Effects of Simulated Herbivory on the Vegetative Reproduction and Compensatory Growth of Hordeum brevisubulatum at Different Ontogenic Stages


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Plant Species
2.3. Experimental Design
2.4. Harvest and Measurement
2.5. Data Analysis
3. Results
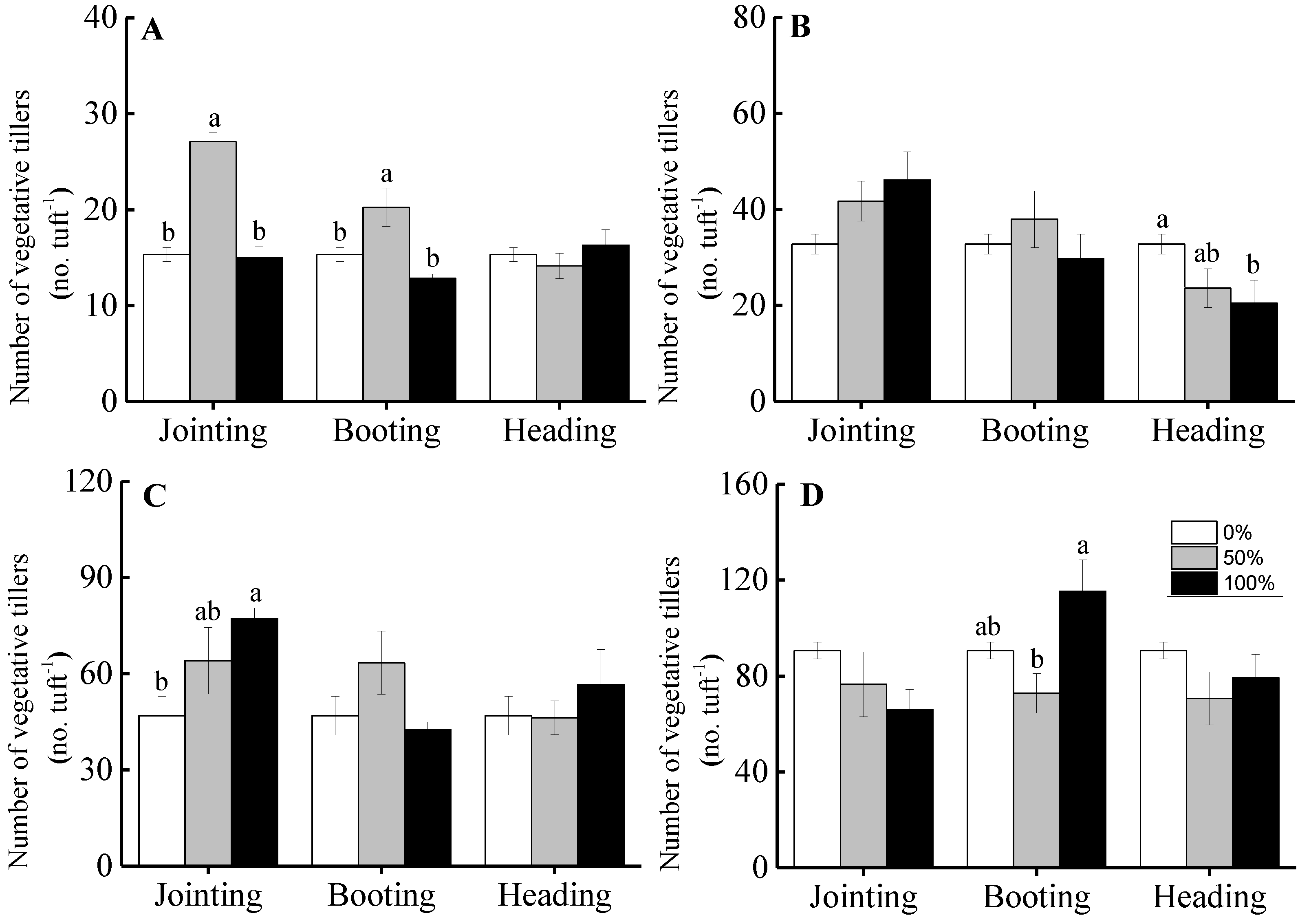
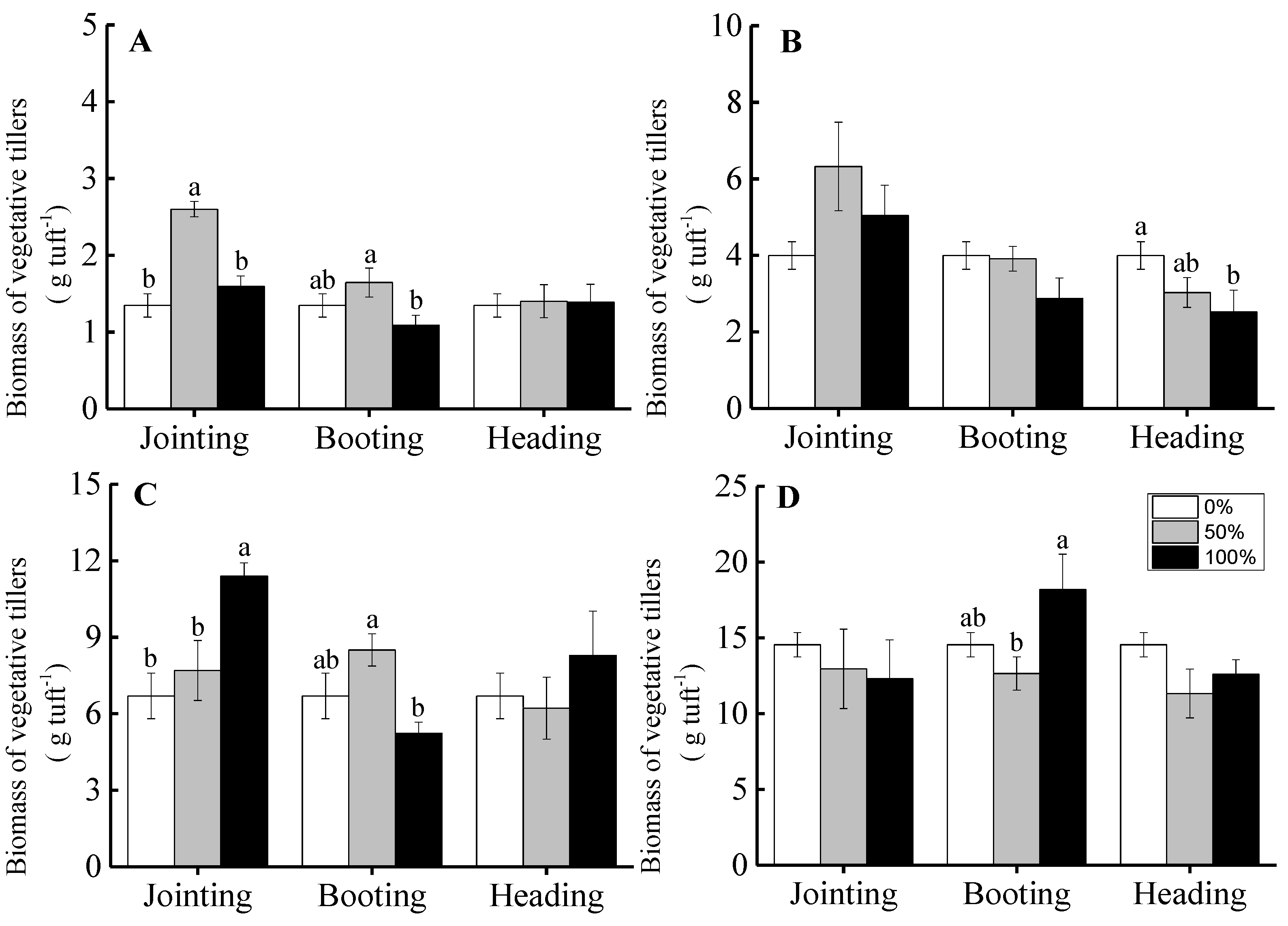
3.1. Vegetative Tillers
3.2. Total Number of Juvenile Tillers and Buds
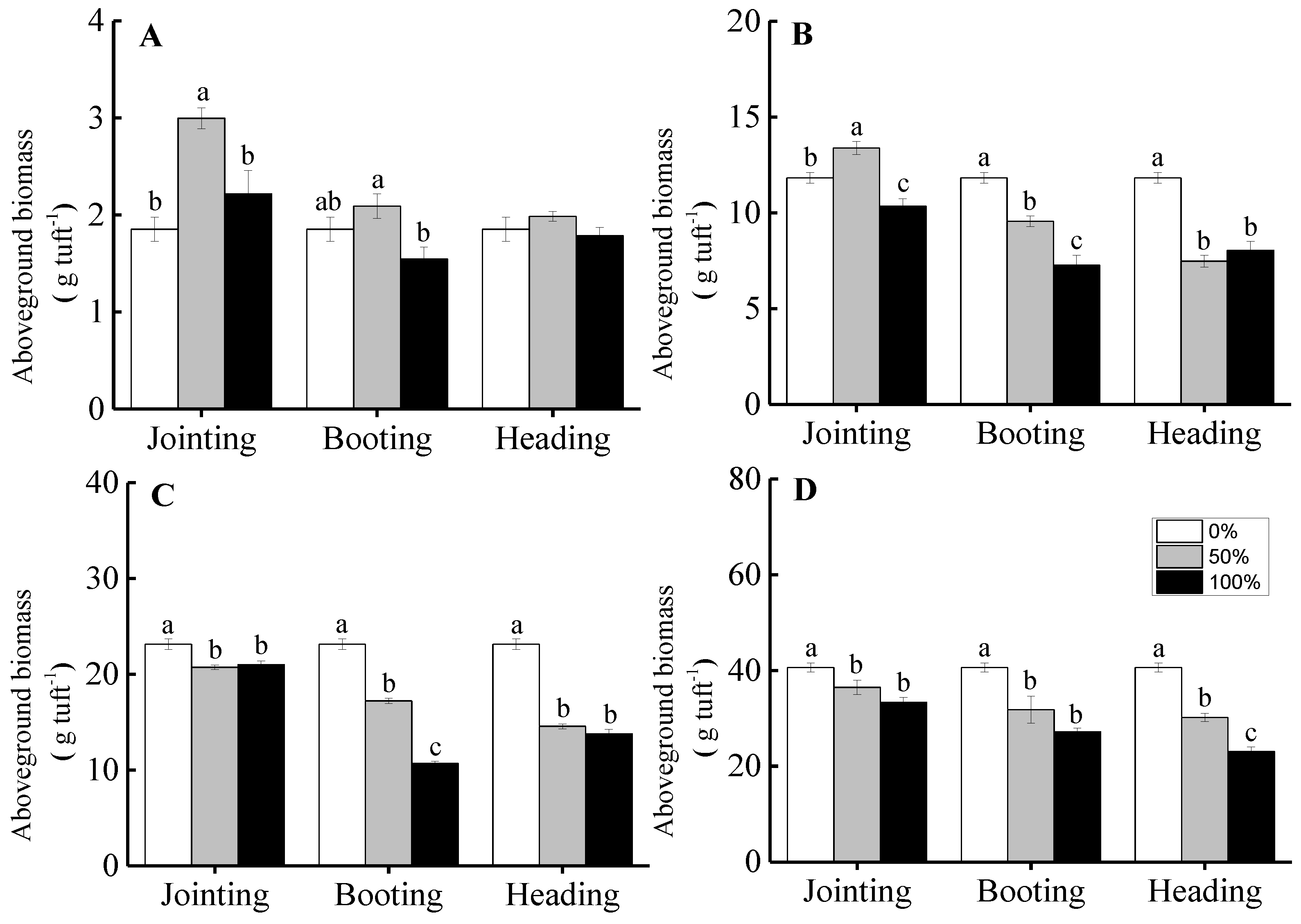
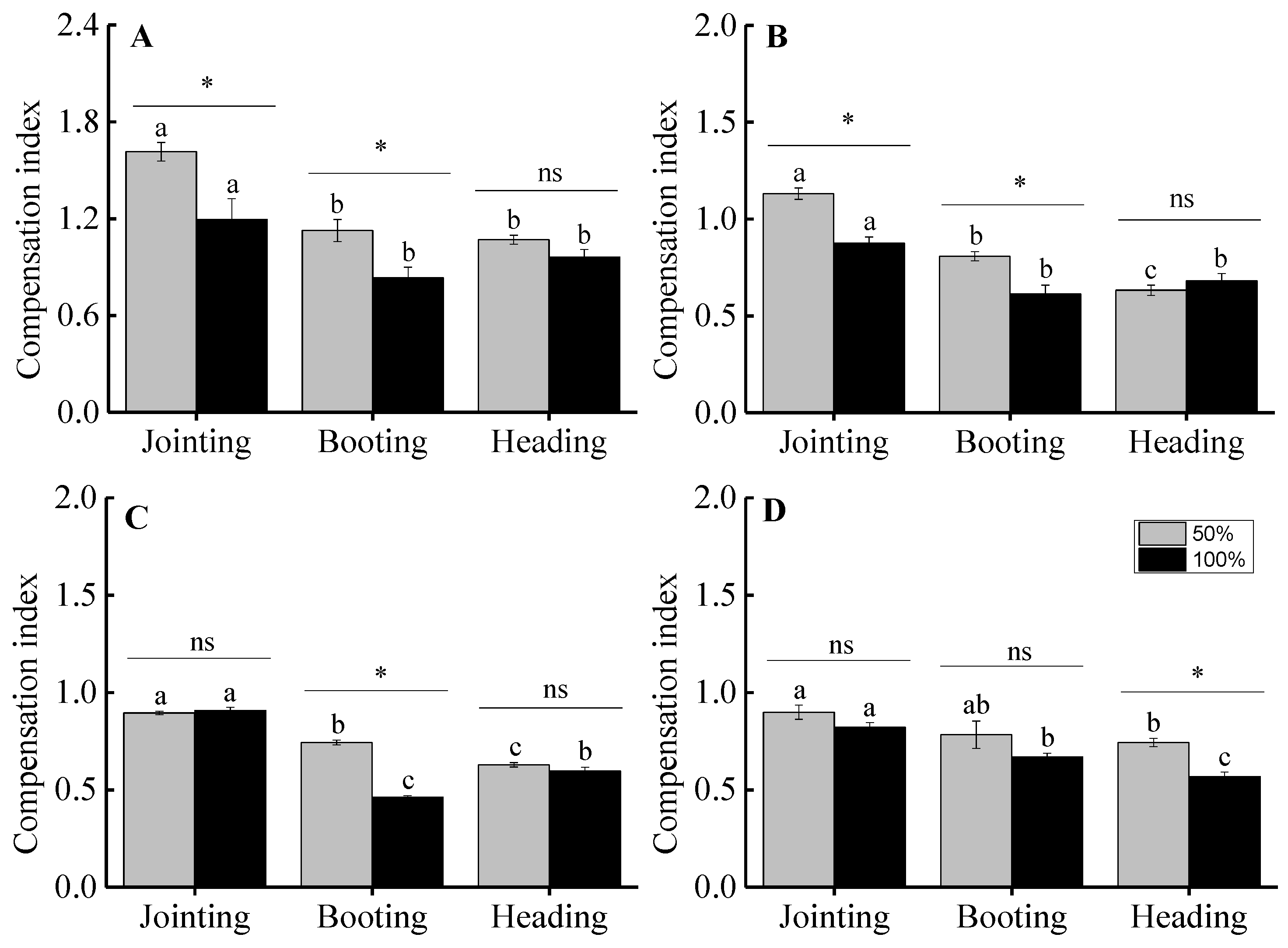
3.3. Aboveground Biomass and Compensation Index
4. Discussion
4.1. The Effects of Clipping on the Vegetative Reproduction of Plants
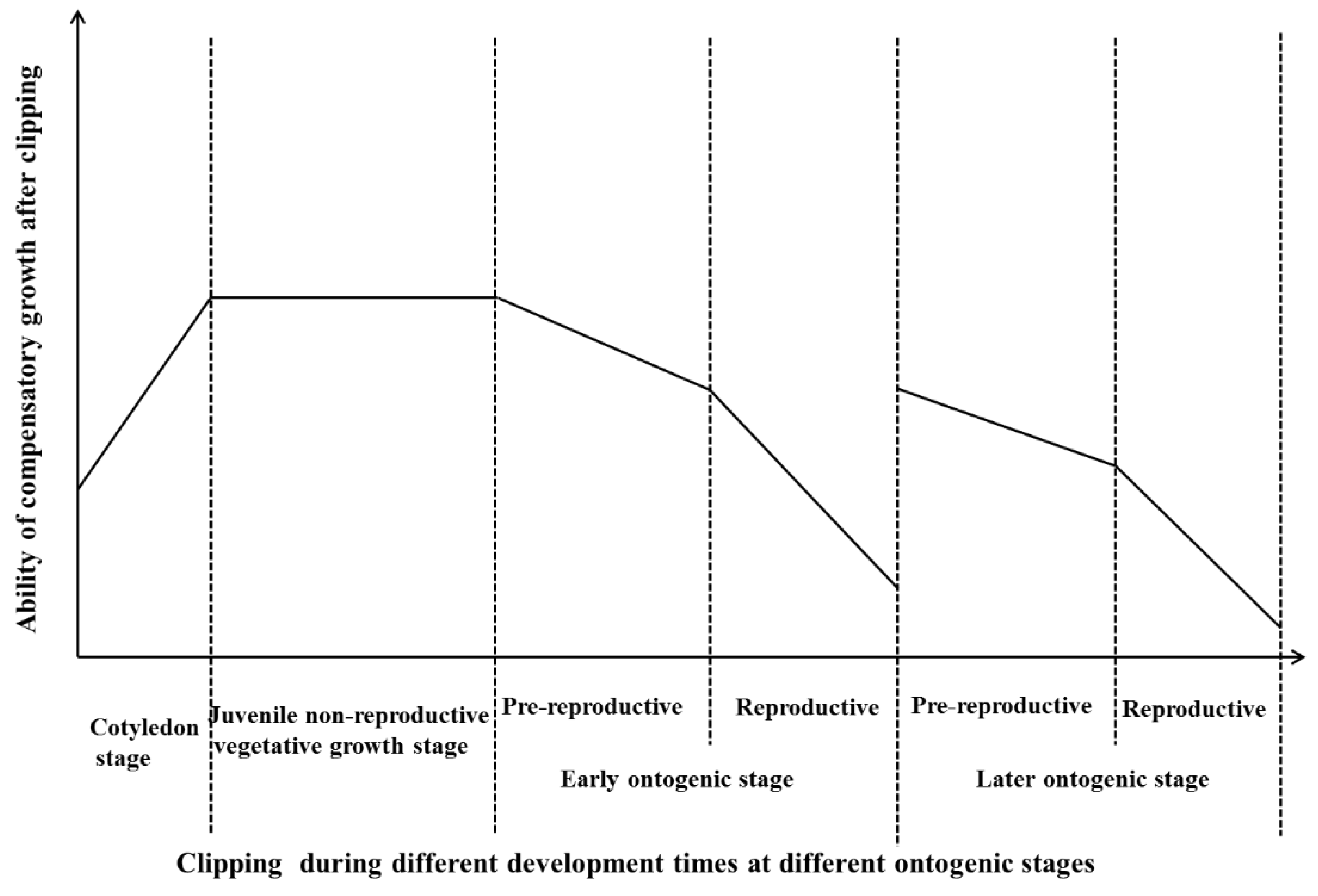
4.2. The Effects of Clipping on the Compensation Growth of Plants
5. Conclusions
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Gedge, K.E.; Maun, M.A. Effects of simulated herbivory on growth and reproduction of two beach annuals, Cakile edentula and Corispermum hyssopifolium. Can. J. Bot. 1992, 70, 2467–2475. [Google Scholar] [CrossRef]
- McNaughton, S.J. Compensatory plant growth as a response to herbivory. Oikos 1983, 40, 329–336. [Google Scholar] [CrossRef]
- Da Silveira Pontes, L.; Louault, F.; Carrère, P.; Maire, V.; Andueza, D.; Soussana, J.F. The role of plant traits and their plasticity in the response of pasture grasses to nutrients and cutting frequency. Ann. Bot. 2010, 105, 957–965. [Google Scholar] [CrossRef]
- Anten, N.P.R.; Ackerly, D.D. Canopy-level photosynthetic compensation after defoliation in a tropical understorey palm. Funct. Ecol. 2001, 15, 252–262. [Google Scholar]
- Del-Val, E.; Crawley, M.J. Are grazing increaser species better tolerators than decreasers? An experimental assessment of defoliation tolerance in eight British grassland species. J. Ecol. 2005, 93, 1005–1016. [Google Scholar] [CrossRef]
- Gruntman, M.; Novoplansky, A. Ontogenetic contingency of tolerance mechanisms in response to apical damage. Ann. Bot. 2011, 108, 965–973. [Google Scholar] [CrossRef]
- Holland, J.N.; Cheng, W.; Crossley, D.A. Herbivore-induced changes in plant carbon allocation: Assessment of below-ground C fluxes using carbon-14. Oecologia 1996, 107, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Chen, S.P.; Lin, G.H. Compensatory growth responses to clipping defoliation in Leymus chinensis (Poaceae) under nutrient addition and water deficiency conditions. Plant Ecol. 2008, 196, 85–99. [Google Scholar] [CrossRef]
- Tiffin, P. Mechanisms of tolerance to herbivore damage: What do we know? Evol. Ecol. 2000, 14, 523–536. [Google Scholar] [CrossRef]
- Tuomi, J.; Nilsson, P.; Astrom, M. Plant compensatory responses: Bud dormancy as an adaptation to herbivory. Ecology 1994, 75, 1429–1436. [Google Scholar] [CrossRef]
- Chen, X.S.; Deng, Z.M.; Xie, Y.H.; Li, F.; Hou, Z.Y.; Wu, C. Consequences of repeated defoliation on belowground bud banks of Carex brevicuspis (Cyperaceae) in the Dongting Lake wetlands, China. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Dalgleish, H.J.; Hartnett, D.C. The effects of fire frequency and grazing on tallgrass prairie productivity and plant composition are mediated through bud bank demography. Plant Ecol. 2009, 201, 411–420. [Google Scholar] [CrossRef]
- Escarre, J.; Lepart, J.; Sentuc, J.J. Effects of simulated herbivory in three old field Compositae with different inflorescence architectures. Oecologia 1996, 105, 501–508. [Google Scholar] [CrossRef]
- Hanley, M.E.; May, O.C. Cotyledon damage at the seedling stage affects growth and flowering potential in mature plants. New Phytol. 2006, 169, 243–250. [Google Scholar] [CrossRef]
- Tucker, C.; Avila-Sakar, G. Ontogenetic changes in tolerance to herbivory in Arabidopsis. Oecologia 2010, 164, 1005–1015. [Google Scholar] [CrossRef]
- Anderson, M.T.; Frank, D.A. Defoliation effects on reproductive biomass: Importance of scale and timing. J. Range Manag. 2003, 56, 501–516. [Google Scholar] [CrossRef]
- Massad, T.J. Ontogenetic differences of herbivory on woody and herbaceous plants: A meta-analysis demonstrating unique effects of herbivory on the young and the old, the slow and the fast. Oecologia 2013, 172, 1–10. [Google Scholar] [CrossRef]
- Tito, R.; Castellani, T.T.; Fáveri, S.B.; Lopes, B.C.; Vasconcelos, H.L. From over to undercompensation: Variable responses to herbivory during ontogeny of a Neotropical monocarpic plant. Biotropica 2016, 48, 608–617. [Google Scholar] [CrossRef]
- Boege, K.; Dirzo, R.; Siemens, D.; Brown, P. Ontogenetic switches from plant resistance to tolerance: Minimizing costs with age? Ecol. Lett. 2007, 10, 177–187. [Google Scholar] [CrossRef]
- Cisneros-Silva, A.; Castillo, G.; Chávez-Pesqueira, M.; Bello-Bedoy, R.; Camargo, I.D.; Núñez-Farfán, J. Light limitation reduces tolerance to leaf damage in Datura stramonium. Evol. Ecol. Res. 2017, 18, 351–362. [Google Scholar]
- Levine, M.T.; Paige, K.N. Direct and indirect effects of drought on compensation following herbivory in scarlet gilia. Ecology 2004, 85, 3185–3191. [Google Scholar] [CrossRef]
- Hanley, M.E.; Fegan, E.L. Timing of cotyledon damage affects growth and flowering in mature plants. Plant Cell Environ. 2007, 30, 812–819. [Google Scholar] [CrossRef]
- Weltzin, J.F.; Archer, S.R.; Heitschmidt, R.K. Defoliation and woody plant (Prosopis glandulosa) seedling regeneration: Potential vs. realized herbivory tolerance. Plant Ecol. 1998, 138, 127–135. [Google Scholar] [CrossRef]
- Boege, K. Influence of plant ontogeny on compensation to leaf damage. Am. J. Bot. 2005, 92, 1632–1640. [Google Scholar] [CrossRef]
- Ramula, S. Responses to the timing of damage in an annual herb: Fitness components versus population performance. Basic Appl. Ecol. 2008, 9, 233–242. [Google Scholar] [CrossRef]
- Boege, K.; Marquis, R.J. Facing herbivory as you grow up: The ontogeny of resistance in plants. Trends Ecol. Evol. 2005, 20, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.T.; Schmelz, E.A. Immunological “Memory” in the Induced Accumulation of Nicotine in Wild Tobacco. Ecology 1996, 77, 236–246. [Google Scholar] [CrossRef]
- Bowers, M.D.; Stamp, N.E. Effects of Plant Age, Genotype and Herbivory on Plantago Performance and Chemistry. Ecology 1993, 74, 1778–1791. [Google Scholar] [CrossRef]
- Hanley, M.E.; Fenner, M.; Edwards, P.J. The effect of seedling age on the likelihood of herbivory by the slug deroceras reticulatum. Funct. Ecol. 1995, 9, 754–759. [Google Scholar] [CrossRef]
- Li, J.D.; Zhang, W.Z. The natural conditions of alkaline grassland in Songnen plain. In The Bioecological Mechamism and Management of Alkaline Grassland in Songnen Plain; Li, J.D., Zheng, H.Y., Eds.; Science Press: Beijing, China, 1997; pp. 7–27. [Google Scholar]
- Li, H.Y.; Lindquist, J.L.; Yang, Y.F. Effects of sowing date on phenotypic plasticity of fitness-related traits in two annual weeds on the Songnen Plain of China. PLoS ONE 2015, 10, e0127795. [Google Scholar] [CrossRef]
- Wang, P.; Zhou, D.W.; Valentine, I. Seed maturity and harvest time effects seed quantity and quality of Hordeum brevisubulatum. Seed Sci. Technol. 2006, 34, 125–132. [Google Scholar] [CrossRef]
- Yang, Y.F.; Zhu, T.C. Plant Ecology, 2nd ed.; Higher Education Press: Beijing, China, 2011; pp. 202–203. [Google Scholar]
- Benzel, K.R.; Mosley, T.K.; Mosley, J.C. Defoliation timing effects on spotted knapweed seed production and viability. Rangel. Ecol. Manag. 2009, 62, 550–556. [Google Scholar] [CrossRef]
- Thomson, V.P.; Cunningham, S.A.; Ball, M.C.; Nicotra, A.B. Compensation for herbivory by Cucumis sativus through increased photosynthetic capacity and efficiency. Oecologia 2003, 134, 167–175. [Google Scholar] [CrossRef]
- Tolvanen, A.; Laine, K. Effects of reproduction and artificial herbivory on vegetative growth. Can. J. Bot. 1997, 75, 656–666. [Google Scholar] [CrossRef]
- Gruntman, M.; Novoplansky, A. Implications of local-scale productivity on compensatory growth in a semi-arid shrubland. J. Arid Environ 2011, 75, 279–283. [Google Scholar] [CrossRef]
- Long, J.D.; Porturas, L.D. Herbivore impacts on marsh production depend upon a compensatory continuum mediated by salinity stress. PLoS ONE 2014, 9, e110419. [Google Scholar] [CrossRef] [PubMed]
- Belsky, A.J. Does Herbivory Benefit Plants? A Review of the Evidence. Am. Nat. 1986, 127, 870–892. [Google Scholar] [CrossRef]
- Rooke, T.; Bergström, R. Growth, chemical responses and herbivory after simulated leaf browsing in Combretum apiculatum. Plant Ecol. 2007, 189, 201–212. [Google Scholar] [CrossRef]
- Chen, L.P.; Zhao, N.X.; Zhang, L.H.; Gao, Y.B. Responses of two dominant plant species to drought stress and defoliation in the Inner Mongolia Steppe of China. Plant Ecol. 2013, 214, 221–229. [Google Scholar] [CrossRef]
- Hilbert, D.W.; Swift, D.M.; Detling, J.K.; Dyer, M.I. Relative growth rates and the grazing optimization hypothesis. Oecologia 1981, 51, 14–18. [Google Scholar] [CrossRef]
- Hikosaka, K.; Takashima, T.; Kabeya, D.; Hirose, T.; Kamata, N. Biomass allocation and leaf chemical defence in defoliated seedlings of Quercus serrata with respect to carbon-nitrogen balance. Ann. Bot. 2005, 95, 1025–1032. [Google Scholar] [CrossRef]
- Mager, H.J.; Young, B.G.; Preece, J.E. Characterization of compensatory weed growth. Weed Sci. 2006, 54, 274–281. [Google Scholar] [CrossRef]
- Oesterheld, M.; Mcnaughton, S.J. Effect of stress and time for recovery on the amount of compensatory growth after grazing. Oecologia 1991, 85, 305–313. [Google Scholar] [CrossRef]
- Benson, E.J.; Mann, H.K.H. Belowground bud banks and meristem limitation in tallgrass prairie plant populations. Am. J. Bot. 2004, 91, 416–421. [Google Scholar] [CrossRef]
- Zhang, J.T.; Mu, C.S.; Wang, D.L.; Wang, J.F.; Chen, G.X. Shoot population recruitment from a bud bank over two seasons of undisturbed growth of leymus chinensis. Botany 2009, 87, 1242–1249. [Google Scholar] [CrossRef]
- Ferraro, D.O.; Oesterheld, M. Effect of defoliation on grass growth. A quantitative review. Oikos 2002, 98, 125–133. [Google Scholar] [CrossRef]
- Sullivan, A.T.; Howe, H.F. Prairie forb response to timing of vole herbivory. Ecology 2009, 90, 1346–1355. [Google Scholar] [CrossRef]
- Li, C.C.; Li, H.Y.; Yang, Y.F. Senescence in growth and reproductive allocation in a bunchgrass. Plant Biol. 2019, 21, 300–306. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indexes | O | P | I | O × P | O × I | P × I | O × P × I |
|---|---|---|---|---|---|---|---|
| Number of vegetative tillers | 193.04 ** | 4.08 * | 0.38 ns | 2.90 * | 3.41 ** | 1.26 ns | 3.18 ** |
| Biomass of vegetative tillers (g) | 235.34 ** | 3.16 * | 0.43 ns | 2.14 ns | 2.13 ns | 0.90 ns | 2.33 ** |
| Total number of juvenile tillers and buds | 275.55 ** | 0.99 ns | 3.99 * | 1.02 ns | 2.08 ns | 4.46 ** | 1.96 * |
| Aboveground biomass (g) | 3195.62 ** | 71.36 ** | 209.50 ** | 8.61 ** | 46.96 ** | 19.94 ** | 5.01 ** |
| Compensation index | 115.40 ** | 114.47 ** | 71.60 ** | 3.72 ** | 4.86 ** | 7.36 ** | 4.51 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, J.; Wang, P.; Yang, Y. Effects of Simulated Herbivory on the Vegetative Reproduction and Compensatory Growth of Hordeum brevisubulatum at Different Ontogenic Stages. Int. J. Environ. Res. Public Health 2019, 16, 1663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16091663
Yuan J, Wang P, Yang Y. Effects of Simulated Herbivory on the Vegetative Reproduction and Compensatory Growth of Hordeum brevisubulatum at Different Ontogenic Stages. International Journal of Environmental Research and Public Health. 2019; 16(9):1663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16091663
Chicago/Turabian StyleYuan, Jihong, Ping Wang, and Yunfei Yang. 2019. "Effects of Simulated Herbivory on the Vegetative Reproduction and Compensatory Growth of Hordeum brevisubulatum at Different Ontogenic Stages" International Journal of Environmental Research and Public Health 16, no. 9: 1663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16091663