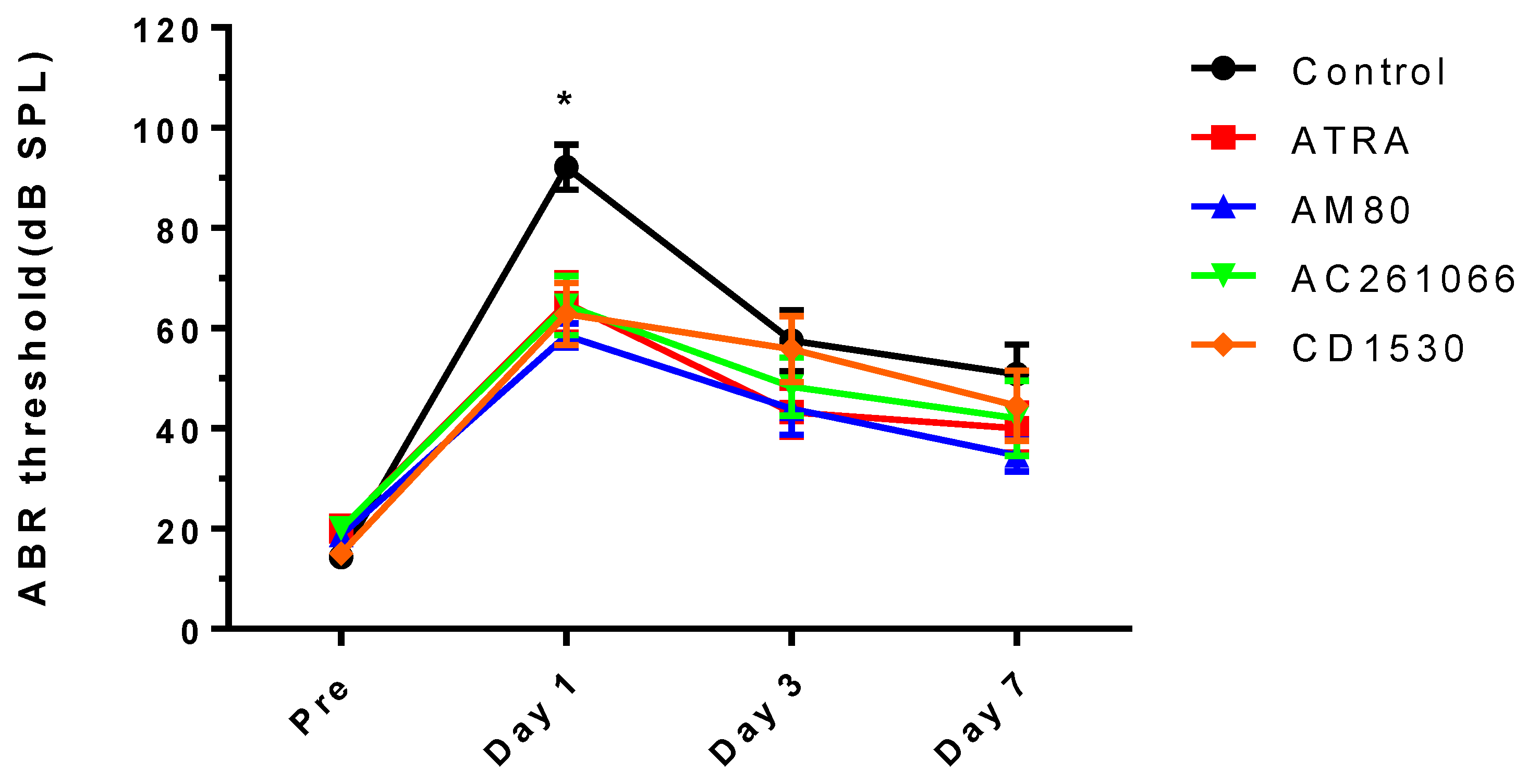
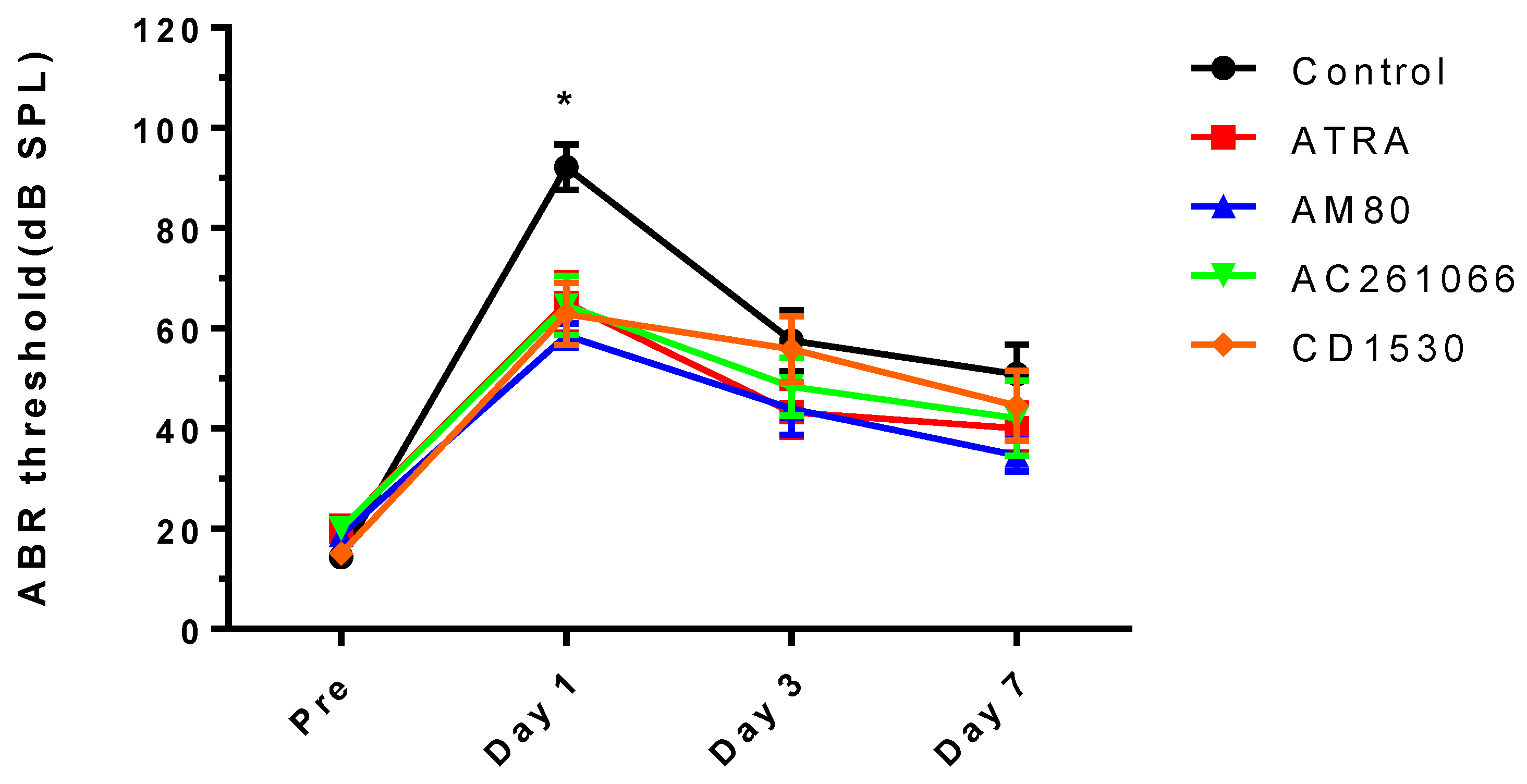
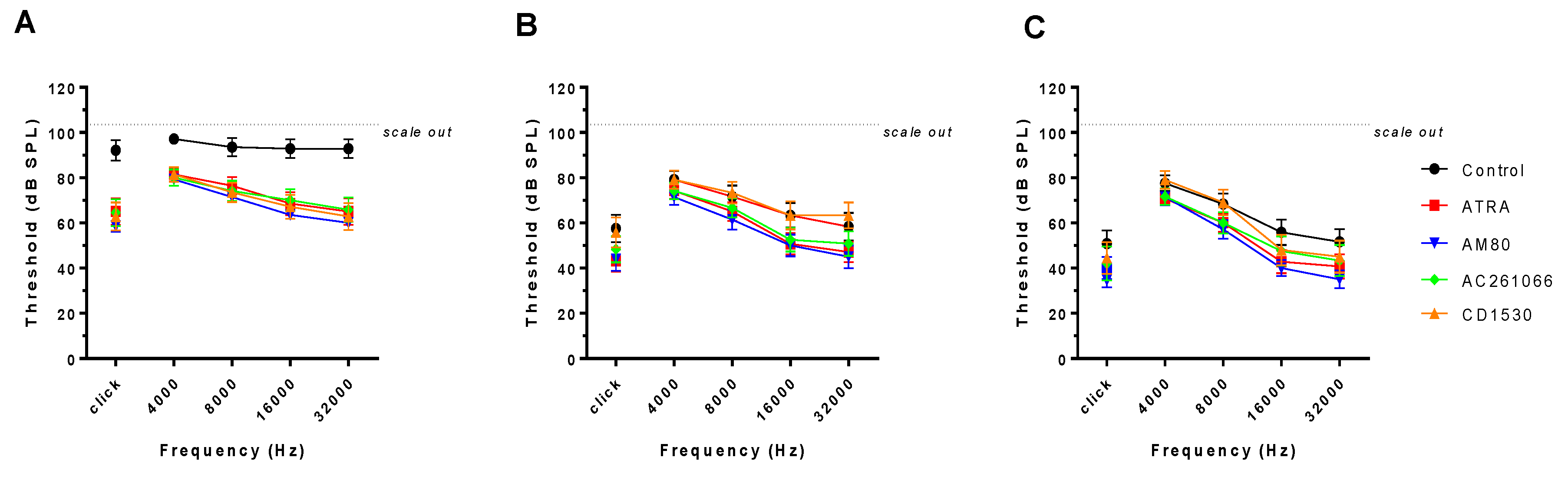
2.4. Audiologic Evaluation
2.5. Cochlear Cell Survival
2.6. Statistics
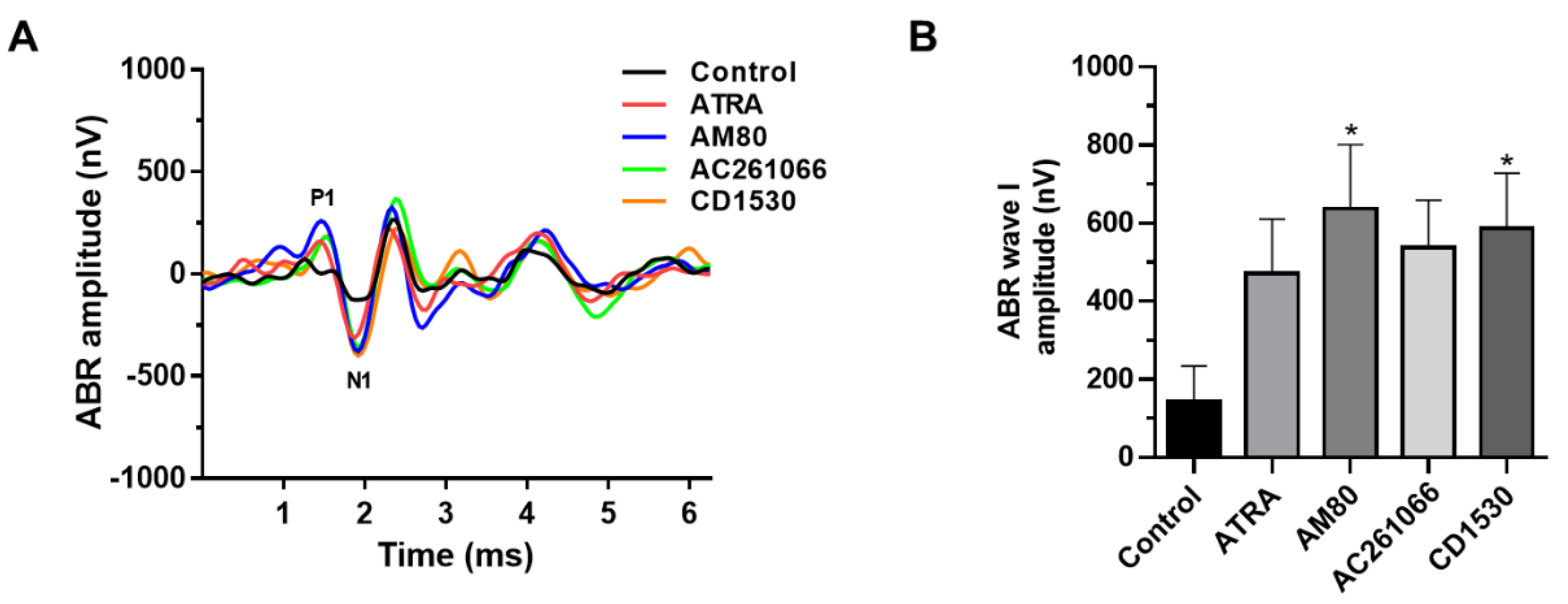
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| NIHL | Noise induced hearing loss |
| RA | Retinoic acid |
| RAR | Retinoic acid receptor |
| ATRA | All-trans retinoic acid |
| IP | Intraperitoneal |
References
- Moszynski, P. WHO warns noise pollution is a growing hazard to health in Europe. BMJ 2011, 342, d2114. [Google Scholar] [CrossRef] [PubMed]
- Henderson, D.; Bielefeld, E.C.; Harris, K.C.; Hu, B.H. The role of oxidative stress in noise-induced hearing loss. Ear Hear. 2006, 27, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Kang, H.H.; Kim, Y.J.; Chung, J.W. Anti-apoptotic role of retinoic acid in the inner ear of noise-exposed mice. Biochem. Biophys. Res. Commun. 2005, 335, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Shim, H.J.; Kang, H.H.; Ahn, J.H.; Chung, J.W. Retinoic acid applied after noise exposure can recover the noise-induced hearing loss in mice. Acta Otolaryngol. 2009, 129, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Shin, J.E.; Chung, B.Y.; Lee, H.M.; Kang, H.H.; Chung, J.W.; Pak, J.H. Involvement of retinoic acid-induced peroxiredoxin 6 expression in recovery of noise-induced temporary hearing threshold shifts. Environmental Toxicol. Pharmacol. 2013, 36, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Romand, R.; Dolle, P.; Hashino, E. Retinoid signaling in inner ear development. J. Neurobiol. 2006, 66, 687–704. [Google Scholar] [CrossRef] [PubMed]
- Romand, R.; Sapin, V.; Dolle, P. Spatial distributions of retinoic acid receptor gene transcripts in the prenatal mouse inner ear. J. Comp. Neurol. 1998, 393, 298–308. [Google Scholar] [CrossRef]
- Romand, R.; Hashino, E.; Dolle, P.; Vonesch, J.L.; Chambon, P.; Ghyselinck, N.B. The retinoic acid receptors raralpha and rargamma are required for inner ear development. Mech. Dev. 2002, 119, 213–223. [Google Scholar] [CrossRef]
- Park, J.S.; Kang, S.J.; Seo, M.K.; Jou, I.; Woo, H.G.; Park, S.M. Role of cysteinyl leukotriene signaling in a mouse model of noise-induced cochlear injury. Proc. Natl. Acad. Sci. USA 2014, 111, 9911–9916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.; Yoo, J.E.; Choe, Y.H.; Park, S.C.; Lee, H.J.; Lee, H.J.; Noh, B.; Kim, S.H.; Kang, G.Y.; Lee, K.M.; et al. Cleaved cochlin sequesters pseudomonas aeruginosa and activates innate immunity in the inner ear. Cell Host Microbe 2019, 25, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, S.C.; Cox, B.C. Whole mount dissection and immunofluorescence of the adult mouse cochlea. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, S.G.; Liberman, M.C. Adding insult to injury: Cochlear nerve degeneration after “temporary” noise-induced hearing loss. J. Neurosci. 2009, 29, 14077–14085. [Google Scholar] [CrossRef] [PubMed]
- Lobarinas, E.; Spankovich, C.; Le Prell, C.G. Evidence of “hidden hearing loss” following noise exposures that produce robust tts and abr wave-i amplitude reductions. Hear. Res. 2017, 349, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Budden, R.; Kuhl, U.G.; Buschmann, G. Studies on the pharmacodynamic activity of several drug solvents. 1st communication: Diethyleneglycol monoethylether, n,n-diethylacetamide, dimethylsufoxide. Arzneimittel-Forschung 1978, 28, 1571–1579. [Google Scholar] [PubMed]
- Pestel, S.; Martin, H.J.; Maier, G.M.; Guth, B. Effect of commonly used vehicles on gastrointestinal, renal, and liver function in rats. J. Pharmacol. Toxicol. Method. 2006, 54, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Clarke, E.; Jarvis, C.I.; Goncalves, M.B.; Kalindjian, S.B.; Adams, D.R.; Brown, J.T.; Shiers, J.J.; Taddei, D.M.A.; Ravier, E.; Barlow, S.; et al. Design and synthesis of a potent, highly selective, orally bioavailable, retinoic acid receptor alpha agonist. Bioorg. Med. Chem. 2018, 26, 798–814. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Bertran, S.; Samuels, T.A.; Mira-y-Lopez, R.; Farias, E.F. Mechanism of inhibition of mmtv-neu and mmtv-wnt1 induced mammary oncogenesis by raralpha agonist am580. Oncogene 2010, 29, 3665–3676. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Dasari, S.; Nowakowski, G.S.; Lazaridis, K.N.; Wieben, E.D.; Kadin, M.E.; Feldman, A.L.; Boddicker, R.L. Retinoic acid receptor alpha drives cell cycle progression and is associated with increased sensitivity to retinoids in t-cell lymphoma. Oncotarget 2017, 8, 26245–26255. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, H.; Hijioka, M.; Hisatsune, A.; Isohama, Y.; Shudo, K.; Katsuki, H. A retinoic acid receptor agonist am80 rescues neurons, attenuates inflammatory reactions, and improves behavioral recovery after intracerebral hemorrhage in mice. J. Cerebr. Blood Flow Metab. 2011, 31, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Wei-Ju, H.A.N.; Xiao-Rui, S.H.I.; Nuttall, A. Noise-induced nitrotyrosine increase and outer hair cell death in guinea pig cochlea. Chin. Med. J. 2013, 126, 2923–2927. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Hearing Threshold (dB SPL) | Wave I Amplitude (nV) | Hair Cell Survival (%) |
|---|---|---|---|
| Control | 92.1 ± 16.8 | 143.2 ± 365.9 | 88.2 ± 7.6 |
| ATRA | 65.0 ± 22.1 * | 471.7 ± 455.7 | 99.7 ± 1.1 *** |
| AM80 | 58.6 ± 9.1 * | 637.5 ± 515.7 * | 97.4 ± 2.5 * |
| AC261066 | 64.6 ± 20.4 * | 538.1 ± 452.2 | 96.5 ± 6.1 * |
| CD1530 | 62.9 ± 23.1 * | 587.4 ± 561.5 * | 97.3 ± 3.4 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwak, S.H.; Nam, G.-S.; Bae, S.H.; Jung, J. Effect of Specific Retinoic Acid Receptor Agonists on Noise-Induced Hearing Loss. Int. J. Environ. Res. Public Health 2019, 16, 3428. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16183428
Kwak SH, Nam G-S, Bae SH, Jung J. Effect of Specific Retinoic Acid Receptor Agonists on Noise-Induced Hearing Loss. International Journal of Environmental Research and Public Health. 2019; 16(18):3428. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16183428
Chicago/Turabian StyleKwak, Sang Hyun, Gi-Sung Nam, Seong Hoon Bae, and Jinsei Jung. 2019. "Effect of Specific Retinoic Acid Receptor Agonists on Noise-Induced Hearing Loss" International Journal of Environmental Research and Public Health 16, no. 18: 3428. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16183428