Can Low-Level Exposure to Radiofrequency Fields Effect Cognitive Behaviour in Laboratory Animals? A Systematic Review of the Literature Related to Spatial Learning and Place Memory

Abstract
:1. Introduction
2. Search Strategy and Inclusion Criteria
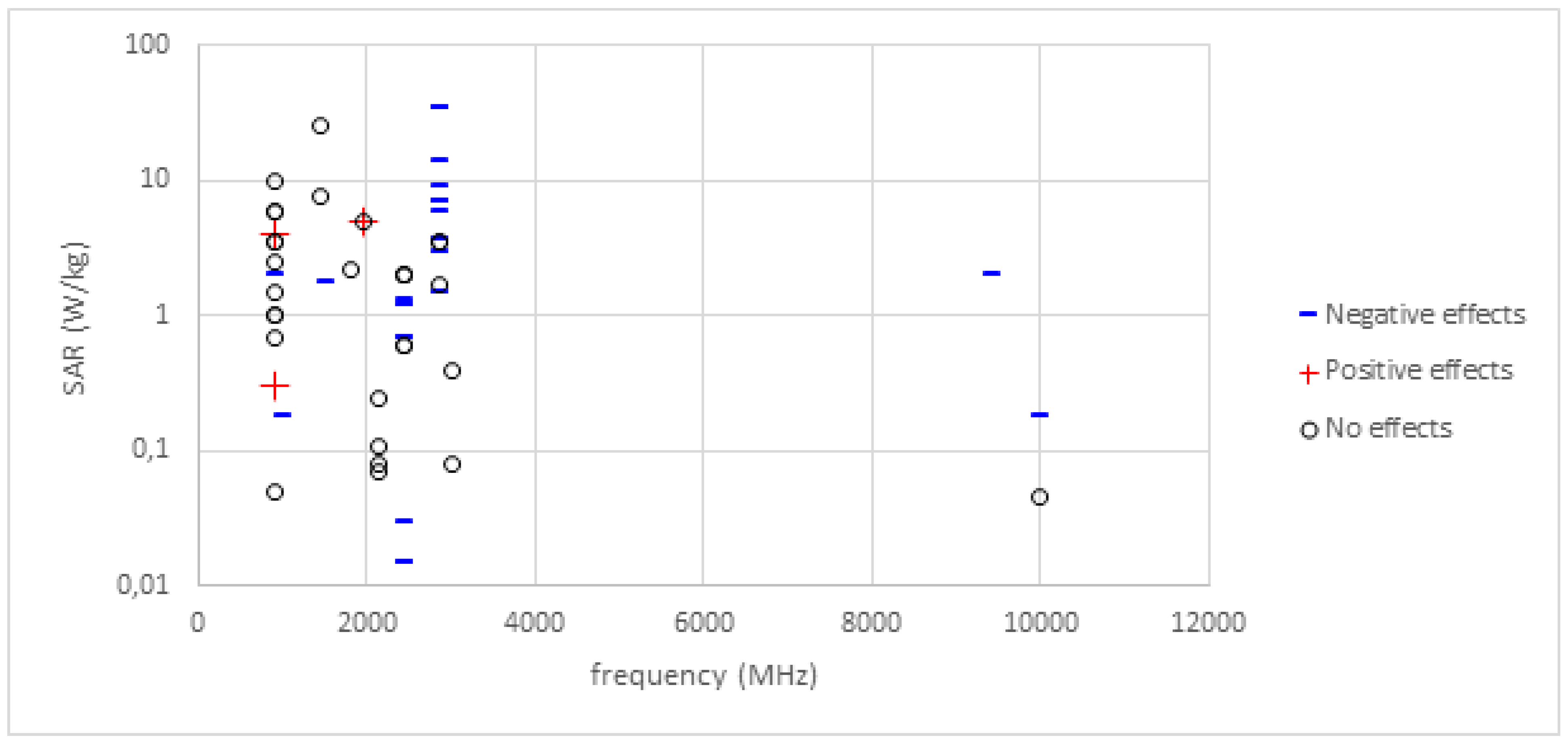
3. Results
3.1. Narrative Review of Studies
3.1.1. Radial Arm Maze Studies: Exposure of Juvenile or Adult Rats
3.1.2. Radial Arm Maze Studies: Exposure of Juvenile or Adult Mice
3.1.3. Morris Water Maze Studies: Exposure of Juvenile or Adult Rats
3.1.4. Morris Water Maze Studies: Early Life Exposures of Rats
3.1.5. Morris Water Maze Studies: Exposure of Juvenile or Adult Mice
3.1.6. Morris Water Maze Studies: Early Life Exposures of Mice
3.1.7. T-Maze Studies: Exposure of Juvenile or Adult Rats
3.1.8. Studies Using Juvenile or Adult Transgenic Animals
3.1.9. Navigation in a Migratory Bird
4. Discussion
5. Conclusions
- Possible confounding resulting from immobilization stress caused by restraining the animals during exposure should be avoided, so exposure systems, like reverberation chambers which can produce well-controlled and repeatable SARs without the need for restraint, are recommended.
- A single whole-body SAR value should also be avoided, and experiments should use a range of whole-body SARs from 0 to 5 W/kg or more (depending on species) to provide some indication of the dose-response relationship. If possible, an appropriate positive control group should also be included. However, exposures causing overt heating of the animal, where body core temperatures are raised by 1 °C or more should be avoided in order to prevent any confounding from thermal effects.
- Of particular importance with studies using RF fields is the necessity for detailed computational dosimetry in heterogeneous, anatomically realistic animal models to determine both the average absorbed energy for the whole animal, and the spatial distribution of the absorbed energies at the organ and tissue level. Unfortunately, not all recent papers (see studies listed in Appendix A) provide sufficient information to allow a reasonable estimate of the SAR to be made.
- Given the ubiquity of the Morris water maze in many laboratories investigating neurobehavioral toxicology and teratology, the use of this maze for these studies seems most appropriate. The task is suitable for animals of all ages, including weaning mice [61] and is amenable to using the many commercial systems that allow automatic recording and analysis of the animals’ behaviour in the maze, so helping to eliminate a potential source of subjective error. The data in the computer files can also be readily re-analysed if necessary.
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
| Study | Reason for Non-Inclusion |
|---|---|
| Kumar, R.S.; Sareesh, N.N.; Nayak, S.; Mailankot, M. Hypoactivity of Wistar rats exposed to mobile phone on elevated plus maze. Indian J Physiol Pharmacol 2009, 53(3), 283–286. | No information on exposure level, no sham-exposed control group. |
| Narayanan, S.N.; Kumar, R.S.; Potu, B.K.; Nayak, S.; Mailankot, M. Spatial memory performance of Wistar rats exposed to mobile phone. Clinics (Sao Paulo) 2009, 64(3), 231–234, doi:10.1590/S1807-59322009000300014. | No information on exposure level. |
| Fragopoulou, A.F.; Miltiadous, P.; Stamatakis, A.; Stylianopoulou, F.; Koussoulakos, S.L.; Margaritis, L.H. Whole body exposure with GSM 900MHz affects spatial memory in mice. Pathophysiology 2010, 17(3), 179–187, doi:10.1016/j.pathophys.2009.11.002. | Exposure level not clear. |
| Arendash, G.W.; Sanchez-Ramos, J.; Mori, T.; Mamcarz, M.; Lin, X.; Runfeldt, M.; Wang, L.; Zhang, G.; Sava, V.; Tan, J.; Cao, C. Electromagnetic field treatment protects against and reverses cognitive impairment in Alzheimer’s disease mice. J Alzheimers Dis 2010, 19(1), 191–210, doi:10.3233/JAD-2010-1228. | Assessment of SAR levels not clear and no information about other relevant exposure quantities provided. |
| Zhao, L.; Peng, R.Y.; Wang, S.M.; Wang, L.F.; Gao, Y.B.; Dong, J.; Li, X.; Su, Z.T.; et al. Relationship between cognition function and hippocampus structure after long-term microwave exposure. Biomed Environ Sci 2012, 25(2), 182–188, doi:10.3967/0895-3988.2012.02.009. | Type of field used and timing of assay relative to exposure not specified. |
| Hao, D.; Yang, L.; Chen, S.; Tian, Y.; Wu, S. 916 MHz electromagnetic field exposure affects rat behavior and hippocampal neuronal discharge. Neural Regen Res 2012, 7(19),1488–1492, doi:10.3969/j.issn.1673-5374.2012.19.007. | Power density measured in centre of cage, but large variation likely, therefore inadequate dosimetry. |
| Hao, D.; Yang, L.; Chen, S.; Tong, J.; Tian, Y.; Su, B.; Wu, S.; Zeng, Y. Effects of long-term electromagnetic field exposure on spatial learning and memory in rats. Neurol Sci 2013, 34(2), 157–164, doi:10.1007/s10072-012-0970-8. | Power density measured in centre of cage, but large variation likely, therefore inadequate exposure description. |
| Banaceur, S.; Banasr, S.; Sakly, M.; Abdelmelek, H. Whole body exposure to 2.4 GHz WIFI signals: effects on cognitive impairment in adult triple transgenic mouse models of Alzheimer’s disease (3xTg-AD). Behav Brain Res 2013, 240, 197–201, doi:10.1016/j.bbr.2012.11.021. | SAR values provided, but no information about dosimetry methods, and no other information about exposure level. |
| İkinci, A.; Odacı, E.; Yıldırım, M.; Kaya, H.; Akça, M.; Hancı, H.; Aslan, A.; Sönmez, O.F.; Baş, O. The effects of prenatal exposure to a 900 megahertz electromagnetic field on hippocampus morphology and learning behavior in rat pups. Neuroquantology 2013, 11(4), 582–590, doi:10.14704/nq.2013.11.4.699. | No dosimetry. |
| Razavinasab, M.; Moazzami, K.; Shabani, M. Maternal mobile phone exposure alters intrinsic electrophysiological properties of CA1 pyramidal neurons in rat offspring. Toxicol Ind Health 2016, 32(6), 968–979, doi:10.1177/0748233714525497. | Assessment of SAR levels not clear and no information about other relevant exposure quantities provided. |
| Saikhedkar, N.; Bhatnagar, M.; Jain, A.; Sukhwal, P.; Sharma, C.; Jaiswal, N. Effects of mobile phone radiation (900 MHz radiofrequency) on structure and functions of rat brain. Neurol Res 2014, 36(12), 1072–1079, doi:10.1179/1743132814Y.0000000392. | Incorrect dosimetry (SAR calculated for human situation), and no other information about exposure level. |
| Maaroufi, K.; Had-Aissouni, L.; Melon, C.; Sakly, M.; Abdelmelek, H.; Poucet, B.; Save, E. Spatial learning, monoamines and oxidative stress in rats exposed to 900 MHz electromagnetic field in combination with iron overload. Behav Brain Res 2014, 25, 80–89, doi:10.1016/j.bbr.2013.10.016. | SAR calculated using external E field, which is not provided, and no other information about exposure level. |
| Narayanan, S.N.; Kumar, R.S.; Karun, K.M.; Nayak, S.B.; Bhat, P.G. Possible cause for altered spatial cognition of prepubescent rats exposed to chronic radiofrequency electromagnetic radiation. Metab Brain Dis 2015, 30(5), 1193–1206, doi:10.1007/s11011-015-9689-6. | Mobile phone in cage, no dosimetry. |
| Nirwane, A.; Sridhar, V.; Majumdar, A. Neurobehavioural changes and brain oxidative stress induced by acute exposure to GSM900 mobile phone radiations in zebrafish (Danio rerio). Toxicol Res 2016, 32(2), 123–132, doi:10.5487/TR.2016.32.2.123. | Mobile phone above fish tank, no dosimetry. |
| Wang, L.F.; Tian, D.W.; Li, H.J.; Gao, Y.B.; Wang, C.Z.; Zhao, L.; Zuo, H.Y.; Dong, J.; Qiao, S.M.; Zou, Y.; Xiong, L.; Zhou, H.M.; Yang, Y.F.; Peng, R.Y.; Hu, X.J. Identification of a novel rat NR2B subunit gene promoter region variant and its association with microwave-induced neuron impairment. Mol Neurobiol 2016, 53(4), 2100–2111, doi:10.1007/s12035-015-9169-3. | Frequency used not given. Incomplete dosimetry |
| Othman, H.; Ammari, M.; Sakly, M.; Abdelmelek, H. Effects of repeated restraint stress and WiFi signal exposure on behavior and oxidative stress in rats. Metab Brain Dis 2017, 32(5), 1459–1469, doi:10.1007/s11011-017-0016-2. | No dosimetry. |
| Varghese, R.; Majumdar, A.; Kumar, G.; Shukla, A. Rats exposed to 2.45 GHz of non-ionizing radiation exhibit behavioral changes with increased brain expression of apoptotic caspase 3. Pathophysiology 2018, 25(1), 19–30, doi:10.1016/j.pathophys.2017.11.001. | Incomplete dosimetry. |
References
- SCENIHR. Potential Health Effects of Exposure to Electromagnetic Fields (EMF). Scientific Committee on Emerging and Newly Identified Health Risks. 27 January 2015. Available online: https://ec.europa.eu/health/sites/health/files/scientific_committees/emerging/docs/scenihr_o_041.pdf (accessed on 24 January 2019).
- WHO. Electromagnetic Fields (300 Hz to 300 GHz); World Health Organization: Geneva, Switzerland, 1993. [Google Scholar]
- AGNIR. Health effects from radiofrequency electromagnetic fields. In Report of an Independent Advisory Group on Non-Ionising Radiation; Doc NRPB: Chilton, UK, 2003; Volume 14, pp. 1–177. [Google Scholar]
- AGNIR. Health effects from radiofrequency electromagnetic fields. In Report of the Independent Advisory Group on Non-Ionising Radiation; Doc HPA: London, UK, 2012; Volume 20, pp. 1–333. [Google Scholar]
- ICNIRP. Exposure to High Frequency Electromagnetic Fields, Biological Effects and Health Consequences (100 kHz–300 GHz); International Commission on Non-Ionizing Radiation Protection: Munich, Germany, 2009; pp. 1–378. [Google Scholar]
- SCENIHR. Possible Effects of Electromagnetic Fields (EMF) on Human Health. Scientific Committee on Emerging and Newly Identified Health Risks. 21 March 2007. Available online: https://ec.europa.eu/health/ph_risk/committees/04_scenihr/docs/scenihr_o_007.pdf (accessed on 24 January 2019).
- Chou, C.K.; Guy, A.W.; McDougall, J.A.; Lai, H. Specific absorption rate in rats exposed to 2450-MHz microwaves under seven exposure conditions. Bioelectromagnetics 1985, 6, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.; Horita, A.; Guy, A.W. Microwave irradiation affects radial-arm maze performance in the rat. Bioelectromagnetics 1994, 15, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Lai, H. Acute exposure to pulsed 2450-MHz microwaves affects water-maze performance of rats. Bioelectromagnetics 2000, 21, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Lai, H. Interaction of microwaves and a temporally incoherent magnetic field on spatial learning in the rat. Physiol. Behav. 2004, 82, 785–789. [Google Scholar] [CrossRef]
- Li, M.; Wang, Y.; Zhang, Y.; Zhou, Z.; Yu, Z. Elevation of plasma corticosterone levels and hippocampal glucocorticoid receptor translocation in rats: A potential mechanism for cognition impairment following chronic low-power-density microwave exposure. J. Radiat. Res. 2008, 49, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, O.P. (Ed.) Biological Effects and Medical Applications of Electromagnetic Energy; Prentice Hall: Englewood Cliffs, NJ, USA, 1990. [Google Scholar]
- Chaturvedi, C.M.; Singh, V.P.; Singh, P.; Basu, P.; Singaravel, M.; Shukla, R.K.; Dhawan, A.; Pati, A.K.; Gangwar, R.K.; Singh, S.P. 2.45 GHz (CW) microwave irradiation alters circadian organization, spatial memory, DNA structure in the brain cells and blood cell counts of male mice, Mus musculus. Prog. Electromagn. Res. B 2011, 29, 23–42. [Google Scholar] [CrossRef]
- Lu, Y.; Xu, S.; He, M.; Chen, C.; Zhang, L.; Liu, C.; Chu, F.; Yu, Z.; Zhou, Z.; Zhong, M. Glucose administration attenuates spatial memory deficits induced by chronic low-power-density microwave exposure. Physiol. Behav. 2012, 106, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, P.S.; Banerjee, B.D.; Abegaonkar, M.P.; Megha, K.; Ahmed, R.S.; Tripathi, A.K.; Mediratta, P.K. Effect of low level microwave radiation exposure on cognitive function and oxidative stress in rats. Indian J. Biochem. Biophys. 2013, 50, 114–119. [Google Scholar] [PubMed]
- Wang, H.; Peng, R.; Zhou, H.; Wang, S.; Gao, Y.; Wang, L.; Yong, Z.; Zuo, H.; Zhao, L.; Dong, J.; et al. Impairment of long-term potentiation induction is essential for the disruption of spatial memory after microwave exposure. Int. J. Radiat. Biol. 2013, 89, 1100–1107. [Google Scholar] [CrossRef]
- Qiao, S.; Peng, R.; Yan, H.; Gao, Y.; Wang, C.; Wang, S.; Zou, Y.; Xu, X.; Zhao, L.; Dong, J.; et al. Reduction of phosphorylated synapsin I (Ser-553) leads to spatial memory impairment by attenuating GABA release after microwave exposure in wistar rats. PLoS ONE 2014, 9, e95503. [Google Scholar] [CrossRef] [PubMed]
- Durney, C.H.; Iskander, M.F.; Massoudi, H.; Johnson, C.C. An empirical formula for broad band SAR calculations of prolate spheroidal models of humans and animal. In Biological Effects of Electromagnetic Radiation; Osepchuk, J.M., Ed.; IEEE Press: New York, NY, USA, 1984; pp. 85–90. [Google Scholar]
- Sharma, A.; Sisodia, R.; Bhatnagar, D.; Saxena, V.K. Spatial memory and learning performance and its relationship to protein synthesis of Swiss albino mice exposed to 10 GHz microwaves. Int. J. Radiat. Biol. 2014, 90, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Peng, R.; Zhao, L.; Wang, S.; Gao, Y.; Wang, L.; Zuo, H.; Dong, J.; Xu, X.; Zhou, H.; et al. The relationship between NMDA receptors and microwave-induced learning and memory impairment: A long-term observation on Wistar rats. Int. J. Radiat. Biol. 2015, 91, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Peng, R.Y.; Wang, C.Z.; Qiao, S.M.; Yong, Z.; Gao, Y.B.; Xu, X.P.; Wang, S.X.; Dong, J.; Zuo, H.Y.; et al. Alterations of cognitive function and 5-HT system in rats after long term microwave exposure. Physiol. Behav. 2015, 140, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Z.; Gao, Y.; Zhang, C. Effects of fetal microwave radiation exposure on offspring behavior in mice. J. Radiat. Res. 2015, 56, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, P.S.; Megha, K.; Nasare, N.; Banerjee, B.D.; Ahmed, R.S.; Abegaonkar, M.P.; Tripathi, A.K.; Mediratta, P.K. Effect of low level subchronic microwave radiation on rat brain. Biomed. Environ. Sci. 2016, 29, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, P.S.; Nasare, N.; Megha, K.; Banerjee, B.D.; Ahmed, R.S.; Singh, D.; Abegaonkar, M.P.; Tripathi, A.K.; Mediratta, P.K. Cognitive impairment and neurogenotoxic effects in rats exposed to low-intensity microwave radiation. Int. J. Toxicol. 2015, 34, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Zhang, Y.; Yang, L.; Chen, Q.; Tan, L.; Zuo, S.; Feng, H.; Chen, Z.; Zhu, G. Exposure to 900 MHz electromagnetic fields activates the mkp-1/ERK pathway and causes blood-brain barrier damage and cognitive impairment in rats. Brain Res. 2015, 1601, 92–101. [Google Scholar] [CrossRef]
- Shahin, S.; Banerjee, S.; Singh, S.P.; Chaturvedi, C.M. 2.45 GHz microwave radiation impairs learning and spatial memory via oxidative/nitrosative stress induced p53-dependent/independent hippocampal apoptosis: Molecular basis and underlying mechanism. Toxicol. Sci. 2015, 148, 380–399. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Wang, H.; Xu, X.; Zhao, L.; Zhang, J.; Dong, J.; Yao, B.; Wang, H.; Zhou, H.; Gao, Y.; et al. Study on dose-dependent, frequency-dependent, and accumulative effects of 1.5 GHz and 2.856 GHz microwave on cognitive functions in Wistar rats. Sci. Rep. 2017, 7, 10781. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Tan, S.; Xu, X.; Zhao, L.; Zhang, J.; Yao, B.; Gao, Y.; Zhou, H.; Peng, R. Long term impairment of cognitive functions and alterations of NMDAR subunits after continuous microwave exposure. Physiol. Behav. 2017, 181, 1–9. [Google Scholar] [CrossRef]
- Sharma, A.; Kesari, K.K.; Saxena, V.K.; Sisodia, R. Ten gigahertz microwave radiation impairs spatial memory, enzymes activity, and histopathology of developing mice brain. Mol. Cell. Biochem. 2017, 435, 1–13. [Google Scholar] [CrossRef]
- Shahin, S.; Banerjee, S.; Swarup, V.; Singh, S.P.; Chaturvedi, C.M. From the cover: 2.45-GHz microwave radiation impairs hippocampal learning and spatial memory: Involvement of local stress mechanism-induced suppression of iGluR/ERK/CREB signaling. Toxicol. Sci. 2018, 161, 349–374. [Google Scholar] [CrossRef]
- Kumlin, T.; Iivonen, H.; Miettinen, P.; Juvonen, A.; van Groen, T.; Puranen, L.; Pitkäaho, R.; Juutilainen, J.; Tanila, H. Mobile phone radiation and the developing brain: Behavioral and morphological effects in juvenile rats. Radiat. Res. 2017, 168, 471–479. [Google Scholar] [CrossRef]
- Mori, T.; Arendash, G.W. Long-term electromagnetic field treatment increases brain neuronal activity: Linkage to cognitive benefit and therapeutic implications for Alzheimer’s disease. J. Alzheimers Dis. 2011, 1, 102–105. [Google Scholar] [CrossRef]
- Arendash, G.W.; Mori, T.; Dorsey, M.; Gonzalez, R.; Tajiri, N.; Borlongan, C. Electromagnetic treatment to old Alzheimer’s mice reverses β-amyloid deposition, modifies cerebral blood flow, and provides selected cognitive benefit. PLoS ONE 2012, 7, e35751. [Google Scholar] [CrossRef]
- Jeong, Y.J.; Kang, G.Y.; Kwon, J.H.; Choi, H.D.; Pack, J.K.; Kim, N.; Lee, Y.S.; Lee, H.J. 1950 MHz electromagnetic fields ameliorate Ab pathology in Alzheimer’s disease mice. Curr. Alzheimer Res. 2015, 12, 481–492. [Google Scholar] [CrossRef]
- Cobb, B.L.; Jauchem, J.R.; Mason, P.A.; Dooley, M.P.; Miller, S.A.; Ziriax, J.M.; Murphy, M.R. Neural and behavioral teratological evaluation of rats exposed to ultra-wideband electromagnetic fields. Bioelectromagnetics 2000, 21, 524–537. [Google Scholar] [CrossRef]
- Johnson, C.C.; Durney, C.H.; Barber, P.W.; Massoudi, H.; Allen, S.J.; Mitchell, J.C. Radiofrequency Dosimetry Handbook, 1st ed.; Report SAM-TR-76-35; USAF School of Aerospace Medicine, Brooks Air Force Base: San Antonio, TX, USA, 1976. [Google Scholar]
- Sienkiewicz, Z.J.; Blackwell, R.P.; Haylock, R.G.; Saunders, R.D.; Cobb, B.L. Low-level exposure to pulsed 900 MHz microwave radiation does not cause deficits in the performance of a spatial learning task in mice. Bioelectromagnetics 2000, 21, 51–158. [Google Scholar] [CrossRef]
- Dubreuil, D.; Jay, T.; Edeline, J.-M. Does head-only exposure to GSM-900 electromagnetic fields affect the performance of rats in spatial learning tasks? Behav. Brain Res. 2002, 129, 203–210. [Google Scholar] [CrossRef]
- Dubreuil, D.; Jay, T.; Edeline, J.-M. Head-only exposure to GSM 900-MHz electromagnetic fields does not alter rat’s memory in spatial and non-spatial tasks. Behav. Brain Res. 2003, 145, 51–61. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Tsurita, G.; Ueno, S.; Watanabe, S.; Wake, K.; Taki, M.; Nagawa, H. 1439 MHz pulsed TDMA fields affect performance of rats in a T-maze task only when body temperature is elevated. Bioelectromagnetics 2003, 24, 223–230. [Google Scholar] [CrossRef]
- Cobb, B.L.; Jauchem, J.R.; Adair, E.R. Radial arm maze performance of rats following repeated low level microwave radiation exposure. Bioelectromagnetics 2004, 25, 49–57. [Google Scholar] [CrossRef]
- Cassel, J.-C.; Cosquer, B.; Galani, R.; Kuster, N. Whole-body exposure to 2.45 GHz electromagnetic fields does not alter radial-maze performance in rats. Behav. Brain Res. 2004, 155, 37–43. [Google Scholar] [CrossRef]
- Cosquer, B.; Vasconcelos, A.P.; Fröhlich, J.; Cassel, J.-C. Blood-brain barrier and electromagnetic fields: Effects of scopolamine methylbromide on working memory after whole-body exposure to 2.45 GHz microwaves in rats. Behav. Brain Res. 2005, 161, 229–237. [Google Scholar] [CrossRef]
- Cosquer, B.; Kuster, N.; Cassel, J.-C. Whole-body exposure to 2.45 GHz electromagnetic fields does not alter 12-arm radial-maze with reduced access to spatial cues in rats. Behav. Brain Res. 2005, 161, 331–334. [Google Scholar] [CrossRef]
- Cosquer, B.; Galani, R.; Kuster, N.; Cassel, J.-C. Whole-body exposure to 2.45 GHz electromagnetic fields does not alter anxiety responses in rats: A plus-maze study including test validation. Behav. Brain Res. 2005, 156, 65–74. [Google Scholar] [CrossRef]
- Ammari, M.; Jacquet, A.; Lecomte, A.; Sakly, M.; Abdelmelek, H.; de Seze, R. Effect of head-only sub-chronic and chronic exposure to 900-MHz GSM electromagnetic fields on spatial memory in rats. Brain Inj. 2008, 22, 1021–1029. [Google Scholar] [CrossRef]
- Daniels, W.M.; Pitout, I.L.; Afullo, T.J.; Mabandla, M.V. The effect of electromagnetic radiation in the mobile phone range on the behaviour of the rat. Metab. Brain Dis. 2009, 24, 629–641. [Google Scholar] [CrossRef]
- Takahashi, S.; Imai, N.; Nabae, K.; Wake, K.; Kawai, H.; Wang, J.; Watanabe, S.; Kawabe, M.; Fujiwara, O.; Ogawa, K.; et al. Lack of adverse effects of whole-body exposure to a mobile telecommunication electromagnetic field on the rat fetus. Radiat. Res. 2010, 173, 362–372. [Google Scholar] [CrossRef]
- Spathmann, O.; Hansen, V.; Streckert, J.; Zhou, Y.; Clemens, M.; Grote, K.; Klose, M.; Lerchl, A. “Head only”—Exposure of continuously growing rats to 900 MHz GSM signals. In Proceedings of the International Symposium on Electromagnetic Compatibility (EMC EUROPE), Rome, Italy, 17–21 September 2012; IEEE Xplore Digital Library: Rome, Italy, 2012. [Google Scholar] [CrossRef]
- Klose, M.; Grote, K.; Spathmann, O.; Streckert, J.; Clemens, M.; Hansen, V.W.; Lerchl, A. Effects of early-onset radiofrequency electromagnetic field exposure (GSM 900 MHz) on behavior and memory in rats. Radiat. Res. 2014, 182, 435–447. [Google Scholar] [CrossRef]
- Engels, S.; Schneider, N.L.; Lefeldt, N.; Hein, C.M.; Zapka, M.; Michalik, A.; Elbers, D.; Kittel, A.; Hore, P.J.; Mouritsen, H. Anthropogenic electromagnetic noise disrupts magnetic compass orientation in a migratory bird. Nature 2014, 509, 353–356. [Google Scholar] [CrossRef]
- Shirai, T.; Imai, N.; Wang, J.; Takahashi, S.; Kawabe, M.; Wake, K.; Kawai, H.; Watanabe, S.; Furukawa, F.; Fujiwara, O. Multigenerational effects of whole body exposure to 2.14 GHz W-CDMA cellular phone signals on brain function in rats. Bioelectromagnetics 2014, 35, 497–511. [Google Scholar] [CrossRef]
- Bouji, M.; Lecomte, A.; Gamez, C.; Blazy, K.; Villégier, A.S. Neurobiological effects of repeated radiofrequency exposures in male senescent rats. Biogerontology 2016, 17, 841–857. [Google Scholar] [CrossRef]
- Son, Y.; Jeong, Y.J.; Kwon, J.H.; Choi, H.D.; Pack, J.K.; Kim, N.; Lee, Y.S.; Lee, H.J. 1950 MHz radiofrequency electromagnetic fields do not aggravate memory deficits in 5xFAD mice. Bioelectromagnetics 2016, 37, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.P.; Zhang, K.Y.; Guo, L.; Chen, Q.L.; Gao, P.; Wang, T.; Li, J.; Guo, G.Z.; Ding, G.R. Effects of 1.8 GHz radiofrequency fields on the emotional behavior and spatial memory of adolescent mice. Int. J. Environ. Res. Public Health 2017, 14, 1344. [Google Scholar] [CrossRef]
- Shirai, T.; Wang, J.; Kawabe, M.; Wake, K.; Watanabe, S.-I.; Takahashi, S.; Fujiwara, O. No adverse effects detected for simultaneous whole-body exposure to multiple-frequency radiofrequency electromagnetic fields for rats in the intrauterine and pre- and post-weaning periods. J. Radiat. Res. 2017, 58, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Lai, H. Comment on “Radial arm maze performance of rats following repeated low level microwave radiation exposure”. Bioelectromagnetics 2005, 26, 81. [Google Scholar] [CrossRef]
- Wiltschko, W.; Wiltschko, R. Magnetic orientation in birds. J. Exp. Biol. 1996, 199 Pt 1, 29–38. [Google Scholar]
- Hodges, H. Maze procedures: The radial-arm and water maze compared. Cogn. Brain Res. 1995, 3, 167–181. [Google Scholar] [CrossRef]
- Voorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef]
- Barnhart, C.D.; Yang, D.; Lein, P.J. Using the Morris water maze to assess spatial learning and memory in weanling mice. PLoS ONE 2015, 10, e0124521. [Google Scholar] [CrossRef]

| Model | Exposure | Response | Comment | Reference |
|---|---|---|---|---|
| 12-arm radial maze SD rat (n = 8) 250–300 g | 2450 MHz, pulsed; 2 μs pulses at 500 pps 45 min/day, 10 days WBA SAR 0.6 W/kg Brain local SAR 0.5–2.0 W/kg Restrained | RF alone: more errors than sham. Pre-treatment with physostigmine or naltrexone: no difference exposed/sham. Pre-treatment with naloxone: no effect | Behaviour assessed after each daily exposure. SAR measured according to Chou et al. [7] | Lai et al. [8] |
| MWM SD rat (n = 11, 12) 2–3 months 250–300 g | 2450 MHz pulsed; 2 μs pulses at 500 pps 60 min × 2/day, 3 days WBA SAR 1.2 W/kg Restrained | Increased escape times, no effect on speed; less time in correct quadrant during probe trial | Differences in probe trial not significant using ANOVA, but significant using Newman–Keuls post-hoc analysis. SAR measured according to Chou et al. [7] | Wang and Lai [9] |
| MWM SD rat (n = 8) 2–3 months 250–300 g | 2450 MHz CW 60 min × 2/day, 3 days WBA SAR 1.2 W/kg Temporally incoherent magnetic noise at 6 µT Restrained | Increased escape times, less time in correct quadrant during probe trial; smaller changes after co-exposure with magnetic noise | Magnetic noise alone had no effect. SAR measured according to Chou et al. [7] | Lai [10] |
| MWM Wistar rat (n = 5) 3 months | Pulsed 2450 MHz ± glucocorticoid receptor antagonist RU468 3 h/day, 30 days Brain SAR 0.7 W/kg; WBA SAR 0.2 W/kg Free | RF: increased escape latency on day 4–6; RF + RU468: on day 6. RF: impaired memory RF + U468: no effects on memory | SAR calculated by calorimetry. Brain SAR seems doubtful | Li et al. [11] |
| MWM, activity wheel Parkes mouse (n = 5) 40 days | 2450 MHz CW WBA SAR 0.03 W/kg 120 min/day, 30 days Restrained | No effect on escape latency during acquisition; less time in correct quadrant during probe trial. Phase shift in activity after 30 days, less active at night | Mice given 20 s to locate the escape platform Size of pool not reported. Modest group sizes, non-standard testing protocol. SAR calculated according to Gandhi [12] | Chaturvedi et al. [13] |
| MWM, 8-arm radial maze Wistar rat (n = 6) 3 months | 2450 MHz, pulsed 10 µs, 800 pps WBA SAR 0.2 W/kg Brain SAR 0.7 W/kg 3 h/day for 30 days Free | Deficits in both tasks reduced by i.p. injection of glucose before each trial | SAR calculated using calorimetry | Lu et al. [14] |
| MWM Fischer rat (n = 6) 150–200 g | 900 MHz WBA SAR 0.085 mW/kg 2 h/day, 5 days/week for 30 days Restrained | Increased time to locate, and decreased time in target quadrant in probe trial | SAR used is very low | Deshmukh et al. [15] |
| MWM Wistar rat (n = 10) 8 weeks | 2856 MHz, pulsed 200 or 500 pps, pulse width 500 ns 6 min Average brain SAR 3.5, 7 or 35 W/kg Restrained | Increased escape latency at 6 h after exposure at 7 or 35 W/kg; and for 24 h after exposure at 35 W/kg. Reduced number of crossings of platform location in probe trial at 7 W/kg and 35 W/kg | Highest SAR caused a rise in brain temperature of 1.2 °C and in rectal temperature of 0.6 °C of anesthetized rats. SAR calculated using FDTD methods | Wang et al. [16] |
| MWM Wistar rat (n = 15) 200 g | 2856 MHz, pulsed 500 pps pulse width 500 ns 5 min WBA SAR 14 W/kg Restrained | Increased escape latency 1 day, 2, 3 and 7 days after exposure. Non-significant increase after 6 h, 4 and 14 days. | Rise in body temperature of 0.3 °C. No probe trial. SAR calculated using FDTD methods | Qiao et al. [17] |
| MWM Swiss albino mouse (n = 6) 6–8 weeks | 10 GHz WBA SAR 0.18 W/kg 2 h/day, 30 days Restrained | Increased escape latency | Two mice exposed together in same cage. No probe trial. SAR calculated according to Durney et al. [18] | Sharma et al. [19] |
| MWM Wistar rat (n = 15) 8 weeks | 2856 MHz, pulsed 500 pps, pulse width 500 ns 6 min Brain SAR 35 W/kg WBA SAR 15 W/kg Restrained | Increased escape latency up to 18 months after exposure | Rise in brain temperature of 1.2 °C and in rectal temperature of 0.6 °C of anesthetized rats. No probe trial. SAR calculated using FDTD methods | Wang et al. [20] |
| MWM Wistar rat (n = 15) 4 weeks | 2.856 GHz 6 min × 3 per week, 6 weeks PD 5, 10, 20 or 30 mW/cm2 (50, 100, 200, 300 W/m2) Restrained | Escape latency increased at 5 mW/cm2 at 14 days, at 10 mW/cm2 at 4, 14, 28 days and at 20 and 30 mW/cm2 at 3, 4, 14, 28 days after exposure. All exposed groups spent less time in the target quadrant in probe trial 5 days after exposure, and escape latency increased at 14 days after exposure | WBA SAR 1.5, 3, 6 or 9 W/kg estimated from Wang et al. [16,20] | Li et al. [21] |
| MWM, OFA, EPM, tail suspension, forced swim CD-1 mouse 5 weeks old | 9.417 GHz 200 V/m (SAR 2 W/kg) 12 h/day from gestational day 3.5 to 18 Free | Impaired learning and memory only in male mice. Increased anxiety and decreased depression in males and females | Basis of SAR calculation not given | Zhang et al. [22] |
| MWM Fischer 344 rat (n = 6) 180 days | 900, 1800 or 2450 MHz 2 h/day, 5 days/week for 90 days WBA SAR 0.59, 0.58, 0.67 mW/kg Restrained | All exposures impaired performance in probe trial, with increased time to target quadrant and decreased time in quadrant | Also increased HSP70 levels and increased DNA strand breaks Identical results published by Deshmukh et al. [23] following exposure for 90 days | Deshmukh et al. [24] |
| MWM SD rat (n = 9) 220–250 g | 900 MHz CW 3 h/day for 14 or 28 days Average SAR in the head 2 W/kg, WBA SAR 0.016 W/kg. Restrained | No effects on learning, exposure for 28 days significantly impaired memory | Ultrastructural changes and increased serum albumin leakage. Effects attributed to changes in mkp-1/ERK pathway | Tang et al. [25] |
| MWM Swiss mouse (n = 20) 12 weeks | 2450 MHz CW 2 h/day for 15, 30 for 60 days WBA SAR 0.0146 W/kg Restrained | Significantly increased escape latency during acquisition, impaired memory in probe trial. Deficits increased with increasing exposure time | No effect on rectal temperature. Exposure time-dependent changes in neuronal morphology, apoptosis, oxidative state | Shahin et al. [26] |
| MWM Wistar rat (n = 25) 200 g | 1500 MHz, 2856 MHz or both sequentially 6 min/frequency WBA SAR 1.8/1.7 W/kg or 3.7/3.3 W/kg Free | Escape latency increased only at higher SAR (both frequencies). No increased effect with sequential exposure | Skin temperature increased by <1 °C (n = 4). Changes in EEG and hippocampal morphology at higher SAR | Tan et al. [27] |
| MWM Wistar rat (n = 15) 8 weeks | 2586 MHz, pulsed 6 min/day, 5 days/week for 6 weeks Brain SAR 1.7, 3.5 or 7 W/kg Restrained | Effects only at 7 W/kg: escape latency increased; impaired memory in probe trial. | No change in measured body temperature (n = 10). Changes in EEG, NMDAR, hippocampal structure | Wang et al. [28] |
| MWM Swiss mouse (n = 6) 14 days | 1000 MHz 2 h/day for 15 days WBA SAR 0.179 W/kg Lightly restrained | Deficits in learning and memory when tested at 6 weeks of age | Biochemical and histological changes | Sharma et al. [29] |
| Radial arm maze, 4/8 task Swiss mouse (n = 20) 12 weeks | 2450 MHz CW 2 h/day for 15, 30 or 60 days WBA SAR 14.6 mW/kg | Exposure-time dependent increase in errors in working and reference memory | No increase in rectal temperature | Shahin et al. [30] |
| Model | Exposure | Response | Comment | Reference |
|---|---|---|---|---|
| MWM, OFA, EPM Wistar rat (n = 18) 60.5–60.7 g (24 days) | 900 MHz, GSM 2 h/day, 5 days/week for 5 weeks WBA SAR 0.3, 3 W/kg Free | Improved learning of escape platform with both SARs; improved memory of platform location with 3 W/kg | No effect on activity, anxiety, blood brain barrier | Kumlin et al. [31] |
| Y maze APPsw mouse, Tg or NT (n = 4–10) 21–26 months | 918 MHz GSM E field 17–35 V/m 2 × 2 h/day, 1 month Free | Alternations in Y maze increased 26% when Tg and NT mice combined | SAR provided, but incorrectly calculated from external E field | Mori and Arendash [32] |
| MWM, radial arm maze, Y maze, circular platform APPsw mouse, Tg or NT (n = 5–9) 21–26 months | 918 MHz GSM E field 17–35 V/m 2 × 2 h/day, up to 2 months Free | No field-dependent effects except alternations in Y maze increased when Tg and NT mice were combined (p < 0.05) | Exposure continued during testing period. No effects on agility, activity or exploration.SAR provided, but incorrectly calculated from external E field | Arendash et al. [33] |
| Y maze, OFA, passive avoidance tests 5xFAD mouse and WT (n = 7–11) 1.5 months | 1950 MHz W-CDMA 2 h/day, 5 day/week for 8 months WBA SAR 5 W/kg Free | Decreased alternation in Y maze, increased time in centre of OFA, impairments in passive avoidance, all rescued by exposure; no effects on WT | Maximum increase of body temperature of 0.5 °C Aβ plagues and other pathology reduced by exposure | Jeong et al. [34] |
| Model | Exposure | Response | Comment | Reference |
|---|---|---|---|---|
| MWM, hippocampus morphology of offspring after exposure SD rat (n = 6) Day 3–18 of pregnancy (maze) or day 3–18 plus 1–10 postnatal day (morphology) | 0.1–1 GHz ultra-wideband pulses 2 min/day, 16 days prenatally, 10 days postnatally WBA SAR 45 mW/kg Restrained | No effects on maze task (males only, on day 50), increased medial-to-lateral length of the hippocampus (day 21). Overall, no effects on 39 out of 42 endpoints | Clear responses to positive control (lead acetate). SAR calculated from power spectrum | Cobb et al. [35] |
| 8-arm radial maze C57BL/6J mouse (n = 5) 12 weeks | 900 MHz pulsed at 217 Hz 45 min/day, 10 days WBA SAR 0.05 W/kg Tested immediately or 15 or 30 min after exposure Restrained | No effects on performance | Animals tested immediately took longer to complete task both after RF and sham exposure. SAR calculated according to Johnson et al. [36] | Sienkiewicz et al. [37] |
| 8-arm radial maze. spatial task in OFA SD rat (n = 8) 150 g | 900 MHz pulsed at 217 Hz 45 min/day, 10 days (radial maze) or 14 days (spatial task) Brain average SAR 1 or 3.5 W/kg Restrained | No effects | Head-only exposure. SAR calculated from temperature measurements and using FDTD methods | Dubreuil et al. [38] |
| Two versions of 8-arm radial maze SD rat (n = 12 or 9) 120 g | 900 MHz pulsed at 217 Hz 45 or 60 min/day, 4, 12 or 16 days Brain SAR 1 or 3.5 W/kg Restrained | No effects | Head-only exposure. SAR calculated from temperature measurements and using FDTD methods | Dubreuil et al. [39] |
| T-maze reversal learning SD rat (n = 15–28) 670 g | 1439 MHz pulsed 6.7 ms pulses at 50 pps 45 or 60 min/day, 4 days or 60 min/day, 4 × 5 days Brain average SAR 7.5 or 25 W/kg Restrained | No effect on performance at lower SAR, decreased performance at higher SAR resulting in increased core temperature | Head-mainly exposure (animals positioned with head towards antenna). SAR calculated from temperature measurements and using FDTD methods | Yamaguchi et al. [40] |
| 12-arm radial maze SD rat (n = 7 or 8) 250–300 g | 2450 MHz pulsed; 2 μs pulses at 500 pps 45 min/day, 10 days WBA SAR 0.6 W/kg Restrained | No effects on performance and no effect of treatment with physostigmine, naltrexone or naloxone | Did not confirm Lai at al. [8]. SAR calculated using calorimetry and input/output difference | Cobb et al. [41] |
| 12-arm radial maze SD rat (n = 12) 3 months, 270–320 g | 2450 MHz pulsed; 2 μs pulses at 500 pps 45 min/day, 10 days WBA SAR 0.6 W/kg Restrained | No effects on performance in maze with access to distal spatial cues | Did not confirm Lai at al. [8]. SAR calculated using FDTD methods | Cassel et al. [42] |
| 12-arm radial maze Sprague Dawley rat (n = 12) 3 months, 270–320 g | 2450 MHz pulsed; 2 μs pulses at 500 pps 45 min/day, 10 days WBA SAR 2 W/kg Restrained | No effects on performance in maze with access to distal spatial cues | Did not confirm Lai et al. [8]. SAR calculated using FDTD methods. | Cosquer et al. [43] |
| 12-arm radial maze SD rat (n = 12) 3 months, 270–320 g | 2450 MHz pulsed; 2 μs pulses at 500 pps 45 min/day, 10 days WBA SAR 2 W/kg Restrained | No effects on performance in maze with reduced access to distal spatial cues | Did not confirm Lai et al. [8]. SAR calculated using FDTD methods | Cosquer et al. [44] |
| EPM SD rat (n = 12) 3 months, 270–320 g | 2450 MHz pulsed; 2 μs pulses at 500 pps 45 min/day, 10 days WBA SAR 2 W/kg Restrained | No effect on anxiety with ambient light of 2.5 or 30 lux | SAR calculated using FDTD methods | Cosquer et al. [45] |
| 8-arm radial maze over 10 days with further 8 days with 45 min inter trial delay after 4 correct responses SD rat (n = 6) 6 weeks | 900 MHz GSM 45 min/day at average brain SAR 1.5 W/kg or 15 min/day at brain SAR 6 W/kg, 5 days/week, 8 or 24 weeks before testing Restrained | No effects | Head-only exposure. SAR calculated from temperature measurements and FDTD methods in Dubreuil et al. [39] | Ammari et al. [46] |
| MWM SD rat (n = 6) 2 days | 840 MHz 3 h/day, 12 days PD 60 µW/m2 Free moving | No effects | Increased freezing behaviour in males (mood disturbance) | Daniels et al. [47] |
| MWM, OFA CR1:CD(SD) rat (n = 42–28) Adults: 10 weeks + 5 days acclimatization Offspring: 4 days | 2140 MHz, W-CDMA 20 h/day, from day 7 of gestation to delivery and day 4–21 after birth Dams: WBA SAR 0.066–0.093, 0.028–0.04 W/kg; Foetus/progeny: WBA SAR 0.068–0.146, 0.029–0.067 W/kg Free | No effects on offspring | SAR calculated using FDTD methods | Takahashi et al. [48] |
| MWM, 8-arm radial maze (4/8 version), OFA Wistar rat (n = 20) 14 days | 900 MHz GSM Average brain SAR 0.7, 2.5 or 10 W/kg 2 h/day, 5 days/week, 18 months Restrained | No effects observed as juveniles, adults or pre-senile (more specific ages not given) | Head-only exposure. No probe trial. SAR calculated using FDTD methods as in Spathman et al. [49] | Klose et al. [50] |
| Orientation response to magnetic north European robin (n = 18–42) Age not specified | Background fields 0.01–5 MHz, 1008 nT Screened condition 0.01–5 MHz, 2.56 nT Applied fields 0.01–5 MHz, 278 nT; 0.02–0.45 MHz, 133 nT; 0.6–3 MHz, 34 nT Free | No response with background fields, or with applied fields. Response with screen | Exposures given as accumulated time-dependent magnetic field intensity summed over relevant frequency range | Engels et al. [51] |
| MWM, OFA SD rat (n = 16) 9 weeks | 2.14 GHz W-CDMA WBA SAR <0.24, <0.08 W/kg 20 h/day gestational day 7 to postnatal day 21 (weaning) then 24 h/day for 3 weeks Free | No consistent effects | SAR variable due to growth and movement No effects on development or anxiety | Shirai et al. [52] |
| Radial arm maze, OFA, EPM, fear conditioning Wistar rat (n = 6–8) 4–6 and 22–24 months | 900 MHz 2 h/day, 5 days/week for 4 weeks Brain SAR 6 W/kg Restrained | Age-related differences. No field-related effects except for decrease in anxiety | No field-related changes in IL-1β, IL-6 or GFAP levels | Bouji et al. [53] |
| MWM, Y maze, OFA, object recognition Mice: 5xFAD (n = 8) 1.5 months old | 1950 MHz 2 h/day, 5 days/week, for 3 months WBA SAR 5 W/kg Free | No effects | Measured rectal temperatures changed from −1.9 °C to +0.5 °C | Son et al. [54] |
| MWM, OFA, EPM C57/BL mouse (n = 10, 15) 4 weeks | 1800 MHz 6 h/day for 28 days WBA SAR 2.7 W/kg, Brain SAR 2.2 W/kg Free | No effects except decreased behavioural anxiety | No increase in skin temperature Increased levels of GABA and Asp in cortex and hippocampus. | Zhang et al. [55] |
| MWM, OFA SD rat (n = 8) Gestational day 7 | Multiple frequency signal, 880 to 5180 MHz 20 h/day from gestational day 7 to 6 weeks of age WBA SAR 0.08, 0.4 W/kg Free | No consistent effects | No consistent teratological or developmental effects | Shirai et al. [56] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sienkiewicz, Z.; van Rongen, E. Can Low-Level Exposure to Radiofrequency Fields Effect Cognitive Behaviour in Laboratory Animals? A Systematic Review of the Literature Related to Spatial Learning and Place Memory. Int. J. Environ. Res. Public Health 2019, 16, 1607. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16091607
Sienkiewicz Z, van Rongen E. Can Low-Level Exposure to Radiofrequency Fields Effect Cognitive Behaviour in Laboratory Animals? A Systematic Review of the Literature Related to Spatial Learning and Place Memory. International Journal of Environmental Research and Public Health. 2019; 16(9):1607. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16091607
Chicago/Turabian StyleSienkiewicz, Zenon, and Eric van Rongen. 2019. "Can Low-Level Exposure to Radiofrequency Fields Effect Cognitive Behaviour in Laboratory Animals? A Systematic Review of the Literature Related to Spatial Learning and Place Memory" International Journal of Environmental Research and Public Health 16, no. 9: 1607. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16091607