Sulforaphane Protects the Male Reproductive System of Mice from Obesity-Induced Damage: Involvement of Oxidative Stress and Autophagy

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Diet
2.4. Diet-Induced Obesity (DIO) Definition
2.5. SFN Exposure
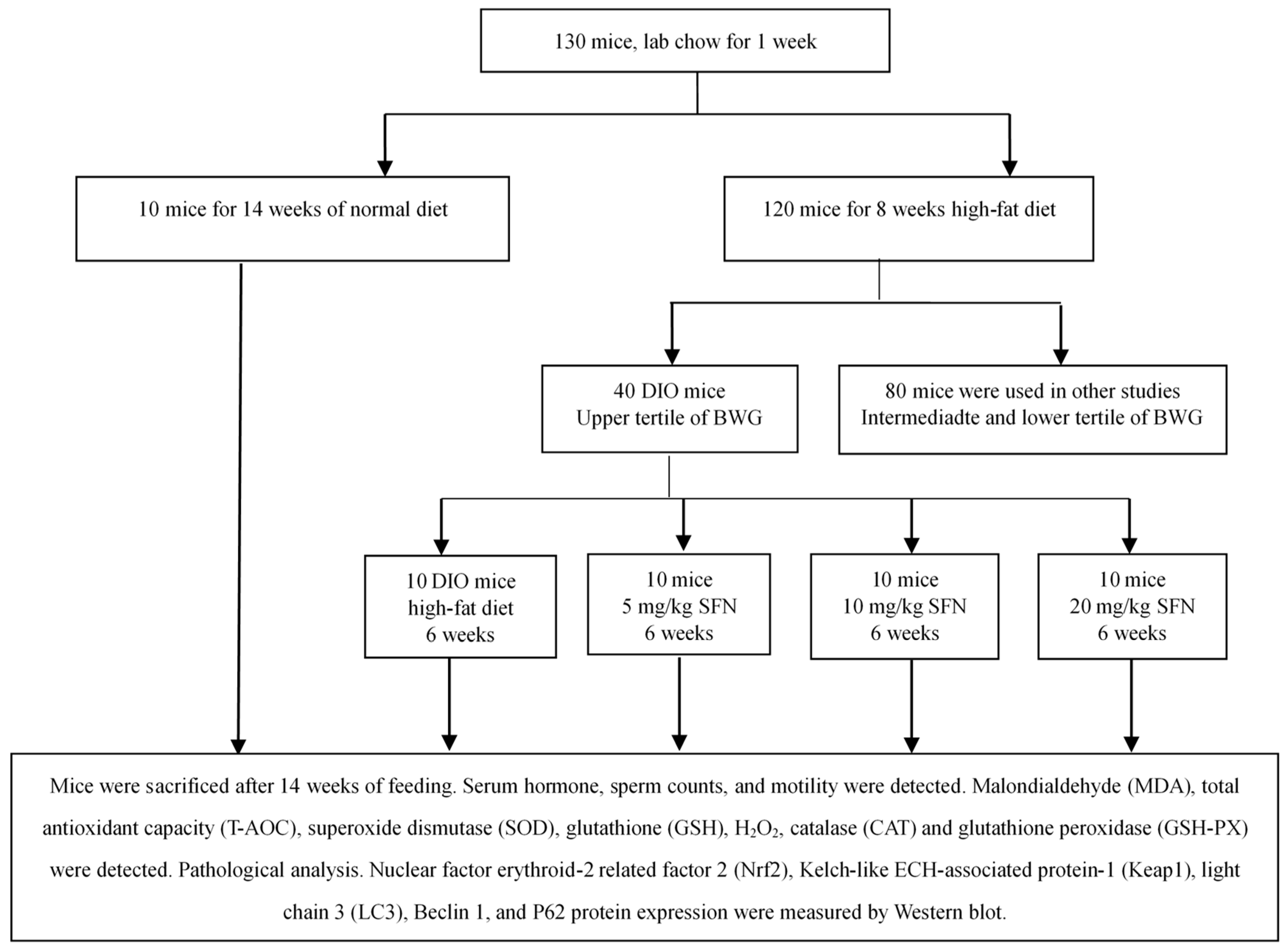
2.6. Grouping
2.7. Experimental Procedure
2.8. Tissue Processing
2.9. Epididymal Sperm Count and Motility
2.10. Light Microscopy
2.11. Transmission Electron Microscope
2.12. Hormone Measurements
2.13. Biochemical Parameters
2.14. Protein Preparation from Testes
2.15. Western Blotting
2.16. Statistical Analyses
3. Results
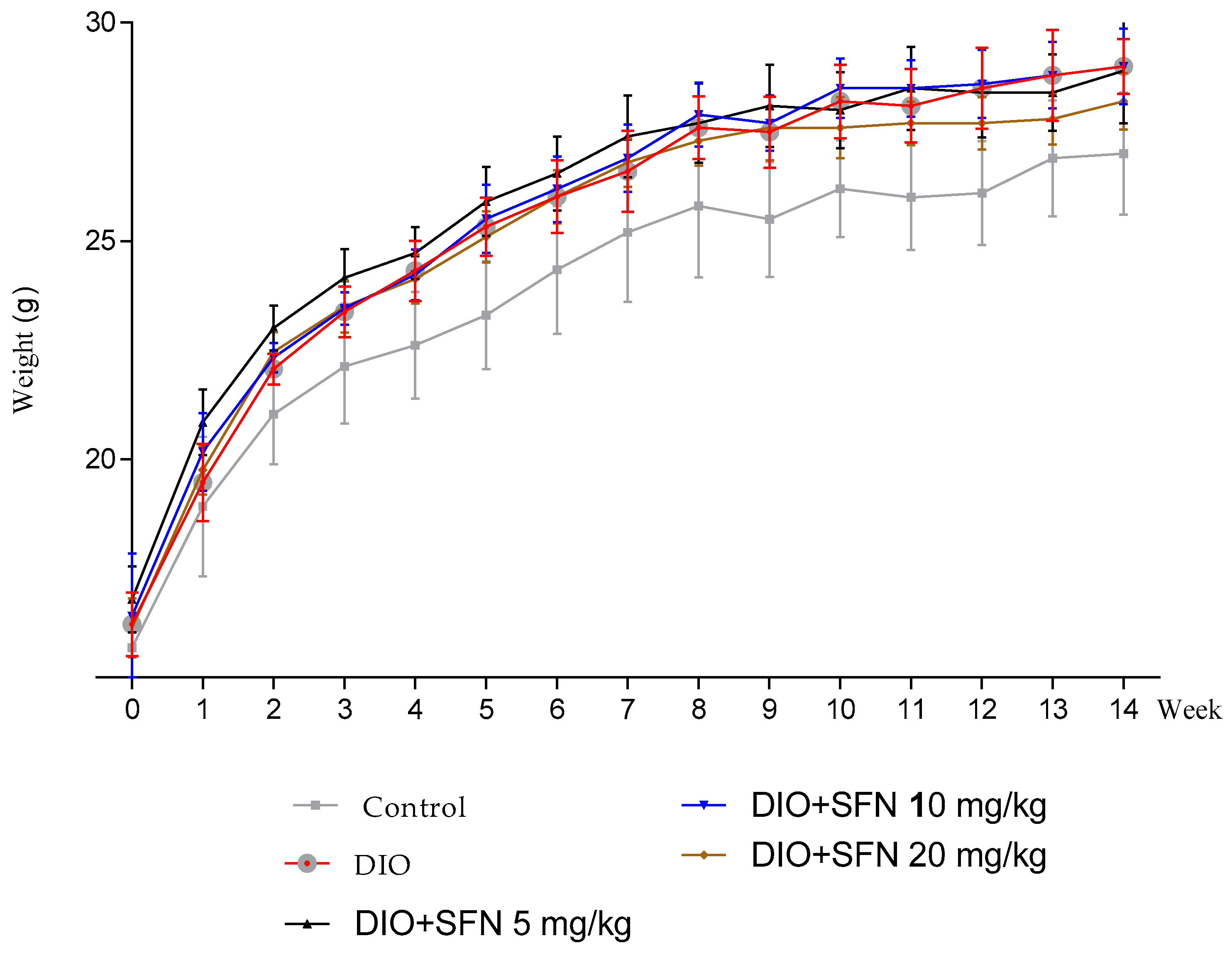
3.1. Body Weight
3.2. Effects of SFN on Reproductive Organs Index and Sperm Characteristics in Obese Mice
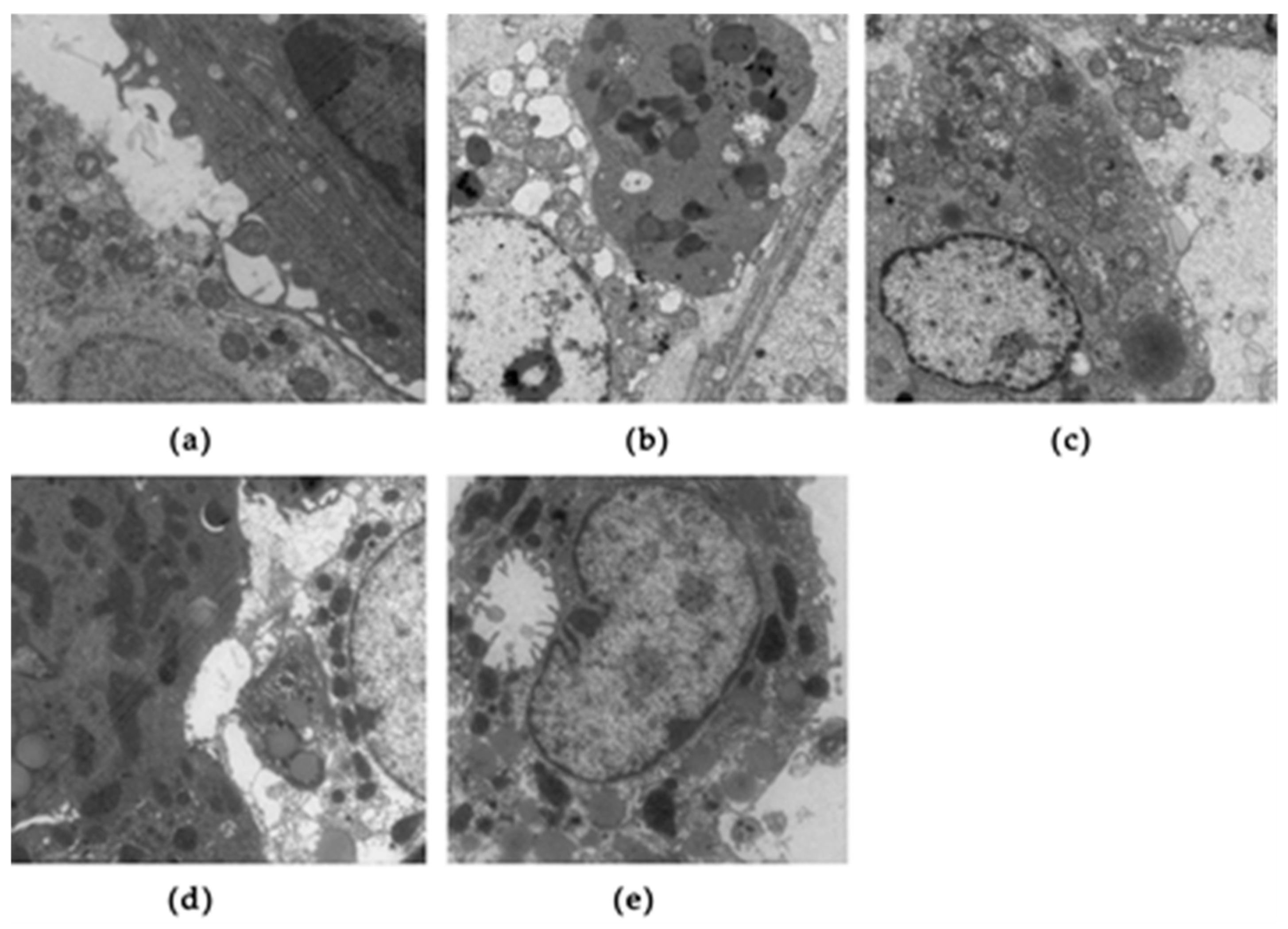
3.3. Effects of SFN on Testicular Histology in Obese Mice
3.4. Effects of SFN on Serum Hormone Levels in Obese Mice
3.5. Effects of SFN on Biochemical Parameters Related with Oxidative Stress in Obese Mice
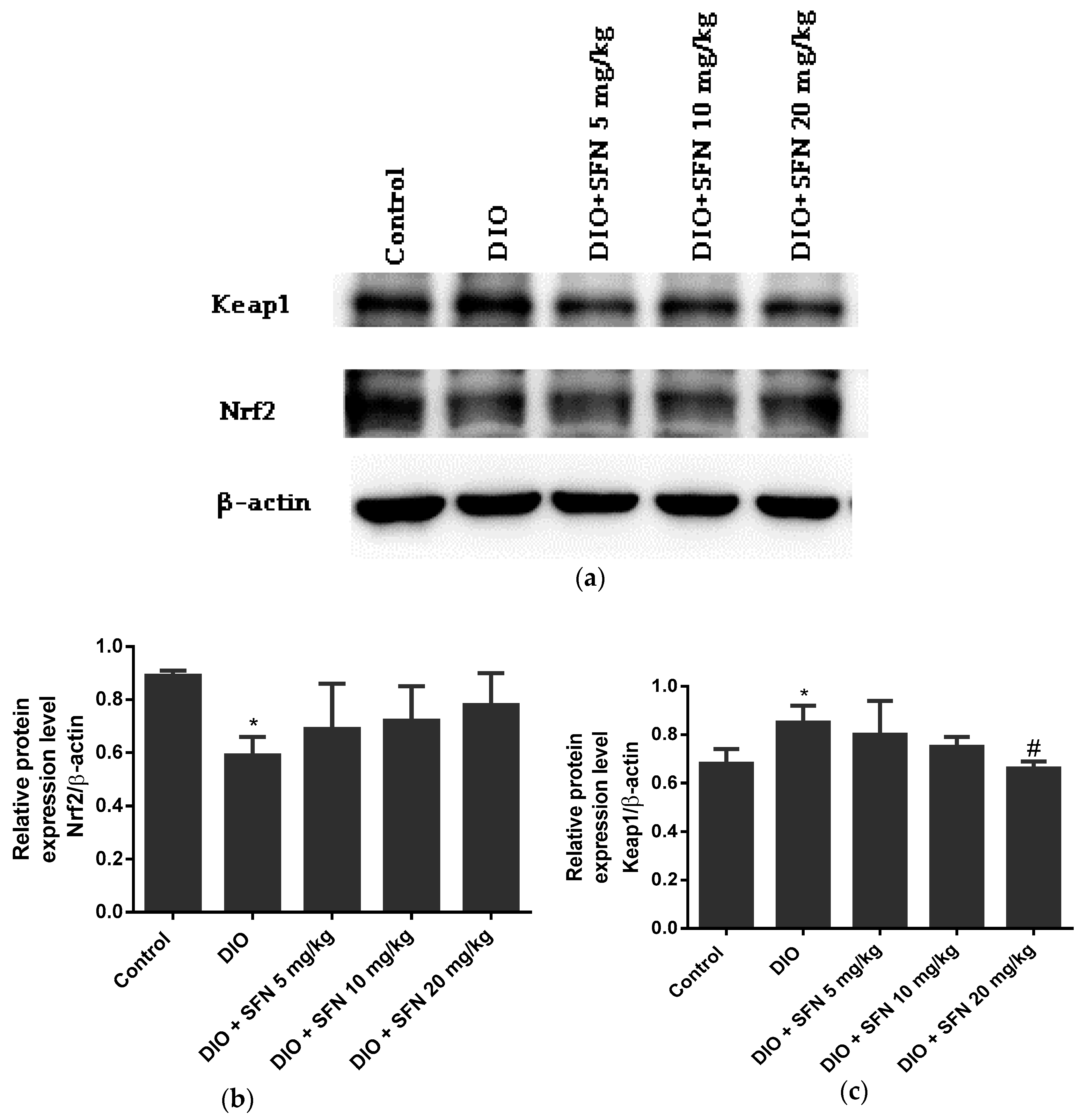
3.6. Effects of SFN on the Protein Expression in Obese Mice
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- NCD Risk Factor Collaboration (NCD-RisC). Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef]
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity Among Adults and Youth: United States, 2015–2016. NCHS. Data. Brief. 2017, 288, 1–8. [Google Scholar]
- Aly, J.M.; Polotsky, A.J. Paternal Diet and Obesity: Effects on Reproduction. Semin. Reprod. Med. 2017, 35, 313–317. [Google Scholar] [PubMed]
- McPherson, N.O.; Fullston, T.; Aitken, R.J.; Lane, M. Paternal obesity, interventions, and mechanistic pathways to impaired health in offspring. Ann. Nutr. Metab. 2014, 64, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Soubry, A.; Guo, L.; Huang, Z.; Hoyo, C.; Romanus, S.; Price, T.; Murphy, S.K. Obesity-related DNA methylation at imprinted genes in human sperm: Results from the TIEGER study. Clin. Epigenetics 2016, 8, 51. [Google Scholar] [CrossRef]
- Calderon, B.; Gomez-Martin, J.M.; Vega-Pinero, B.; Martin-Hidalgo, A.; Galindo, J.; Luque-Ramirez, M.; Escobar-Morreale, H.F.; Botella-Carretero, J.I. Prevalence of male secondary hypogonadism in moderate to severe obesity and its relationship with insulin resistance and excess body weight. Andrology 2016, 4, 62–67. [Google Scholar] [CrossRef]
- Escobar-Morreale, H.F.; Santacruz, E.; Luque-Ramirez, M.; Botella Carretero, J.I. Prevalence of ‘obesity-associated gonadal dysfunction’ in severely obese men and women and its resolution after bariatric surgery: A systematic review and meta-analysis. Hum. Reprod. Update 2017, 23, 390–408. [Google Scholar] [CrossRef]
- Eriksson, J.; Haring, R.; Grarup, N.; Vandenput, L.; Wallaschofski, H.; Lorentzen, E.; Hansen, T.; Mellstrom, D.; Pedersen, O.; Nauck, M.; et al. Causal relationship between obesity and serum testosterone status in men: A bi-directional mendelian randomization analysis. PLoS ONE 2017, 12, e0176277. [Google Scholar] [CrossRef]
- Makary, S.; Abdo, M.; Fekry, E. Oxidative stress burden inhibits spermatogenesis in adult male rats: Testosterone protective effect. Can. J. Physiol. Pharmacol. 2017, 96, 372–381. [Google Scholar] [CrossRef]
- Khosropour, S.; Hamidi, M.; Fattahi, A.; Khodadadi, I.; Karami, M.; Fazilati, M.; Vaisi-Raygani, A.; Tavilani, H. Leptin and leptin-receptor polymorphisms in fertile and infertile men. Syst. Biol. Reprod. Med. 2017, 63, 7–14. [Google Scholar] [CrossRef]
- Zhao, J.; Zhai, L.; Liu, Z.; Wu, S.; Xu, L. Leptin level and oxidative stress contribute to obesity-induced low testosterone in murine testicular tissue. Oxid. Med. Cell. Longev. 2014, 2014, 190945. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alahmar, A.T. Role of Oxidative Stress in Male Infertility: An Updated Review. J. Hum. Reprod. Sci. 2019, 12, 4–18. [Google Scholar] [CrossRef] [PubMed]
- D’Errico, G.; Vitiello, G.; De Tommaso, G.; Abdel-Gawad, F.K.; Brundo, M.V.; Ferrante, M.; De Maio, A.; Trocchia, S.; Bianchi, A.R.; Ciarcia, G.; et al. Electron Spin Resonance (ESR) for the study of Reactive Oxygen Species (ROS) on the isolated frog skin (Pelophylax bergeri): A non-invasive method for environmental monitoring. Environ. Res. 2018, 165, 11–18. [Google Scholar]
- Guerriero, G.; Parisi, C.; Abdel-Gawad, F.K.; Hentati, O.; D’Errico, G. Seasonal and pharmaceutical- induced changes in selenoprotein glutathione peroxidase 4 activity in the reproductive dynamics of the soil biosentinel Podarcis sicula (Chordata: Reptilia). Mol. Reprod. Dev. 2019. Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Weide, T.; Huber, T.B. Implications of autophagy for glomerular aging and disease. Cell. Tissue. Res. 2011, 343, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jiang, M.; Bi, Y.; Zhu, H.; Zhou, Z.; Sha, J. Autophagy and apoptosis act as partners to induce germ cell death after heat stress in mice. PLoS ONE 2012, 7, e41412. [Google Scholar] [CrossRef]
- Wang, H.; Wan, H.; Li, X.; Liu, W.; Chen, Q.; Wang, Y.; Yang, L.; Tang, H.; Zhang, X.; Duan, E.; et al. Atg7 is required for acrosome biogenesis during spermatogenesis in mice. Cell. Res. 2014, 24, 852–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebato, C.; Uchida, T.; Arakawa, M.; Komatsu, M.; Ueno, T.; Komiya, K.; Azuma, K.; Hirose, T.; Tanaka, K.; Kominami, E.; et al. Autophagy is important in islet homeostasis and compensatory increase of beta cell mass in response to high-fat diet. Cell. Metab. 2008, 8, 325–332. [Google Scholar] [CrossRef]
- Rhee, J.S.; Saben, J.L.; Mayer, A.L.; Schulte, M.B.; Asghar, Z.; Stephens, C.; Chi, M.M.; Moley, K.H. Diet-induced obesity impairs endometrial stromal cell decidualization: A potential role for impaired autophagy. Hum. Reprod. 2016, 31, 1315–1326. [Google Scholar] [CrossRef]
- Jiang, X.P.; Tang, J.Y.; Xu, Z.; Han, P.; Qin, Z.Q.; Yang, C.D.; Wang, S.Q.; Tang, M.; Wang, W.; Qin, C.; et al. Sulforaphane attenuates di-N-butylphthalate-induced reproductive damage in pubertal mice: Involvement of the Nrf2-antioxidant system. Environ. Toxicol. 2017, 32, 1908–1917. [Google Scholar] [CrossRef]
- Yang, S.H.; Long, M.; Yu, L.H.; Li, L.; Li, P.; Zhang, Y.; Guo, Y.; Gao, F.; Liu, M.D.; He, J.B. Sulforaphane Prevents Testicular Damage in Kunming Mice Exposed to Cadmium via Activation of Nrf2/ARE Signaling Pathways. Int. J. Mol. Sci. 2016, 17, 1703. [Google Scholar] [CrossRef]
- Zhao, J.; Ren, S.; Liu, C.; Huo, L.; Liu, Z.; Zhai, L. Di-(2-Ethylhexyl) Phthalate Increases Obesity-Induced Damage to the Male Reproductive System in Mice. Oxid. Med. Cell. Longev. 2018, 2018, 1861984. [Google Scholar] [CrossRef]
- Levin, B.E.; Keesey, R.E. Defense of differfing body weight set points in diet-induced obese and resistant rats. AM. J. Physiol. 1998, 274, R412–R419. [Google Scholar] [CrossRef]
- Organization, W.H. Who laboratory manual for the examination of human semen and sperm-cervical mucus interaction. J. Androl. 1996, 17, 442. [Google Scholar]
- Erdemir, F.; Atilgan, D.; Markoc, F.; Boztepe, O.; Suha-Parlaktas, B.; Sahin, S. The effect of diet induced obesity on testicular tissue and serum oxidative stress parameters. Actas. Urol. Esp. 2012, 36, 153–159. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, A.A.; Herbison, G.P.; Showell, M.; Farquhar, C.M. The impact of body mass index on semen parameters and reproductive hormones in human males: A systematic review with meta-analysis. Hum. Reprod. Update 2010, 16, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.D.; Moreira, A.C.; Sa, R.; Monteiro, M.P.; Sousa, M.; Carvalho, R.A.; Silva, B.M.; Oliveira, P.F.; Alves, M.G. Leptin modulates human Sertoli cells acetate production and glycolytic profile: A novel mechanism of obesity-induced male infertility? Biochim. Biophys. Acta 2015, 1852, 1824–1832. [Google Scholar] [CrossRef] [PubMed]
- McPherson, N.O.; Lane, M. Male obesity and subfertility, is it really about increased adiposity? Asian. J. Androl. 2015, 17, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Niederberger, C. Re: A Randomized Prospective Double-Blind Comparison Trial of Clomiphene Citrate and Anastrozole in Raising Testosterone in Hypogonadal Infertile Men. J. Urol. 2016, 195, 1075–1077. [Google Scholar] [CrossRef]
- Sinclair, M.; Grossmann, M.; Gow, P.J.; Angus, P.W. Testosterone in men with advanced liver disease: Abnormalities and implications. J. Gastroenterol. Hepatol. 2015, 30, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Palmer, N.O.; Bakos, H.W.; Fullston, T.; Lane, M. Impact of obesity on male fertility, sperm function and molecular composition. Spermatogenesis 2012, 2, 253–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristian Leisegang, P.J.B.; Menkveld, R.; Henkel, R.R. Obesity is associated with increased seminal insulin and leptin alongside reduced fertility parameters in a controlled male cohort. Reprod. Biol. Endocrin. 2014, 12, 34. [Google Scholar] [CrossRef] [PubMed]
- Shawky, N.M.; Pichavaram, P.; Shehatou, G.S.; Suddek, G.M.; Gameil, N.M.; Jun, J.Y.; Segar, L. Sulforaphane improves dysregulated metabolic profile and inhibits leptin-induced VSMC proliferation: Implications toward suppression of neointima formation after arterial injury in western diet-fed obese mice. J. Nutr. Biochem. 2016, 32, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, H.; Misro, M.M.; Sharma, S.B.; Prakash, S. Cytoprotective effects of fruit pulp of Eugenia jambolana on H2O2-induced oxidative stress and apoptosis in rat Leydig cells in vitro. Andrologia 2013, 45, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.T.; Lysiak, J.J. Oxidative stress: A common factor in testicular dysfunction. J. Androl. 2008, 29, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Justiniano, R.; Zhang, D.D.; Wondrak, G.T. The Nrf2-inducers tanshinone I and dihydrotanshinone protect human skin cells and reconstructed human skin against solar simulated UV. Redox. Biol. 2013, 1, 532–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef]
- Mutter, F.E.; Park, B.K.; Copple, I.M. Value of monitoring Nrf2 activity for the detection of chemical and oxidative stress. Biochem. Soc.Trans. 2015, 43, 657–662. [Google Scholar] [CrossRef] [Green Version]
- Niture, S.K.; Kaspar, J.W.; Shen, J.; Jaiswal, A.K. Nrf2 signaling and cell survival. Toxicol. Appl. Pharmacol. 2010, 244, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Araya, J.; Hara, H.; Kuwano, K. Autophagy in the pathogenesis of pulmonary disease. Intern. Med. 2013, 52, 2295–2303. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhao, J.; Liu, Q.; Diao, X.; Kong, B. Sulforaphane protects human umbilical vein cells against lipotoxicity by stimulating autophagy via an AMPK-mediated pathway. J. Funct. Foods 2015, 15, 23–34. [Google Scholar] [CrossRef]
- Xu, Z.; Yang, L.; Xu, S.; Zhang, Z.; Cao, Y. The receptor proteins: Pivotal roles in selective autophagy. Acta. Biochim. Biophys. Sin. 2015, 47, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, I.M.; Espino, J.; Bejarano, I.; Gallardo-Soler, A.; Campo, M.L.; Salido, G.M.; Pariente, J.A.; Pena, F.J.; Tapia, J.A. Autophagy-related proteins are functionally active in human spermatozoa and may be involved in the regulation of cell survival and motility. Sci. Rep. 2016, 6, 33647. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.C.; Chung, S.D.; Chien, C.T.; Yu, H.J. Sulforaphane Improves Ischemia-Induced Detrusor Overactivity by Downregulating the Enhancement of Associated Endoplasmic Reticulum Stress, Autophagy, and Apoptosis in Rat Bladder. Sci. Rep. 2016, 6, 36110. [Google Scholar] [CrossRef]
- Zhang, H.; Kong, X.; Kang, J.; Su, J.; Li, Y.; Zhong, J.; Sun, L. Oxidative stress induces parallel autophagy and mitochondria dysfunction in human glioma U251 cells. Toxicol. Sci. 2009, 110, 376–388. [Google Scholar] [CrossRef]
- Hariharan, N.; Zhai, P.; Sadoshima, J. Oxidative stress stimulates autophagic flux during ischemia/reperfusion. Antioxid. Redox. Signal. 2011, 14, 2179–2190. [Google Scholar] [CrossRef]
- Essick, E.E.; Wilson, R.M.; Pimentel, D.R.; Shimano, M.; Baid, S.; Ouchi, N.; Sam, F. Adiponectin modulates oxidative stress-induced autophagy in cardiomyocytes. PLoS ONE 2013, 8, e68697. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n | Relative Tes.Weight (g/100g) | Relative Epididymis Weight (g/100) | Relative Sem Weight (g/100g) | Relative Liver Weight (g/100g) | Relative Kidney Weight (g/100g) | Relative Epi Fat (g/100g) | Relative Ret Fat (g/100g) | Sperm Motility (%) | Sperm Count (×106/mL) |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 10 | 0.84 ± 0.09 | 0.30 ± 0.03 | 0.95 ± 0.10 | 4.44 ± 0.24 | 1.21 ± 0.07 | 1.04 ± 0.21 | 0.19 ± 0.09 | 30.50 ± 2.22 | 4.91 ± 0.28 |
| DIO | 10 | 0.74 ± 0.04 ** | 0.28 ± 0.02 * | 1.0 ± 0.11 | 4.18 ± 0.17 ** | 1.15 ± 0.06 * | 1.42 ± 0.25 ** | 0.37 ± 0.08 ** | 21.50 ± 1.97 ** | 3.72 ± 0.28 ** |
| DIO + SFN 5 mg/kg | 10 | 0.74 ± 0.03 | 0.28 ± 0.03 | 0.98 ± 0.15 | 4.36± 0.17 # | 1.17 ± 0.03 | 1.48 ± 0.15 | 0.37 ± 0.09 | 23.80 ± 2.10 # | 4.04 ± 0.40 ## |
| DIO + SFN 10 mg/kg | 10 | 0.76 ± 0.04 | 0.30 ± 0.04 # | 0.95 ± 0.13 | 4.43 ± 0.15 ## | 1.18 ± 0.07 | 1.35 ± 0.18 | 0.35 ± 0.10 | 29.10 ± 1.61 ## | 4.79 ± 0.16 ## |
| DIO + SFN 20 mg/kg | 10 | 0.77 ± 0.04 | 0.31 ± 0.03 ## | 0.97 ± 0.13 | 4.48 ± 0.20 ## | 1.22 ± 0.07 ## | 1.33 ± 0.11 | 0.24 ± 0.09 ## | 31.20 ± 2.96 ## | 4.76 ± 0.55 ## |
| Group | n | Leptin (ng/mL) | Testosterone (pg/mL) | Estradiol (pg/mL) |
|---|---|---|---|---|
| Control | 10 | 1.56 ± 0.37 | 4724.28 ± 619.50 | 14.15 ± 3.24 |
| DIO | 10 | 4.46 ± 1.35 ** | 3580.00 ± 808.21 ** | 19.10 ± 2.04 ** |
| DIO + SFN 5 mg/kg | 10 | 3.70 ± 1.41 | 4622.02 ± 864.37# | 18.31 ± 3.43 |
| DIO + SFN 10 mg/kg | 10 | 3.62 ± 0.86 | 4651.58 ± 905.40# | 16.10 ± 3.15# |
| DIO + SFN 20 mg/kg | 10 | 3.49 ± 0.60# | 4865.13 ± 519.91## | 14.27 ± 3.00## |
| Group | n | SOD (U/mg prot) | CAT (U/mg prot) | GSH-Px (U/mg prot) | T-AOC (U/mg prot) | GSH (mg/g prot) | H2O2 (mmol/gprot) | MDA (nmol/mg prot) |
|---|---|---|---|---|---|---|---|---|
| Control | 10 | 234.65 ± 28.81 | 14.40 ± 1.69 | 57.27 ± 10.14 | 0.70 ± 0.21 | 146.23 ± 16.63 | 4.53 ± 0.72 | 1.48 ± 0.68 |
| DIO | 10 | 171.76 ± 40.66 ** | 13.70 ± 2.20 | 44.97 ± 10.71 * | 0.59 ± 0.22 | 100.89 ± 16.34 ** | 18.13 ± 3.20 ** | 6.37 ± 1.11 ** |
| DIO + SFN 5 mg/kg | 10 | 234.51 ± 38.57 ## | 13.90 ± 2.70 | 45.14 ± 8.03 | 0.61 ± 0.11 | 94.62 ± 12.30 | 16.11 ± 4.61 | 5.64 ± 0.91 |
| DIO + SFN 10 mg/kg | 10 | 249.96 ± 40.29 ## | 15.75 ± 2.93 | 46.14 ± 9.01 | 0.61 ± 0.19 | 104.98 ± 20.15 | 16.57 ± 2.98 | 5.82 ± 1.35 |
| DIO + SFN 20mg/kg | 10 | 251.52 ± 45.25 ## | 15.41 ± 2.85 | 46.48 ± 9.65 | 0.62 ± 0.18 | 112.63 ± 19.54 | 16.25 ± 2.72 | 4.65 ± 1.09 ## |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, L.; Su, Y.; Xu, G.; Zhai, L.; Zhao, J. Sulforaphane Protects the Male Reproductive System of Mice from Obesity-Induced Damage: Involvement of Oxidative Stress and Autophagy. Int. J. Environ. Res. Public Health 2019, 16, 3759. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16193759
Huo L, Su Y, Xu G, Zhai L, Zhao J. Sulforaphane Protects the Male Reproductive System of Mice from Obesity-Induced Damage: Involvement of Oxidative Stress and Autophagy. International Journal of Environmental Research and Public Health. 2019; 16(19):3759. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16193759
Chicago/Turabian StyleHuo, Li, Yu Su, Gaoyang Xu, Lingling Zhai, and Jian Zhao. 2019. "Sulforaphane Protects the Male Reproductive System of Mice from Obesity-Induced Damage: Involvement of Oxidative Stress and Autophagy" International Journal of Environmental Research and Public Health 16, no. 19: 3759. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16193759