Aerobic and Anaerobic Biodegradation of 1,2-Dibromoethane by a Microbial Consortium under Simulated Groundwater Conditions

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cultivation of Microorganisms
2.3. Microcosm Study
2.4. DNA Extraction and 16S rRNA Analysis
2.5. Analytical Methods
3. Results and Discussion
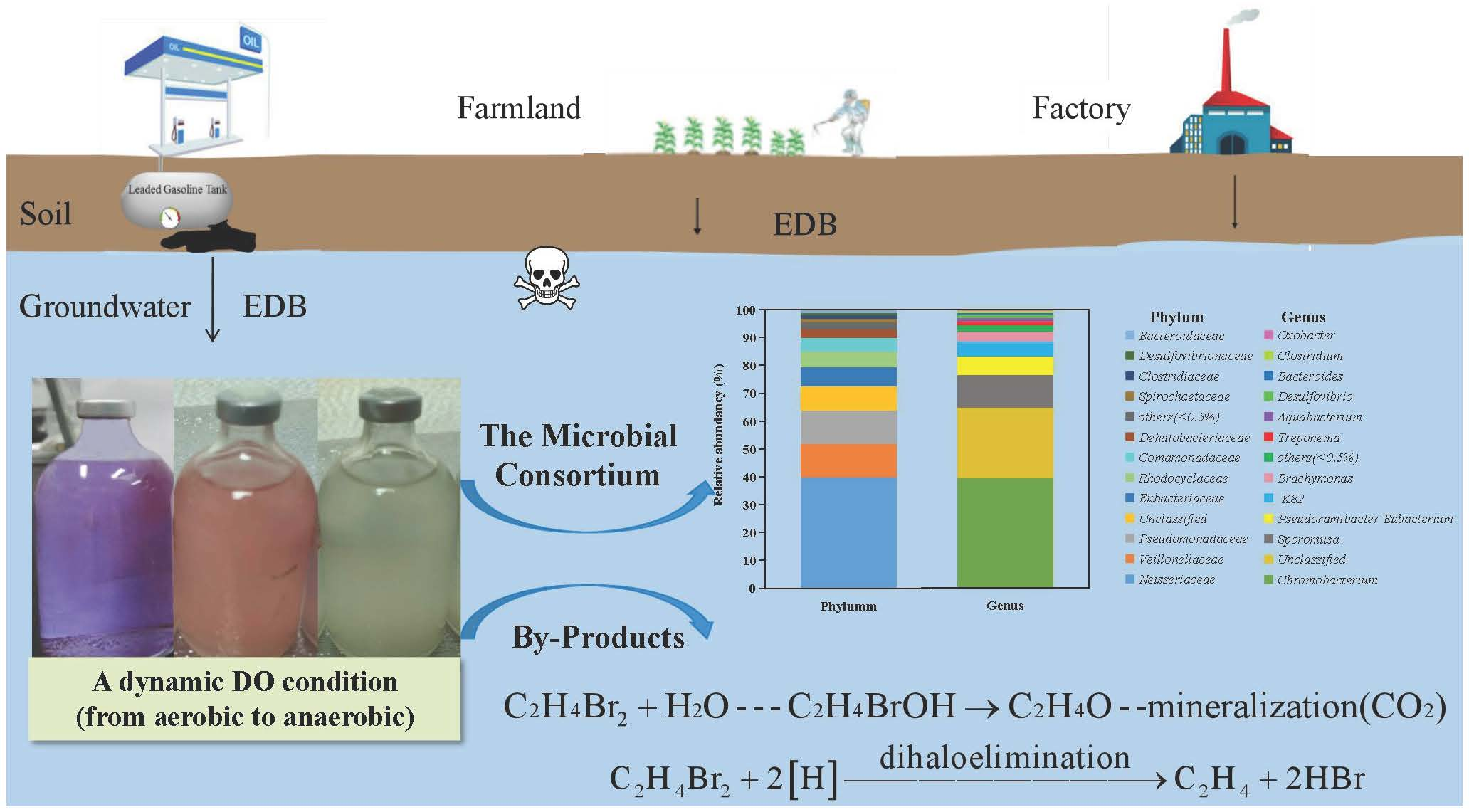
3.1. Composition of the Acclimated Microbial Consortium
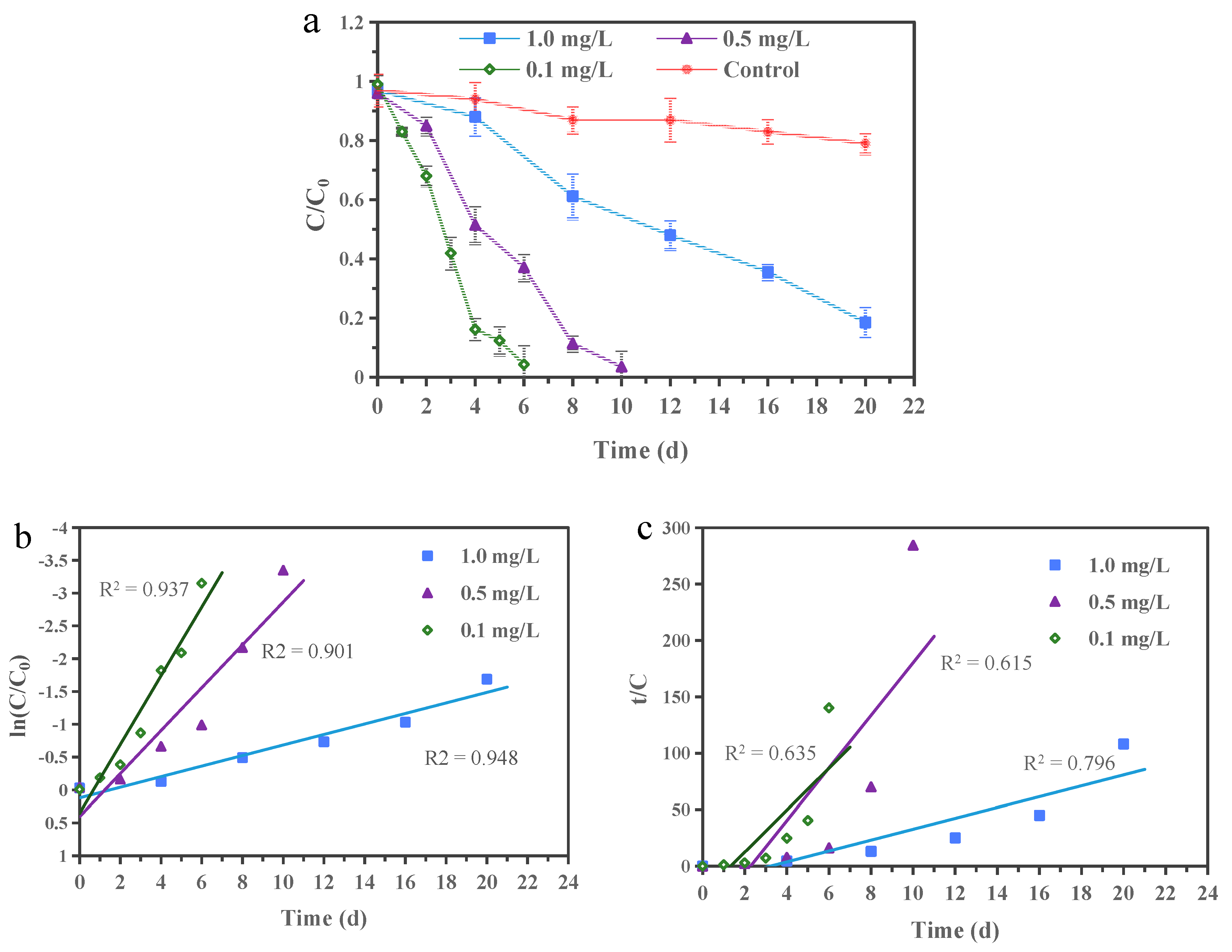
3.2. EDB Biodegradation
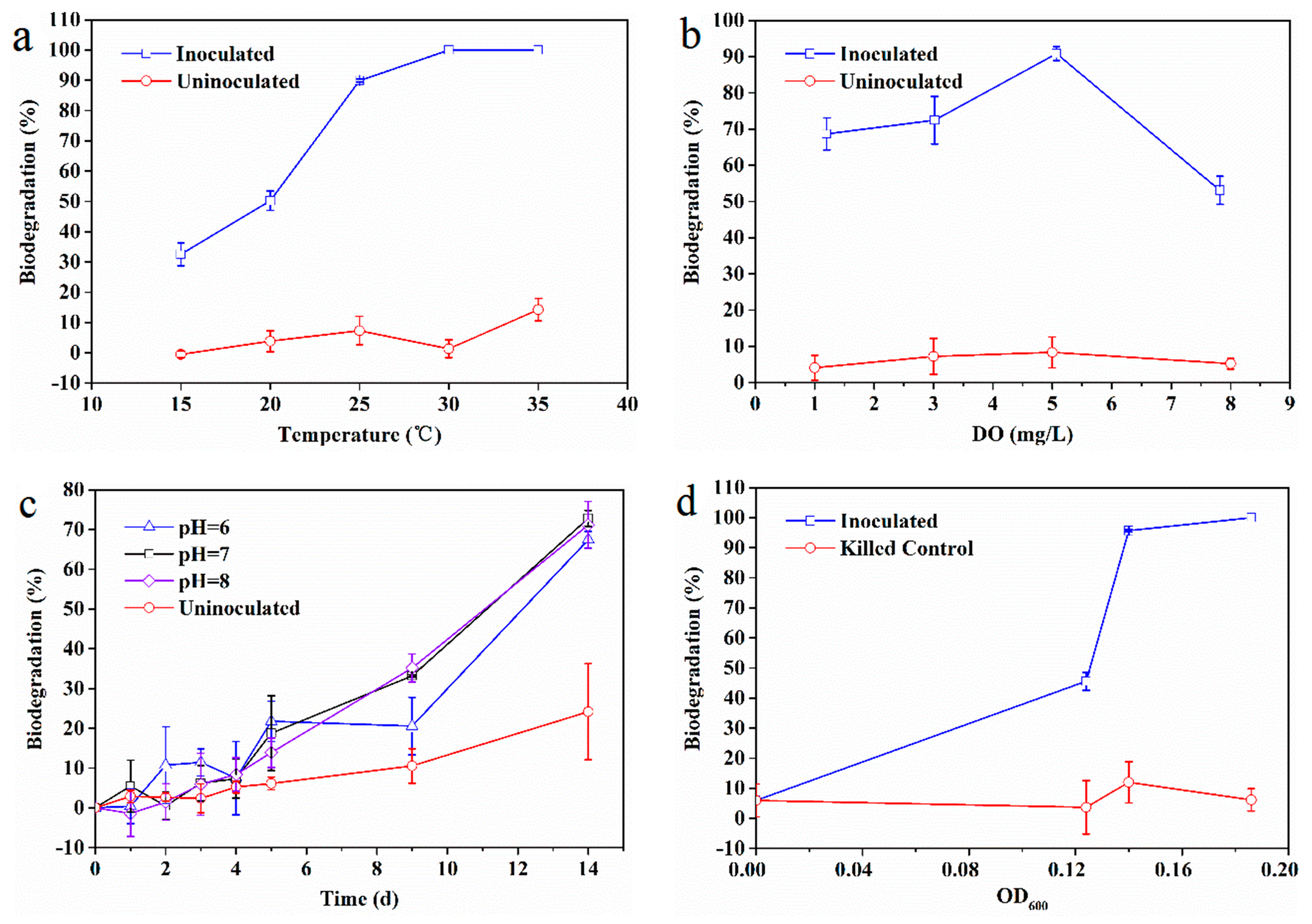
3.3. Influence of Environment Factors on EDB Biodegradation
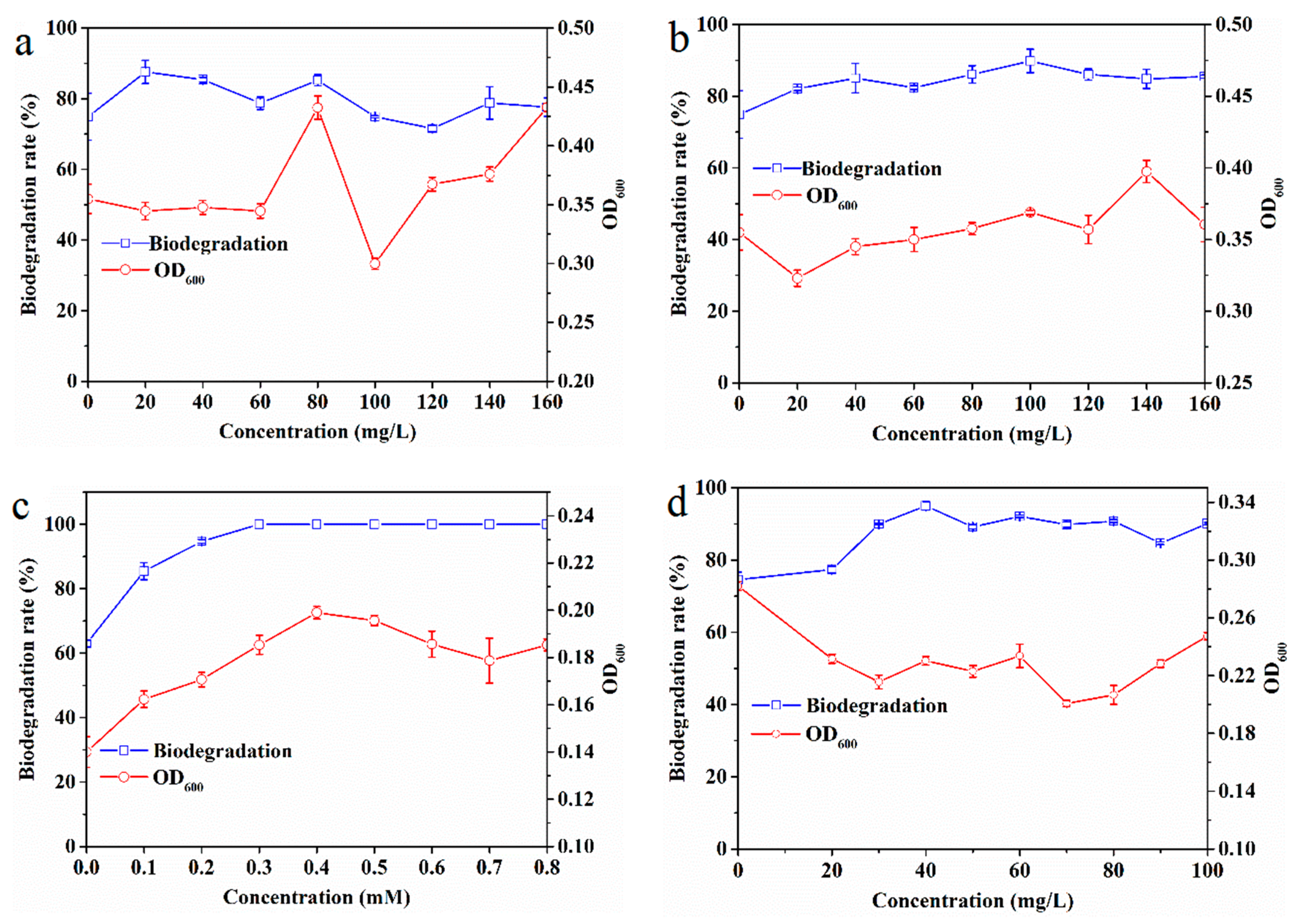
3.4. Effects of Co-Substrates and Surfactant on EDB Biodegradation
3.5. Potential EDB Biodegradation Pathways
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Falta, R.W.; Bulsara, N.; Henderson, J.K.; Mayer, R.A. Leaded-gasoline ADDITIVES still contaminate groundwater. Environ. Sci. Technol. 2005, 39, 378A–384A. [Google Scholar] [CrossRef] [PubMed]
- McKeever, R.; Sheppard, D.; Nuesslein, K.; Baek, K.-H.; Rieber, K.; Ergas, S.J.; Forbes, R.; Hilyard, M.; Park, C. Biodegradation of ethylene dibromide (1,2-dibromoethane EDBß) in microcosms simulating in situ and biostimulated conditions. J. Hazard. Mater. 2012, 209, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Falta, R.W. The potential for ground water contamination by the gasoline lead scavengers ethylene dibromide and 1,2-dichloroethane. Groundw. Monit. Remediat. 2004, 24, 76–87. [Google Scholar] [CrossRef]
- Pignatello, J.J.; Cohen, S.Z. Environmental chemistry of ethylene dibromide in soil and ground water. Rev. Environ. Contam. Toxicol. 1990, 112, 1–47. [Google Scholar] [PubMed]
- Wang, Y. Prohibited pesticide in China. Pestic. Sci. Admin. 2003, 24, 42–43. [Google Scholar]
- Xin, M. Study on the Residue Methods and Degradation Dynamical Rhythm of Ethylene Dibromide Pesticide in Fruit and Canned Fruit. Master thesis, Guangxi University, Nanning, China, 2011. [Google Scholar]
- Hatzinger, P.B.; Streger, S.H.; Begley, J.F. Enhancing aerobic biodegradation of 1,2-dibromoethane in groundwater using ethane or propane and inorganic nutrients. J. Contam. Hydrol. 2015, 172, 61–70. [Google Scholar] [CrossRef]
- Rajagopal, V.K.; Burris, D.R. Reduction of 1,2-dibromoethane in the presence of zero-valent iron. Environ. Toxicol. Chem. 1999, 18, 1779–1782. [Google Scholar] [CrossRef]
- Kuder, T.; Wilson, J.T.; Philp, P.; He, Y.T. Carbon Isotope Fractionation in Reactions of 1,2-Dibromoethane with FeS and Hydrogen Sulfide. Environ. Sci. Technol. 2012, 46, 7495–7502. [Google Scholar] [CrossRef]
- van Groos, P.G.K.; Hatzinger, P.B.; Streger, S.H.; Vainberg, S.; Philp, R.P.; Kuder, T. Carbon Isotope Fractionation of 1,2-Dibromoethane by Biological and Abiotic Processes. Environ. Sci. Technol. 2018, 52, 3440–3448. [Google Scholar] [CrossRef]
- Nguyen, T.; Ollis, D.F. Complete heterogeneously photocatalyzed transformation of 1,1- and 1,2-dibromoethane to carbon dioxide and hydrogen bromide. J. Phys. Chem. 1984, 88, 3386–3388. [Google Scholar] [CrossRef]
- Yu, R.; Peethambaram, H.S.; Falta, R.W.; Verce, M.F.; Henderson, J.K.; Bagwell, C.E.; Brigmon, R.L.; Freedman, D.L. Kinetics of 1,2-Dichloroethane and 1,2-Dibromoethane Biodegradation in Anaerobic Enrichment Cultures. Appl. Environ. Microbiol. 2013, 79, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Kuntze, K.; Kozell, A.; Richnow, H.H.; Halicz, L.; Nijenhuis, I.; Gelman, F. Dual Carbon-Bromine Stable Isotope Analysis Allows Distinguishing Transformation Pathways of Ethylene Dibromide. Environ. Sci. Technol. 2016, 50, 9855–9863. [Google Scholar] [CrossRef] [PubMed]
- Poelarends, G.J.; Vlieg, J.; Marchesi, J.R.; Dos Santos, L.M.F.; Janssen, D.B. Degradation of 1,2-dibromoethane by Mycobacterium sp. strain GP1. J. Bacteriol. 1999, 181, 2050–2058. [Google Scholar] [PubMed]
- Belay, N.; Daniels, L. Production of ethane, ethylene, and acetylene from halogenated hydrocarbons by methanogenic bacteria. Appl. Environ. Microbiol. 1987, 53, 1604–1610. [Google Scholar]
- Hatzinger, P.B.; Begley, J.F.; Lippincott, D.R.; Bodou, A.; Forbes, R. In situ bioremediation of 1,2-dibromoethane (EDB) in groundwater to part-per-trillion concentrations using cometabolism. J. Contam. Hydrol. 2018, 218, 120–129. [Google Scholar] [CrossRef]
- Baek, K.; Wang, M.; McKeever, R.; Rieber, K.; Park, C.; Nuesslein, K. Biodegradation of low concentrations of 1,2-dibromoethane in groundwater is enhanced by phenol. Appl. Microbiol. Biotechnol. 2014, 98, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Henderson, J.K.; Freedman, D.L.; Falta, R.W.; Kuder, T.; Wilson, J.T. Anaerobic biodegradation of ethylene dibromide and 1,2-dichloroethane in the presence of fuel hydrocarbons. Environ. Sci. Technol. 2008, 42, 864–870. [Google Scholar] [CrossRef]
- Kao, C.-M.; Liao, H.-Y.; Chien, C.-C.; Tseng, Y.-K.; Tang, P.; Lin, C.-E.; Chen, S.-C. The change of microbial community from chlorinated solvent-contaminated groundwater after biostimulation using the metagenome analysis. J. Hazard. Mater. 2016, 302, 144–150. [Google Scholar] [CrossRef]
- Sheu, Y.T.; Lien, P.J.; Chen, K.F.; Ou, J.H.; Kao, C.M. Application of NZVI-contained emulsified substrate to bioremediate PCE-contaminated groundwater—A pilot-scale study. Chem. Eng. J. 2016, 304, 714–727. [Google Scholar] [CrossRef]
- Das, S.; Bora, S.S.; Yadav, R.N.S.; Barooah, M. A metagenomic approach to decipher the indigenous microbial communities of arsenic contaminated groundwater of Assam. Genom. Data 2017, 12, 89–96. [Google Scholar] [CrossRef]
- Bai, N.; Wang, S.; Abuduaini, R.; Zhang, M.; Zhu, X.; Zhao, Y. Rhamnolipid-aided biodegradation of carbendazim by Rhodococcus sp. D-1: Characteristics, products, and phytotoxicity. Sci. Total Environ. 2017, 590–591, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Xia, Y.; Zhang, T. Characterization of Thauera-dominated hydrogen-oxidizing autotrophic denitrifying microbial communities by using high-throughput sequencing. Bioresour. Technol. 2013, 128, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, P.A.; Crowley, J.R.; Sambandam, N.; Muegge, B.D.; Costello, E.K.; Hamady, M.; Knight, R.; Gordon, J.I. Regulation of myocardial ketone body metabolism by the gut microbiota during nutrient deprivation. Proc. Natl. Acad. Sci. USA 2009, 106, 11276–11281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Gloeckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Zemb, O.; Lee, M.; Low, A.; Manefield, M. Reactive iron barriers: A niche enabling microbial dehalorespiration of 1,2-dichloroethane. Appl. Microbiol. Biotechnol. 2010, 88, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.K.; Ding, C.; Yang, K.-L.; He, J. Complete Debromination of Tetra- and Penta-Brominated Diphenyl Ethers by a Coculture Consisting of Dehalococcoides and Desulfovibrio Species. Environ. Sci. Technol. 2011, 45, 8475–8482. [Google Scholar] [CrossRef]
- Grostern, A.; Edwards, E.A. A 1,1,1-trichloroethane-degrading anaerobic mixed microbial culture enhances biotransformation of mixtures of chlorinated ethenes and ethanes. Appl. Environ. Microbiol. 2006, 72, 7849–7856. [Google Scholar] [CrossRef]
- Zhang, H.; Ziv-El, M.; Rittmann, B.E.; Krajmalnik-Brown, R. Effect of Dechlorination and Sulfate Reduction on the Microbial Community Structure in Denitrifying Membrane-Biofilm Reactors. Environ. Sci. Technol. 2010, 44, 5159–5164. [Google Scholar] [CrossRef]
- Kim, E.-S.; Nomura, I.; Hasegawa, Y.; Takamizawa, K. Characterization of a newly isolated cis-1,2-dichloroethylene and aliphatic compound-degrading bacterium, Clostridium sp. strain KYT-1. Biotechnol. Bioprocess Eng. 2006, 11, 553–556. [Google Scholar] [CrossRef]
- Bajaj, A.; Kumar, A.; Yadav, S.; Kaur, G.; Bala, M.; Singh, N.K.; Kumar, R.M.; Manickam, N.; Mayilraj, S. Isolation and characterization of a novel Gram-negative bacterium Chromobacterium alkanivorans sp. nov., strain IITR-71(T) degrading halogenated alkanes. Int. J. Syst. Evol. Microbiol. 2016, 66, 5228–5235. [Google Scholar] [PubMed]
- Silva, Í.S.; dos Santos, E.C.; de Menezes, C.R.; de Faria, A.F.; Franciscon, E.; Grossman, M.; Durrant, L.R. Bioremediation of a polyaromatic hydrocarbon contaminated soil by native soil microbiota and bioaugmentation with isolated microbial consortia. Bioresour. Technol. 2009, 100, 4669–4675. [Google Scholar] [CrossRef] [PubMed]
- Antony, M.; Jayachandran, K. Regulation of Acyl Homoserine Lactone Synthesis in Pseudomonas putida JMQS1 Under Phenol Stress. Water Air Soil Pollut. 2016, 227, 228. [Google Scholar] [CrossRef]
- Cord-Ruwisch, R.; Ollivier, B. Interspecific hydrogen transfer during methanol degradation by Sporomusa acidovorans and hydrogenophilic anaerobes. Arch. Microbiol. 1986, 144, 163–165. [Google Scholar] [CrossRef]
- Kato, S.; Chino, K.; Kamimura, N.; Masai, E.; Yumoto, I.; Kamagata, Y. Methanogenic degradation of lignin-derived monoaromatic compounds by microbial enrichments from rice paddy field soil. Sci. Rep. 2015, 5, 14295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.I.; Kim, J.Y.; Yun, S.H.; Kim, J.H.; Leem, S.H.; Lee, C. Proteome analysis of Pseudomonas sp. K82 biodegradation pathways. Proteomics 2004, 4, 3610–3621. [Google Scholar] [CrossRef] [PubMed]
- Lors, C.; Ryngaert, A.; Perie, F.; Diels, L.; Damidot, D. Evolution of bacterial community during bioremediation of PAHs in a coal tar contaminated soil. Chemosphere 2010, 81, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Chen, M.; Li, F.; Liu, C.; Liao, C. Changes in the microbial community during repeated anaerobic microbial dechlorination of pentachlorophenol. Biodegradation 2017, 28, 219–230. [Google Scholar] [CrossRef]
- Van Hong Thi, P.; Jeong, S.-W.; Kim, J. Aquabacterium olei sp. nova, an oil-degrading bacterium isolated from oil-contaminated soil. Int. J. Syst. Evol. Microbiol. 2015, 65, 3597–3602. [Google Scholar]
- Jechalke, S.; Franchini, A.G.; Bastida, F.; Bombach, P.; Rosell, M.; Seifert, J.; von Bergen, M.; Vogt, C.; Richnow, H.H. Analysis of structure, function, and activity of a benzene-degrading microbial community. FEMS Microbiol. Ecol. 2013, 85, 14–26. [Google Scholar] [CrossRef] [Green Version]
- Nakatsu, C.H.; Hristova, K.; Hanada, S.; Meng, X.Y.; Hanson, J.R.; Scow, K.M.; Kamagata, Y. Methylibium petrolelphilum gen. nov., sp. nov., a novel methyl tert-butyl ether-degrading methylotroph of the Betaproteobacteria. Int. J. Syst. Evol. Microbiol. 2006, 56, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Pruden, A.; Suidan, M.T.; Venosa, A.D.; Wilson, G.J. Biodegradation of methyl tert-butyl ether under various substrate conditions. Environ. Sci. Technol. 2001, 35, 4235–4241. [Google Scholar] [CrossRef] [PubMed]
- Danko, A.S.; Leitao, P.O.; Verce, M.F.; Freedman, D.L. Efficacy of pentane, toluene, and benzene to support aerobic cometabolism of ethylene dibromide. New Biotechnol. 2012, 30, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Imron, M.F.; Kurniawan, S.B.; Soegianto, A. Characterization of mercury-reducing potential bacteria isolated from Keputih non-active sanitary landfill leachate, Surabaya, Indonesia under different saline conditions. J. Environ. Manag. 2019, 241, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Kosnar, Z.; Wiesnerova, L.; Castkova, T.; Kroulikova, S.; Boucek, J.; Mercl, F.; Tlustos, P. Bioremediation of polycyclic aromatic hydrocarbons (PAHs) present in biomass fly ash by co-composting and co-vermicomposting. J. Hazard. Mater. 2019, 369, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.S.; McKay, G. Sorption of dye from aqueous solution by peat. Chem. Eng. J. 1998, 70, 115–124. [Google Scholar] [CrossRef]
- Sag, Y.; Aktay, Y. Kinetic studies on sorption of Cr(VI) and Cu(II) ions by chitin, chitosan and Rhizopus arrhizus. Biochem. Eng. J. 2002, 12, 143–153. [Google Scholar] [CrossRef]
- Mukherjee, P.; Roy, P. Copper Enhanced Monooxygenase Activity and FT-IR Spectroscopic Characterisation of Biotransformation Products in Trichloroethylene Degrading Bacterium: Stenotrophomonas maltophilia PM102. Biomed. Res. Int. 2013. [Google Scholar] [CrossRef]
- Romano, S.; Schulz-Vogt, H.N.; Gonzalez, J.M.; Bondarev, V. Phosphate Limitation Induces Drastic Physiological Changes, Virulence- Related Gene Expression, and Secondary Metabolite Production in Pseudovibrio sp. Strain FO-BEG1. Appl. Environ. Microbiol. 2015, 81, 3518–3528. [Google Scholar] [CrossRef]
- Wen, L.-L.; Zhang, Y.; Chen, J.-X.; Zhang, Z.-X.; Yi, Y.-Y.; Tang, Y.; Rittmann, B.E.; Zhao, H.-P. The dechlorination of TCE by a perchlorate reducing consortium. Chem. Eng. J. 2017, 313, 1215–1221. [Google Scholar] [CrossRef]
- Scholz-Muramatsu, H.; Neumann, A.; Meßmer, M.; Moore, E.; Diekert, G. Isolation and characterization of Dehalospirillum multivorans gen. nov., sp. nov., a tetrachloroethene-utilizing, strictly anaerobic bacterium. Arch. Microbiol. 1995, 163, 48–56. [Google Scholar] [CrossRef]
- Sfetsas, C.C.; Milios, L.; Skopelitou, K.; Venieraki, A.; Todou, R.; Flemetakis, E.; Katinakis, P.; Labrou, N.E. Characterization of 1,2-dibromoethane-degrading haloalkane dehalogenase from Bradyrhizobium japonicum USDA110. Enzyme Microb. Technol. 2009, 45, 397–404. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, L.; Lee, Y.; Jahng, D. Novel bioevaporation process for the zero-discharge treatment of highly concentrated organic wastewater. Water Res. 2013, 47, 5678–5689. [Google Scholar] [CrossRef] [PubMed]
- Palau, J.; Yu, R.; Mortan, S.H.; Shouakar-Stash, O.; Rosell, M.; Freedman, D.L.; Sbarbati, C.; Fiorenza, S.; Aravena, R.; Marco-Urrea, E.; et al. Distinct Dual C-C1 Isotope Fractionation Patterns during Anaerobic Biodegradation of 1,2-Dichloroethane: Potential to Characterize Microbial Degradation in the Field. Environ. Sci. Technol. 2017, 51, 2685–2694. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Shang, H.T.; Wang, X.L.; Li, H.D.; Wang, J.L. Anaerobic degradation of tetrachloroethylene using different co-substrates as electron donors. Biomed. Environ. Sci. 2006, 19, 73–76. [Google Scholar] [PubMed]
- Baek, K.; McKeever, R.; Rieber, K.; Sheppard, D.; Park, C.; Ergas, S.J.; Nüsslein, K. Molecular approach to evaluate biostimulation of 1,2-dibromoethane in contaminated groundwater. Bioresour. Technol. 2012, 123, 207–213. [Google Scholar] [CrossRef]
- Im, J.; Semrau, J.D. Pollutant degradation by a Methylocystis strain SB2 grown on ethanol: Bioremediation via facultative methanotrophy. FEMS Microbiol. Lett. 2011, 318, 137–142. [Google Scholar] [CrossRef]
- Krajmalnik-Brown, R.; Holscher, T.; Thomson, I.N.; Saunders, F.M.; Ritalahti, K.M.; Loffler, F.E. Genetic identification of a putative vinyl chloride reductase in Dehalococcoides sp. strain BAV1. Appl. Environ. Microbiol. 2004, 70, 6347–6351. [Google Scholar] [CrossRef]
- Chow, W.L.; Cheng, D.; Wang, S.; He, J. Identification and transcriptional analysis of trans-DCE-producing reductive dehalogenases in Dehalococcoides species. ISME J. 2010, 4, 1020–1030. [Google Scholar] [CrossRef]
- Lamichhane, S.; Krishna, K.C.B.; Sarukkalige, R. Surfactant-enhanced remediation of polycyclic aromatic hydrocarbons: A review. J. Environ. Manag. 2017, 199, 46–61. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Q.; Jiang, J.; Wang, K.; Wei, L.; Ding, J.; Yu, H. Acceleration of organic removal and electricity generation from dewatered oily sludge in a bioelectrochemical system by rhamnolipid addition. Bioresour. Technol. 2017, 243, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Bouwer, E.J.; McCarty, P.L. Transformations of 1- and 2-carbon halogenated aliphatic organic compounds under methanogenic conditions. Appl. Environ. Microbiol. 1983, 45, 1286–1294. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Culture | Temperature (°C) | Co-Substrate | Initial EDB Conc. (mg/L) | Time for Corresponding Degraded Rate (Days) | Degraded Rate | Reference |
|---|---|---|---|---|---|---|---|
| Ancylobacter aquaticus AD20 | aerobic | 28 | No | 187 | - | - | [13] |
| Sulfurospirillum multivorans | anaerobic | 28 | No | 187 | - | - | [13] |
| Dehalococcoides sp. | anaerobic | 23 ± 1 | lactate | 2.2 | 20 | under MCL 2 | [12] |
| Mixed culture | aerobic | - | benzene | 6.6 | 4 | completely | [44] |
| Mixed culture | aerobic | - | pentane | 6.6 | 8 | 0 | [44] |
| Mixed culture | aerobic | - | toluene | 6.6 | 8 | 0 | [44] |
| Indigenous microorganisms | aerobic | 14 ± 1 | ethane | 0.6 | 0.16 | 75% | [7] |
| Indigenous microorganisms | aerobic | 24 ± 1 | ethane | 0.06 | 65 | completely | [7] |
| Indigenous microorganisms | aerobic | 14 ± 1 | methane + DAP 1 | 0.06 | 56 | 60% | [7] |
| Indigenous microorganisms | aerobic | 24 ± 1 | phenol | 0.06 | 56 | 20% | [7] |
| Indigenous microorganisms | aerobic | 24 ± 1 | propane | 0.06 | 65 | completely | [7] |
| Indigenous microorganisms | aerobic | 12 ± 2 | phenol | 0.01 | 200 | 80% | [17] |
| Indigenous microorganisms | aerobic | 12 ± 2 | methane | 0.08 | ≥230 | completely | [2] |
| Indigenous microorganisms | anaerobic | 12 ± 2 | No | 0.05 | 282 | 85% | [2] |
| Indigenous microorganisms | anaerobic | 12 ± 2 | lactate | 0.09 | 282 | 88% | [2] |
| Microbial consortium | From aerobic to anaerobic (initial DO: ~2 mg/L) | 15 | no | 0.1–1.0 | 6–20 | >61% | present study |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Yang, M.; Song, X.; Tang, S.; Yu, L. Aerobic and Anaerobic Biodegradation of 1,2-Dibromoethane by a Microbial Consortium under Simulated Groundwater Conditions. Int. J. Environ. Res. Public Health 2019, 16, 3775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16193775
Wang Q, Yang M, Song X, Tang S, Yu L. Aerobic and Anaerobic Biodegradation of 1,2-Dibromoethane by a Microbial Consortium under Simulated Groundwater Conditions. International Journal of Environmental Research and Public Health. 2019; 16(19):3775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16193775
Chicago/Turabian StyleWang, Qing, Miaoyan Yang, Xin Song, Shiyue Tang, and Lei Yu. 2019. "Aerobic and Anaerobic Biodegradation of 1,2-Dibromoethane by a Microbial Consortium under Simulated Groundwater Conditions" International Journal of Environmental Research and Public Health 16, no. 19: 3775. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16193775