Anti-Biofouling Performance of an Immobilized Indigenous Quorum Quenching Bacterium Bacillus cereus HG10 and Its Influence on the Microbial Community in a Bioreactor


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Synthetic Wastewater
2.3. Bead Preparation
2.4. Operation of the Batch Bioreactor
2.5. Analytical Methods
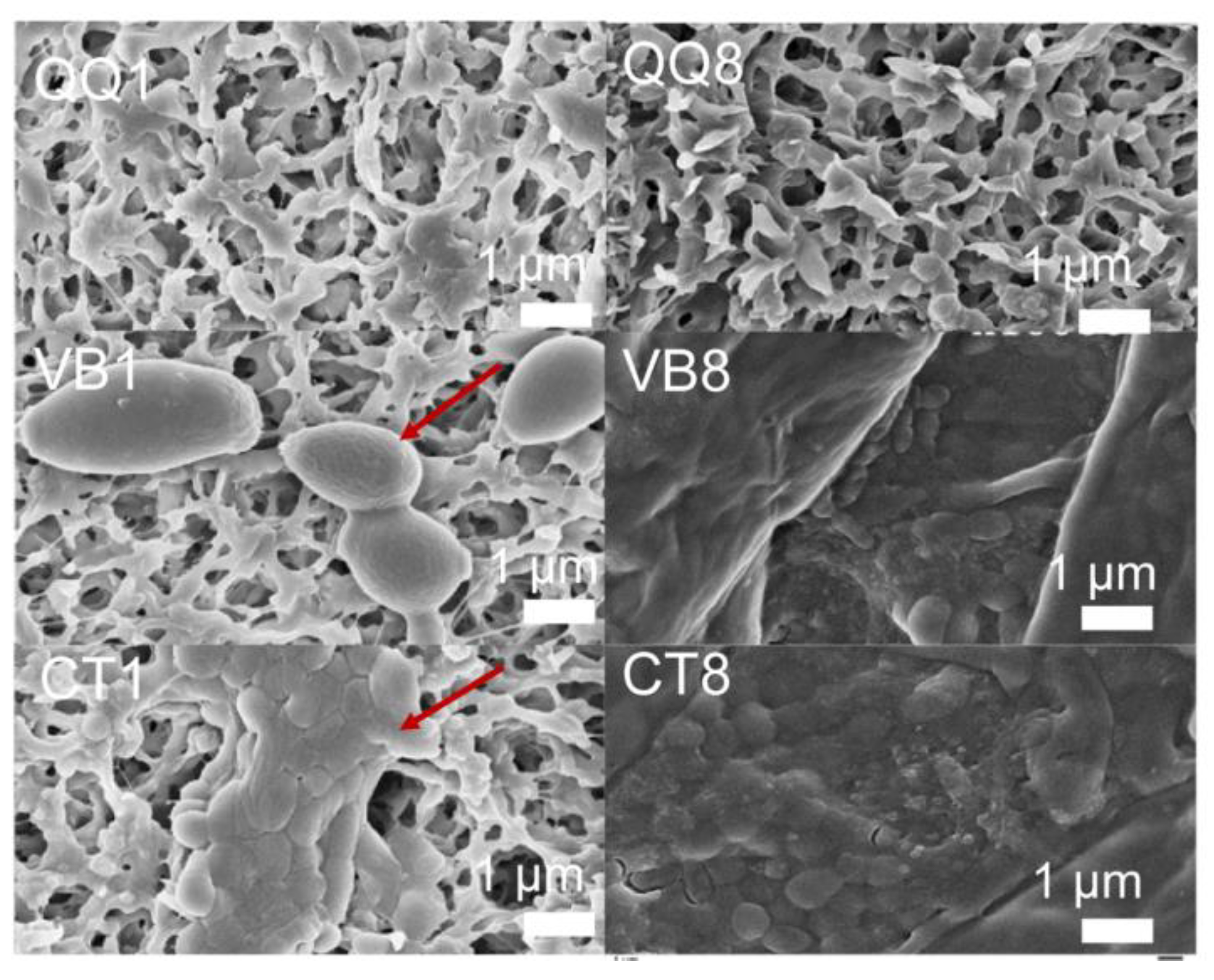
2.6. SEM Observation of Biofouling
2.7. Sample and 16S Library Preparation
2.8. Computational Analysis
3. Results and Discussion
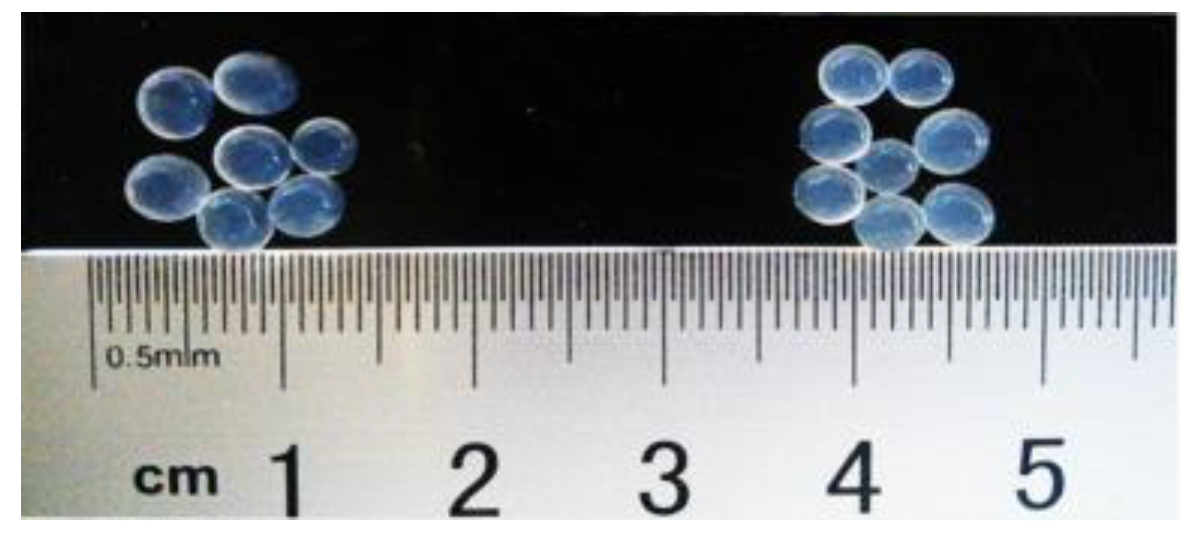
3.1. Characterization of Beads
3.2. Impact of QQ Beads for Alleviating Biofouling on Filter Membranes
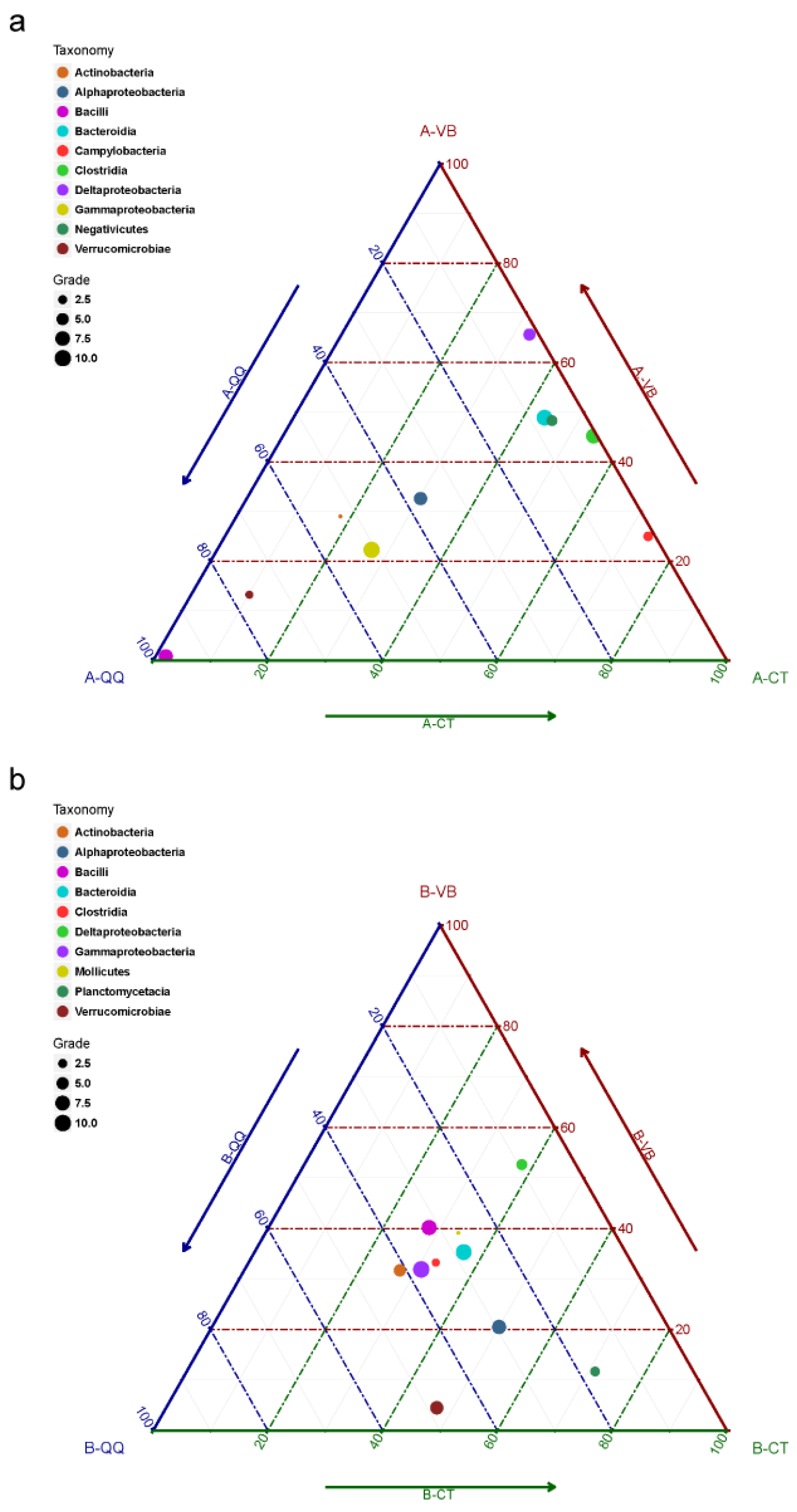
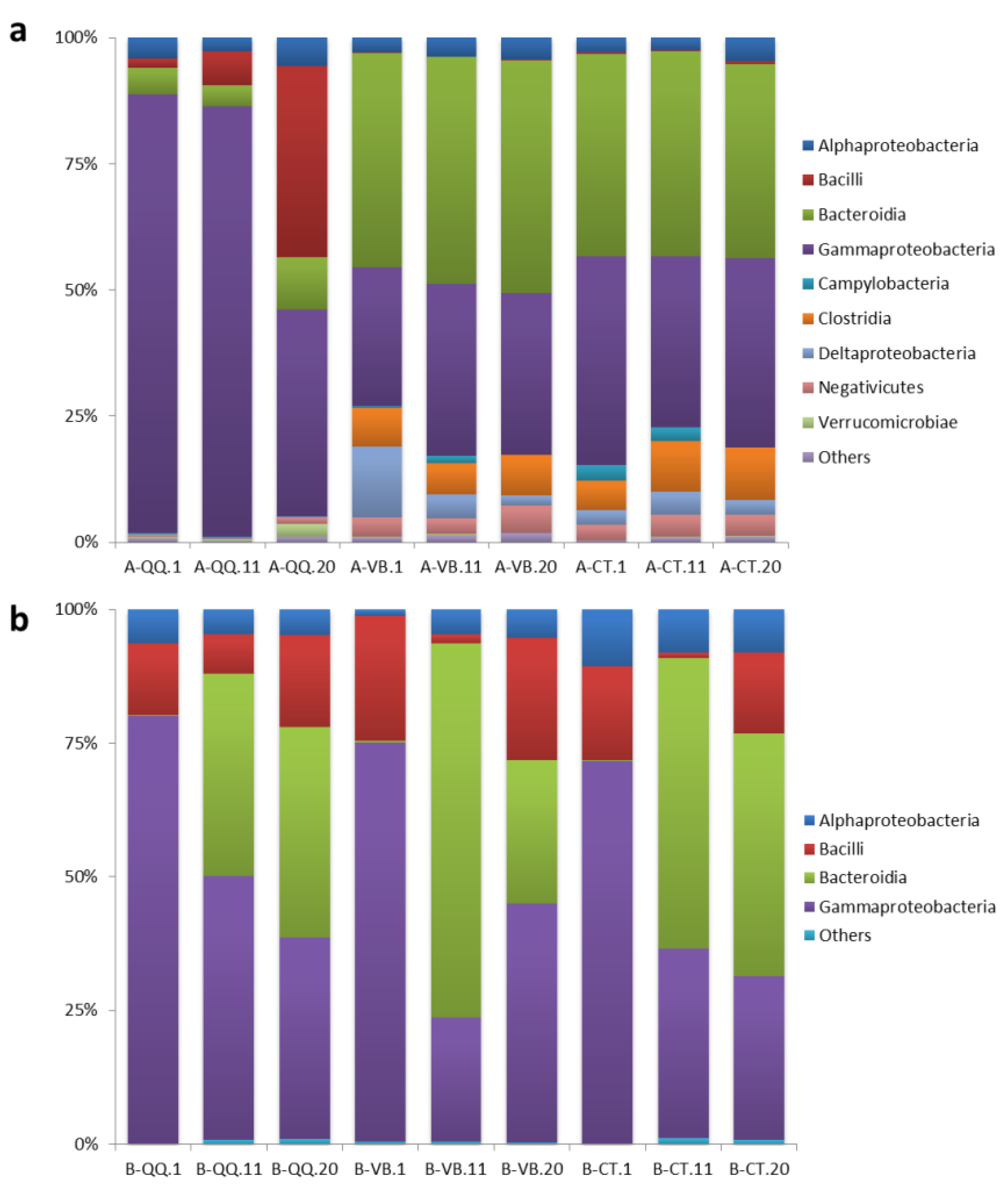
3.3. Sequencing of Activated Sludge and Biofilm Samples
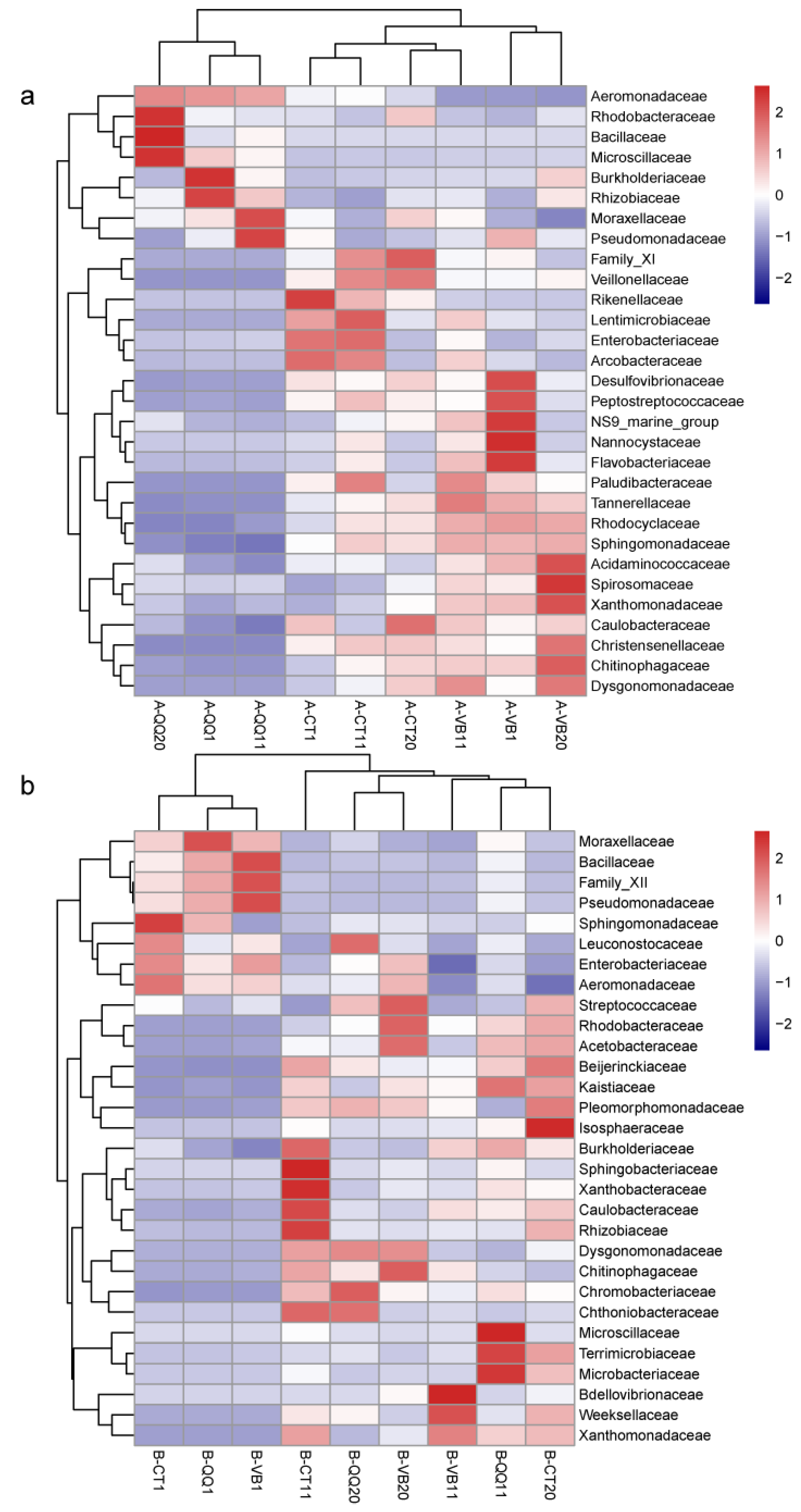
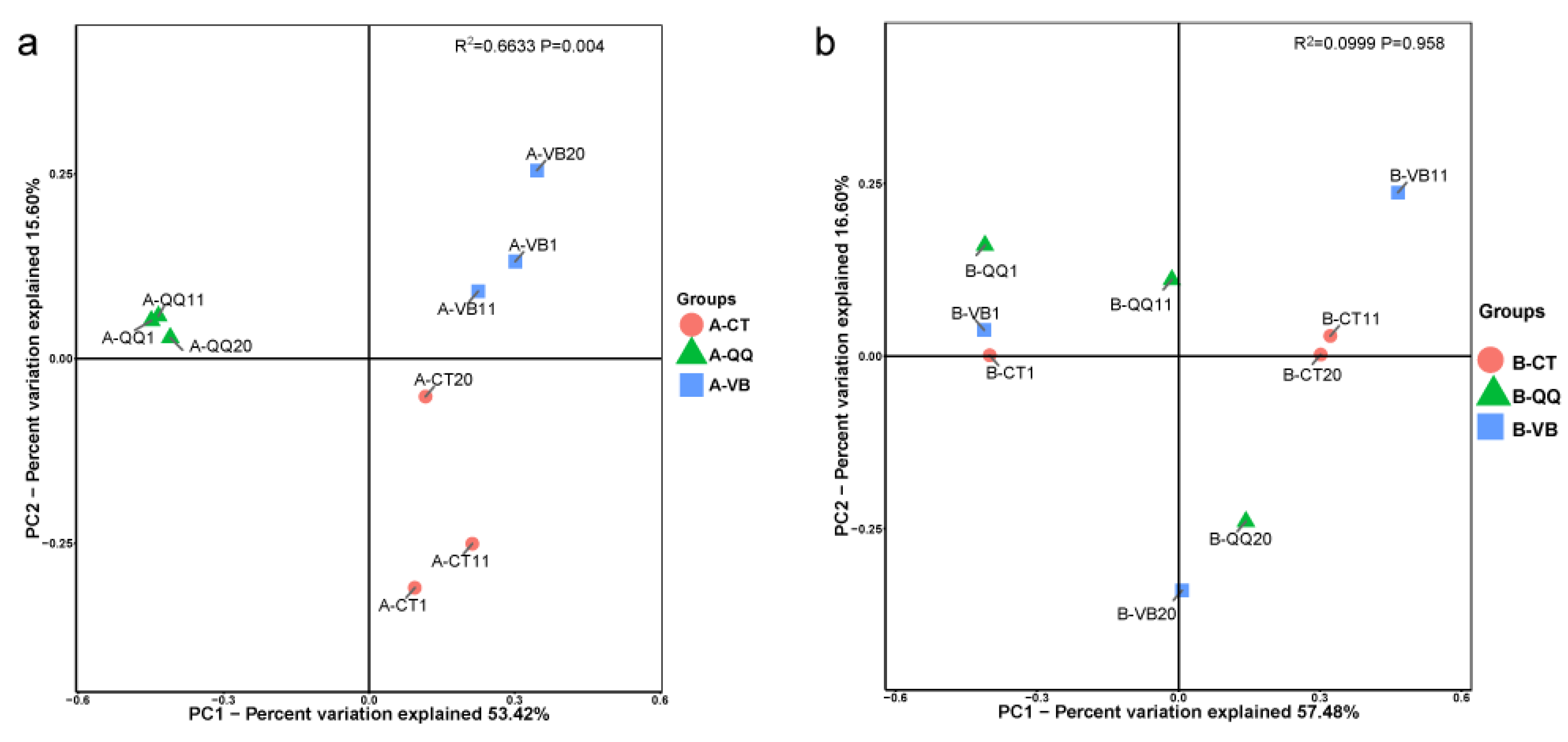
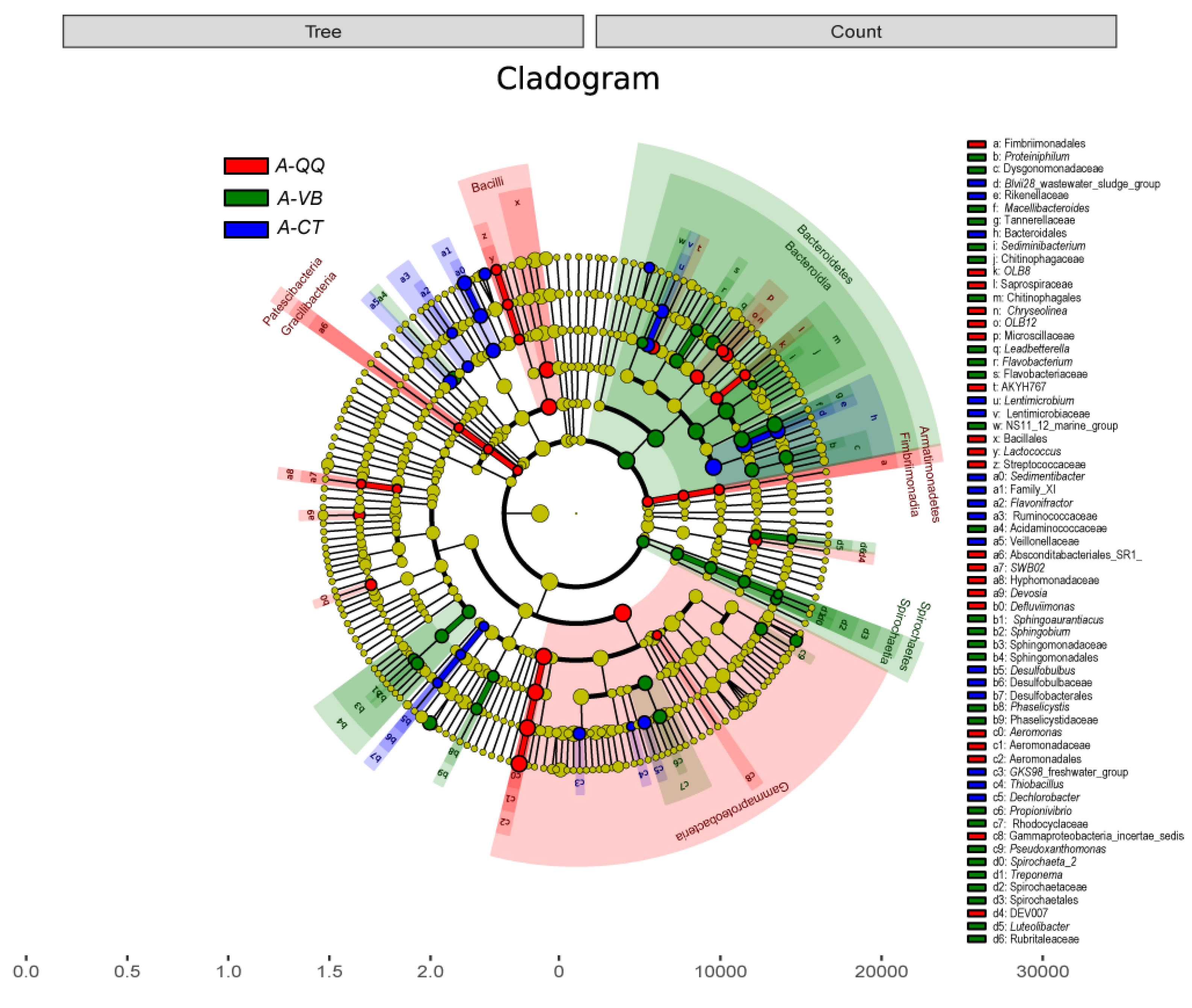
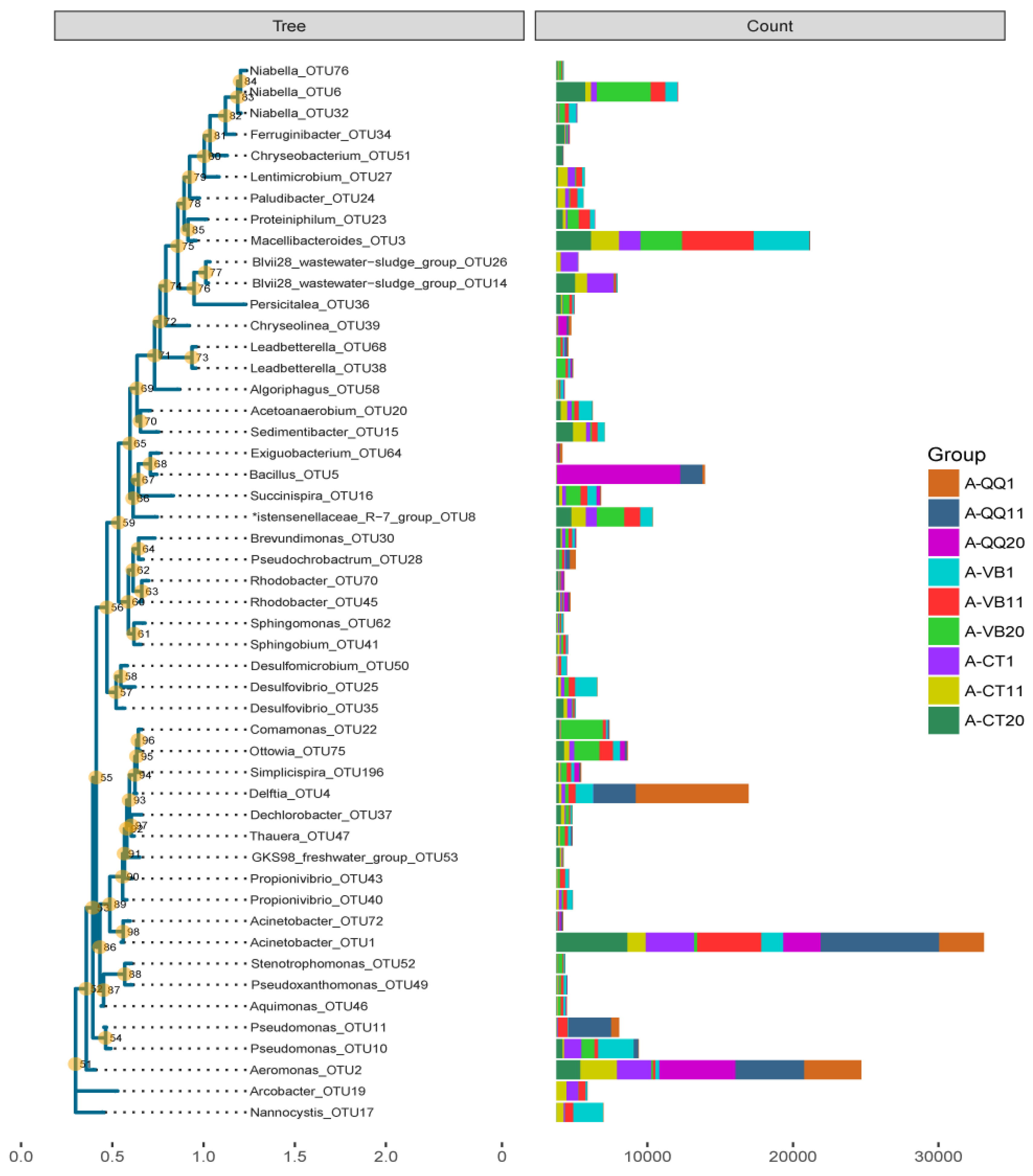
3.4. Effect of QQ Beads on the Microbial Community Structure of Activated Sludge and Biofilm
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Krzeminski, P.; Leverette, L.; Malamis, S.; Katsou, E. Membrane bioreactors—A review on recent developments in energy reduction, fouling control, novel configurations, LCA and market prospects. J. Membr. Sci. 2017, 527, 207–227. [Google Scholar] [CrossRef]
- Meng, F.; Zhang, S.; Oh, Y.; Zhou, Z.; Shin, H.-S.; Chae, S.-R. Fouling in membrane bioreactors: An updated review. Water Res. 2017, 114, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Z.; Chen, M.; Ma, J.; Chen, S.; Wu, Z. Membrane biofouling control using polyvinylidene fluoride membrane blended with quaternary ammonium compound assembled on carbon material. J. Membr. Sci. 2017, 539, 229–237. [Google Scholar] [CrossRef]
- Amouamouha, M.; Badalians Gholikandi, G. Characterization and Antibiofouling Performance Investigation of Hydrophobic Silver Nanocomposite Membranes: A Comparative Study. Membranes 2017, 7, 64. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, X.; Wang, Z.; Wang, L.; Wu, Z. QAC modified PVDF membranes: Antibiofouling performance, mechanisms, and effects on microbial communities in an MBR treating municipal wastewater. Water Res. 2017, 120, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Guo, W.; Ngo, H.H.; Zhang, H.; Wang, J.; Li, J.; Xia, S.; Wu, Y. Biofouling and control approaches in membrane bioreactors. Bioresour. Technol. 2016, 221, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, J.; Hrenovic, J.; Goic-Barisic, I.; Tomic, M.; Djonlagic, J.; Rajic, N. Synergistic anti-biofouling effect of Ag-exchanged zeolite and D-Tyrosine on PVC composite against the clinical isolate of Acinetobacter baumannii. Biofouling 2014, 30, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, N.; Dietrich, N.; Lafforgue, C.; Lee, C.H.; Guigui, C. Process-Oriented Review of Bacterial Quorum Quenching for Membrane Biofouling Mitigation in Membrane Bioreactors (MBRs). Membranes 2016, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- Cheong, W.S.; Kim, S.R.; Oh, H.S.; Lee, S.H.; Yeon, K.M.; Lee, C.H.; Lee, J.K. Design of quorum quenching microbial vessel to enhance cell viability for biofouling control in membrane bioreactor. J. Microbiol. Biotechnol. 2014, 24, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R.; Oh, H.S.; Jo, S.J.; Yeon, K.M.; Lee, C.H.; Lim, D.J.; Lee, C.H.; Lee, J.K. Biofouling control with bead-entrapped quorum quenching bacteria in membrane bioreactors: physical and biological effects. Environ. Sci. Technol. 2013, 47, 836–842. [Google Scholar] [CrossRef]
- Nguyen, T.; Roddick, F.A.; Fan, L. Biofouling of Water Treatment Membranes: A Review of the Underlying Causes, Monitoring Techniques and Control Measures. Membranes 2012, 2, 804–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watnick, P.; Kolter, R. Biofilm, City of Microbes. J. Bacteriol. 2000, 182, 2675–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan-Sagastume, F.; Boon, N.; Dobbelaere, S.; Defoirdt, T.; Verstraete, W. Production of acylated homoserine lactones by Aeromonas and Pseudomonas strains isolated from municipal activated sludge. Can. J. Microbiol. 2005, 51, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Chong, G.; Kimyon, O.; Rice, S.A.; Kjelleberg, S.; Manefield, M. The presence and role of bacterial quorum sensing in activated sludge. Microbiol. Biotechnol. 2012, 5, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Yeon, K.M.; Cheong, W.S.; Oh, H.S.; Lee, W.N.; Hwang, B.K.; Lee, C.H.; Beyenal, H.; Lewandowski, Z. Quorum sensing: a new biofouling control paradigm in a membrane bioreactor for advanced wastewater treatment. Environ. Sci. Technol. 2009, 43, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.F.; Sakinah, M.; Singh, L.; Zularisam, A.W. Targeting N-acyl-homoserine-lactones to mitigate membrane biofouling based on quorum sensing using a biofouling reducer. J. Biotechnol. 2012, 161, 190–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Lee, S.; Lee, S.H.; Kim, S.R.; Oh, H.S.; Park, P.K.; Choo, K.H.; Kim, Y.W.; Lee, J.K.; Lee, C.H. Fungal Quorum Quenching: A Paradigm Shift for Energy Savings in Membrane Bioreactor (MBR) for Wastewater Treatment. Environ. Sci. Technol. 2016, 50, 10914–10922. [Google Scholar] [CrossRef]
- Ochiai, S.; Morohoshi, T.; Kurabeishi, A.; Shinozaki, M.; Fujita, H.; Sawada, I.; Ikeda, T. Production and degradation of N-acylhomoserine lactone quorum sensing signal molecules in bacteria isolated from activated sludge. Biosci. Biotechnol. Biochem. 2013, 77, 2436–2440. [Google Scholar] [CrossRef]
- ZHAO, C.; WANG, W.Z.; XU, Q.Y. Isolation of Quorum Quenching Bacteria and Their Function for Controlling Membrane Biofouling. Environ. Sci. 2016, 37, 4720–4726. [Google Scholar] [CrossRef]
- APHA. APHA Standard Methods for Water and Wastewater Examination; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Judd, S. The MBR Book, 2nd ed.; Butterworth-Heinemann: Oxford, UK, 2011; pp. 497–519. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinformatics 2011, 12, 385. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2--approximately maximum-likelihood trees for large alignments. PloS ONE, 2010; 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevenes, M.H.H.; Wagner, H.H. Vegan: Community Ecology Package. R version 2. 2013. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Martínez, M.S.; Uyttendaele, M.; Rajkovic, A.; Nadal, P.; Debevere, J. Degradation of N-Acyl-l-Homoserine Lactones by Bacillus cereus in Culture Media and Pork Extract. Appl. Environ. Microbiol. 2007, 73, 2329–2332. [Google Scholar] [CrossRef] [PubMed]
- Yavuztürk Gül, B.; Imer, D.Y.; Park, P.-K.; Koyuncu, I. Evaluation of a novel anti-biofouling microorganism (Bacillus sp. T5) for control of membrane biofouling and its effect on bacterial community structure in membrane bioreactors. Water Sci. Technol. 2017. [Google Scholar] [CrossRef]
- Zamani, M.; Behboudi, K.; Ahmadzadeh, M. Quorum quenching by Bacillus cereus U92: a double-edged sword in biological control of plant diseases. Biocontrol Sci. Technol. 2013, 23, 555–573. [Google Scholar] [CrossRef]
- Jo, S.J.; Kwon, H.; Jeong, S.Y.; Lee, S.H.; Oh, H.S.; Yi, T.; Lee, C.H.; Kim, T.G. Effects of Quorum Quenching on the Microbial Community of Biofilm in an Anoxic/Oxic MBR for Wastewater Treatment. J. Microbiol. Biotechn. 2016, 26, 1593–1604. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Mishra, A.; Jha, B. Anti-quorum Sensing and Anti-biofilm Activity of Delftia tsuruhatensis Extract by Attenuating the Quorum Sensing-Controlled Virulence Factor Production in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2017, 7, 337. [Google Scholar] [CrossRef] [PubMed]
- Maisuria, V.B.; Nerurkar, A.S. Interference of Quorum Sensing by Delftia sp. VM4 Depends on the Activity of a Novel N-Acylhomoserine Lactone-Acylase. PloS ONE 2015, 10, e0138034. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Dessaux, Y.; Oger, P. Quorum sensing and quorum quenching: the yin and yang of bacterial communication. Chem. Bio. Chem. 2009, 10, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, E.; Redfern, L.; Cooper, E.M.; Stapleton, H.M.; Gunsch, C.K. Acetate promotes microbial reductive debromination of tetrabromobisphenol A during the startup phase of anaerobic wastewater sludge bioreactors. Sci. Total Environ. 2019, 656, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, S.; Lv, X.; Qiu, Z.; Zhang, Z.; Yan, L. Dissolved organic matter release in overlying water and bacterial community shifts in biofilm during the decomposition of Myriophyllum verticillatum. Sci. Total Environ. 2018, 633, 929–937. [Google Scholar] [CrossRef]
- Dasgupta, D.; Ghosh, R.; Sengupta, T.K. Biofilm-mediated enhanced crude oil degradation by newly isolated pseudomonas species. ISRN Biotechnol. 2013, 2013, 250749. [Google Scholar] [CrossRef]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, F.; Zhao, C.; Lee, C.H.; Wang, W.; Xu, Q. Anti-Biofouling Performance of an Immobilized Indigenous Quorum Quenching Bacterium Bacillus cereus HG10 and Its Influence on the Microbial Community in a Bioreactor. Int. J. Environ. Res. Public Health 2019, 16, 3777. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16193777
Xu F, Zhao C, Lee CH, Wang W, Xu Q. Anti-Biofouling Performance of an Immobilized Indigenous Quorum Quenching Bacterium Bacillus cereus HG10 and Its Influence on the Microbial Community in a Bioreactor. International Journal of Environmental Research and Public Health. 2019; 16(19):3777. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16193777
Chicago/Turabian StyleXu, Fangfang, Chang Zhao, Chuang Hak Lee, Wenzhao Wang, and Qiyong Xu. 2019. "Anti-Biofouling Performance of an Immobilized Indigenous Quorum Quenching Bacterium Bacillus cereus HG10 and Its Influence on the Microbial Community in a Bioreactor" International Journal of Environmental Research and Public Health 16, no. 19: 3777. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16193777