Pre-Implantation Mouse Embryos Cultured In Vitro under Different Oxygen Concentrations Show Altered Ultrastructures

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Embryo Collection, In Vitro Fertilization and Embryo Culture
2.4. Light Microscopy (LM) and Transmission Electron Microscopy (TEM)
3. Results
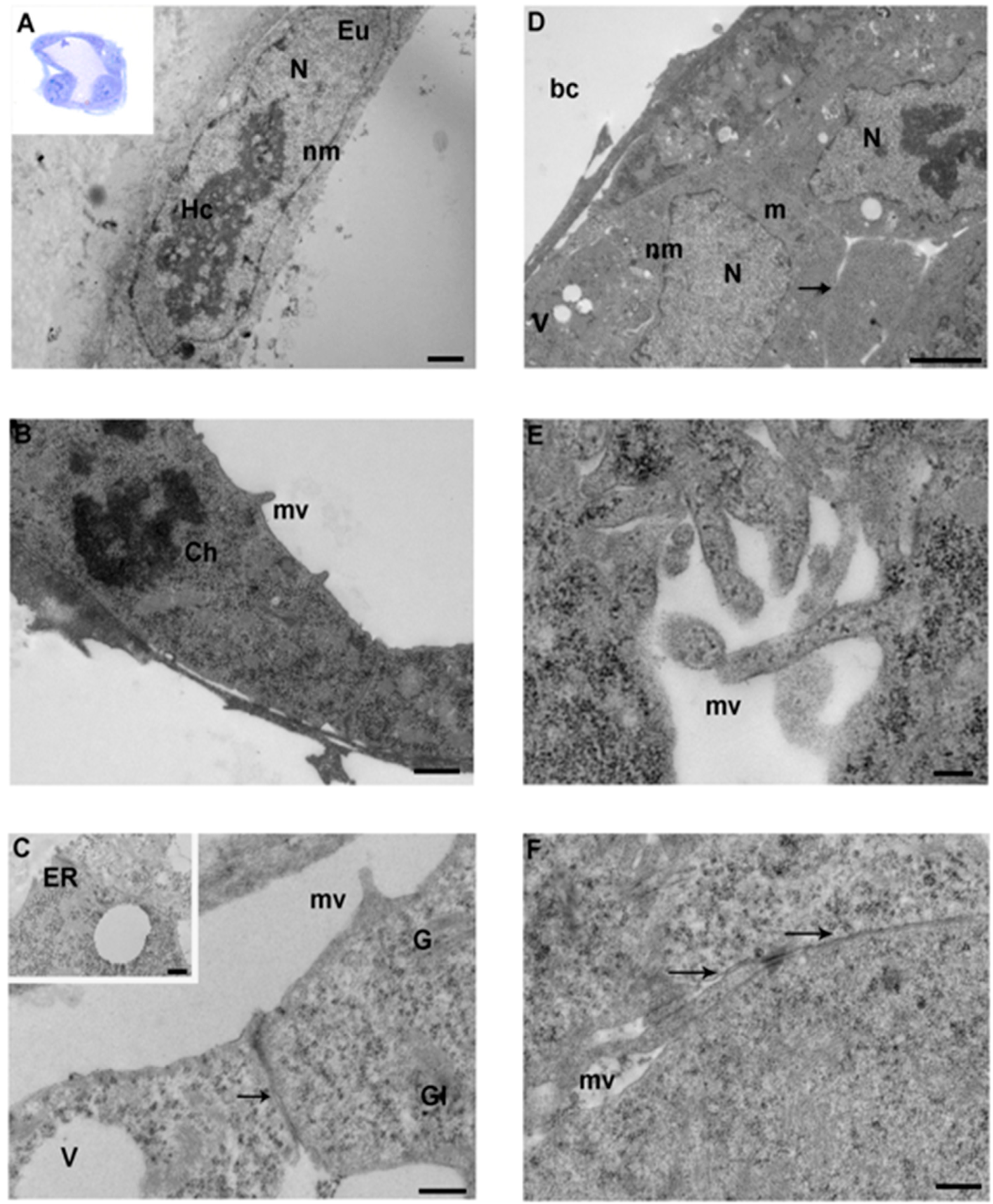
3.1. 2-Cell Stage
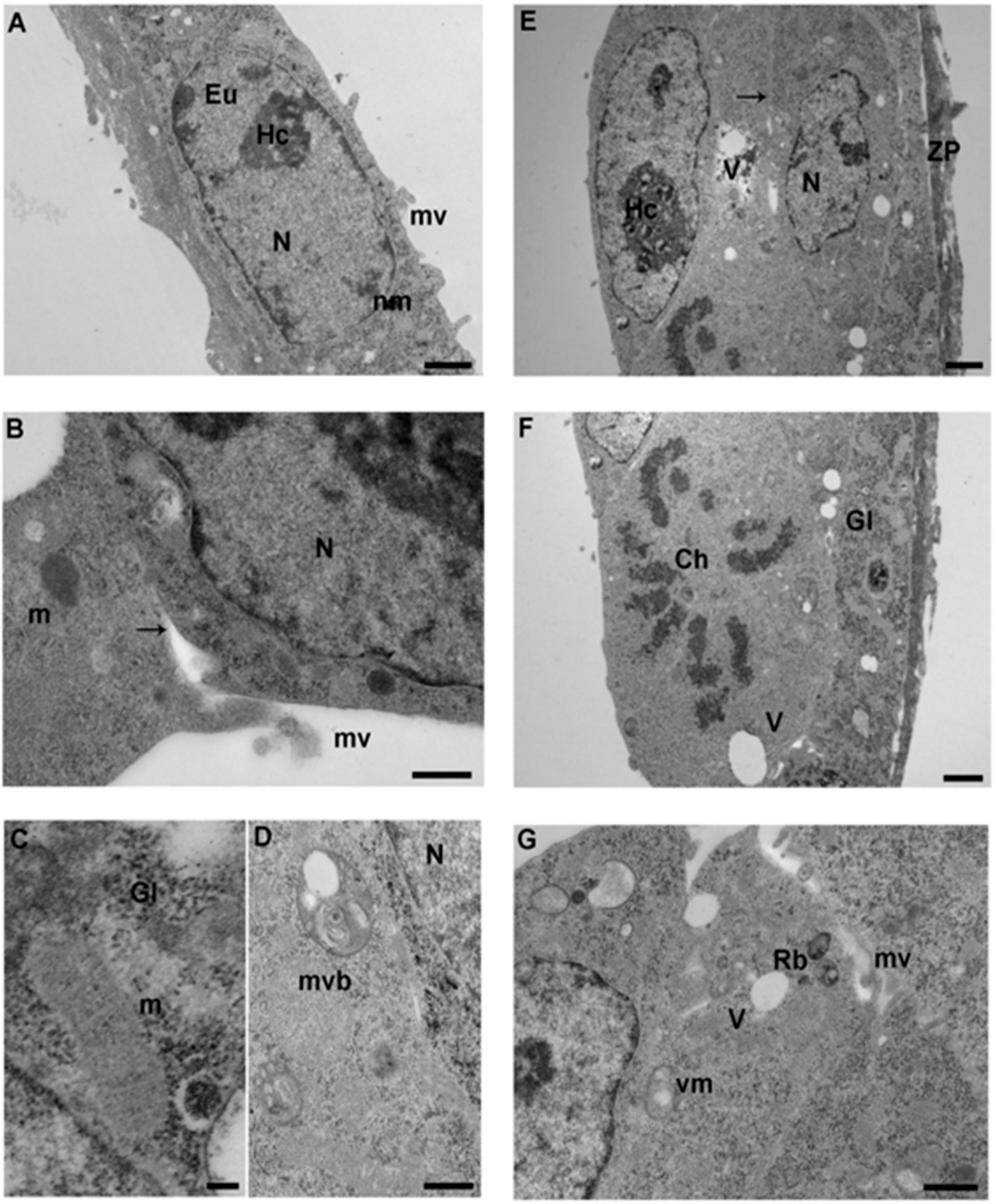
3.2. 4-Cell Stage
3.3. Morula
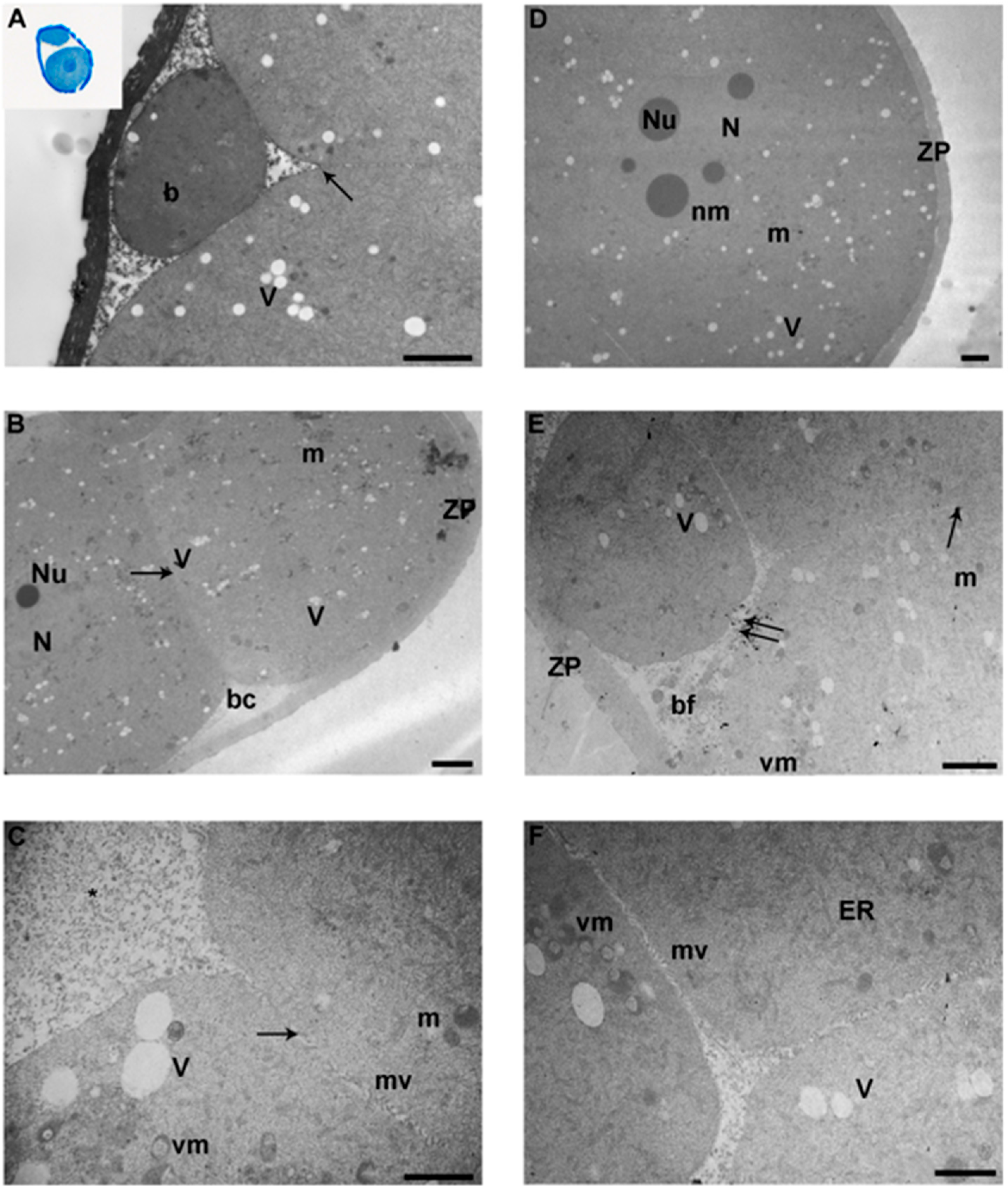
3.4. Blastocyst
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Delle Piane, L.; Lin, W.; Liu, X.; Donjacour, A.; Minasi, P.; Revelli, A.; Maltepe, E.; Rinaudo, P.F. Effect of the method of conception and embryo transfer procedure on mid-gestation placenta and fetal development in an IVF mouse model. Hum. Reprod. 2010, 25, 2039–2046. [Google Scholar] [CrossRef] [Green Version]
- Donjacour, A.; Liu, X.; Lin, W.; Simbulan, R.; Rinaudo, P.F. In vitro fertilization affects growth and glucose metabolism in a sex-specific manner in an outbred mouse model. Biol. Reprod. 2014, 90, 80. [Google Scholar] [CrossRef]
- Feuer, S.K.; Donjacour, A.; Simbulan, R.K.; Lin, W.; Liu, X.; Maltepe, E.; Rinaudo, P.F. Sexually Dimorphic Effect of In Vitro Fertilization (IVF) on Adult Mouse Fat and Liver Metabolomes. Endocrinology 2014, 55, 4554–4567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christianson, M.S.; Zhao, Y.; Shoham, G.; Granot, I.; Safran, A.; Khafagy, A.; Leong, M.; Shoham, Z. Embryo catheter loading and embryo culture techniques: Results of a worldwide Web-based survey. J. Assist. Reprod. Genet. 2014, 31, 1029–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belli, M.; Zhang, L.; Liu, X.; Donjacour, A.; Ruggeri, E.; Palmerini, M.G.; Nottola, S.A.; Macchiarelli, G.; Rinaudo, P. Oxygen concentration alters mitochondrial structure and function in in vitro fertilized preimplantation mouse embryos. Hum. Reprod. 2019, 34, 601–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, B.; Bavister, B.D. Oxygen tension in the oviduct and uterus of rhesus monkeys, hamsters and rabbits. J. Reprod. Fertil. 1993, 99, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Yedwab, G.A.; Paz, G.; Homonnai, T.Z.; David, M.P.; Kraicer, P.F. The temperature, pH, and partial pressure of oxygen in the cervix and uterus of women and uterus of rats during the cycle. Fertil. Steril. 1976, 27, 304–309. [Google Scholar] [CrossRef]
- Mills, R.M.; Brinster, R.L. Oxygen consumption of preimplantation mouse embryos. Exp. Cell. Res. 1967, 47, 337–344. [Google Scholar] [CrossRef]
- Byatt-Smith, J.G.; Leese, H.J.; Gosden, R.G. An investigation by mathematical modelling of whether mouse and human preimplantation embryos in static culture can satisfy their demands for oxygen by diffusion. Hum. Reprod. 1991, 6, 52–57. [Google Scholar] [CrossRef]
- Adam, A.A.G.; Takahashi, Y.; Katagiri, S.; Nagano, M. In vitro culture of mouse preantral follicles using membrane inserts and developmental competence of in vitro ovulated oocytes. J. Reprod. Dev. 2004, 50, 579–586. [Google Scholar] [CrossRef] [Green Version]
- Preis, K.A.; Seidel, G.E.; Gardner, D.K. Reduced oxygen concentration improves the developmental competence of mouse oocytes following in vitro maturation. Mol. Reprod. Dev. 2007, 74, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Chen, H.W.; Tzeng, C.R. Low oxygen tension increases mitochondrial membrane potential and enhances expression of antioxidant genes and implantation protein of mouse blastocyst cultured in vitro. J. Ovarian Res. 2017, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Kasterstein, E.; Strassburger, D.; Komarovsky, D.; Bern, O.; Komsky, A.; Raziel, A.; Friedler, S.; Ron-El, R. The effect of two distinct levels of oxygen concentration on embryo development in a sibling oocyte study. J. Assist. Reprod. Genet. 2013, 30, 1073–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, S.J. Oxygen tension in embryo culture: Does a shift to 2% O2 in extended culture represent the most physiologic system? J. Assist. Reprod. Genet. 2017, 34, 309–314. [Google Scholar] [CrossRef]
- Waldenström, U.; Engström, A.B.; Hellberg, D.; Nilsson, S. Low-oxygen compared with high-oxygen atmosphere in blastocyst culture, a prospective randomized study. Fertil. Steril. 2009, 91, 2461–2465. [Google Scholar] [CrossRef]
- Gelo, N.; Kirinec, G.; Baldani, D.P.; Vrčić, H.; Ježek, D.; Milošević, M.; Stanić, P. Influence of human embryo cultivation in a classic CO2 incubator with 20% oxygen versus benchtop incubator with 5% oxygen on live births: The randomized prospective trial. Zygote 2019, 27, 131–136. [Google Scholar] [CrossRef]
- Nottola, S.A.; Albani, E.; Coticchio, G.; Palmerini, M.G.; Lorenzo, C.; Scaravelli, G.; Borini, A.; Levi-Setti, P.E.; Macchiarelli, G. Freeze/thaw stress induces organelle remodeling and membrane recycling in cryopreserved human mature oocytes. J. Assist. Reprod. Genet. 2016, 33, 1559–1570. [Google Scholar] [CrossRef]
- Ebner, T.; Moser, M.; Sommergruber, M.; Gaiswinkler, U.; Shebl, O.; Jesacher, K.; Tews, G. Occurrence and developmental consequences of vacuoles throughout preimplantation development. Fertil. Steril. 2005, 83, 1635–1640. [Google Scholar] [CrossRef]
- Belli, M.; Antonouli, S.; Palmerini, M.G.; Bianchi, S.; Bernardi, S.; Khalili, M.A.; Donfrancesco, O.; Nottola, S.A.; Macchiarelli, G. The effect of low and ultra-low oxygen tensions on mammalian embryo culture and development in experimental and clinical IVF. Syst. Biol. Reprod. Med. 2020, in press. [Google Scholar] [CrossRef]
- Maheshwari, A.; Hamilton, M.; Bhattacharya, S. Should we be promoting embryo transfer at blastocyst stage? Reprod. Biomed. Online. 2016, 32, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Ma, B.; Mo, S.; Ma, L.; Chang, F.; Zhang, L.; Xu, F.; Wang, L. Improvement of pregnancy outcome by extending embryo culture in IVF-ET during clinical application. J. Assist. Reprod. Genet. 2018, 35, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Feuer, S.K.; Liu, X.; Donjacour, A.; Lin, W.; Simbulan, R.K.; Giritharan, G.; Piane, L.D.; Kolahi, K.; Ameri, K.; Maltepe, E.; et al. Use of a mouse in vitro fertilization model to understand the developmental origins of health and disease hypothesis. Endocrinology 2014, 155, 1956–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macchiarelli, G.; Nottola, S.A.; Palmerini, M.G.; Bianchi, S.; Maione, M.; Lorenzo, C.; Stifano, G.; Di Marco, E.; Correr, S. Morphological expression of angiogenesis in the mammalian ovary as seen by SEM of corrosion casts. Ital. J. Anat. Embryol. 2010, 115, 109–114. [Google Scholar] [PubMed]
- Bernardi, S.; Bianchi, S.; Botticelli, G.; Rastelli, E.; Tomei, A.R.; Palmerini, M.G.; Continenza, M.A.; Macchiarelli, G. Scanning electron microscopy and microbiological approaches for the evaluation of salivary microorganisms behaviour on anatase titanium surfaces: In vitro study. Morphologie 2018, 102, 1–6. [Google Scholar] [CrossRef]
- Bernardi, S.; Bianchi, S.; Tomei, A.R.; Continenza, M.A.; Macchiarelli, G. Microbiological and SEM-EDS Evaluation of Titanium Surfaces Exposed to Periodontal Gel: In Vitro Study. Materials 2019, 12, 1448. [Google Scholar] [CrossRef] [Green Version]
- Palmerini, M.G.; Belli, M.; Nottola, S.A.; Miglietta, S.; Bianchi, S.; Bernardi, S.; Antonouli, S.; Cecconi, S.; Familiari, G.; Macchiarelli, G. Mancozeb impairs the ultrastructure of mouse granulosa cells in a dose-dependent manner. J. Reprod. Dev. 2018, 64, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Nottola, S.A.; Cecconi, S.; Bianchi, S.; Motta, C.; Rossi, G.; Continenza, M.A.; Macchiarelli, G. Ultrastructure of isolated mouse ovarian follicles cultured in vitro. Reprod. Biol. Endocrinol. 2011, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, S.; Macchiarelli, G.; Micara, G.; Linari, A.; Boninsegna, C.; Aragona, C.; Rossi, G.; Cecconi, S.; Nottola, S.A. Ultrastructural markers of quality are impaired in human metaphase II aged oocytes: A comparison between reproductive and in vitro aging. J. Assist. Reprod. Genet. 2015, 32, 1343–1358. [Google Scholar] [CrossRef] [Green Version]
- Palmerini, M.G.; Zhurabekova, G.; Balmagambetova, A.; Nottola, S.A.; Miglietta, S.; Belli, M.; Bianchi, S.; Cecconi, S.; Di Nisio, V.; Familiari, G.; et al. The pesticide Lindane induces dose-dependent damage to granulosa cells in an in vitro culture. Reprod. Biol. 2017, 17, 349–356. [Google Scholar] [CrossRef]
- Gardner, D.K.; Lane, M.; Calderon, I.; Leeton, J. Environment of the preimplantation human embryo in vivo: Metabolite analysis of oviduct and uterine fluids and metabolism of cumulus cells. Fertil. Steril. 1996, 65, 349–353. [Google Scholar] [CrossRef]
- Rinaudo, P.F.; Giritharan, G.; Talbi, S.; Dobson, A.T.; Schultz, R.M. Effects of oxygen tension on gene expression in preimplantation mouse embryos. Fertil. Steril. 2006, 86, 1252–1265. [Google Scholar] [CrossRef] [PubMed]
- McKiernan, S.H.; Bavister, B.D. Environmental variables influencing in vitro development of hamster 2-cell embryos to the blastocyst stage. Biol. Reprod. 1990, 43, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Foote, R.H. Culture of rabbit zygotes into blastocysts in protein-free medium with one to twenty per cent oxygen. J. Reprod. Fertil. 1993, 98, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, F.; Terqui, M. Effects of oxygen, CO2/pH and medium on the in vitro development of individually cultured porcine one- and twocell embryos. Reprod. Nutr. Dev. 1996, 36, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Thompson, T.G.; Simpson, A.C.; Pugh, P.A.; Donnelly, P.E.; Tervit, H.R. Effect of oxygen concentration on in vitro development of preimplantation sheep and cattle embryos. J. Reprod. Fertil. 1990, 89, 573–578. [Google Scholar] [CrossRef] [Green Version]
- De Munck, N.; Janssens, R.; Segers, I.; Tournaye, H.; Van de Velde, H.; Verheyen, G. Influence of ultra-low oxygen (2%) tension on in-vitro human embryo development. Hum. Reprod. 2019, 34, 228–234. [Google Scholar] [CrossRef]
- Feuer, S.; Liu, X.; Donjacour, A.; Simbulan, R.; Maltepe, E.; Rinaudo, P. Common and specific transcriptional signatures in mouse embryos and adult tissues induced by in vitro procedures. Reproduction 2016, 153, 107–122. [Google Scholar] [CrossRef] [Green Version]
- Khalili, M.A.; Maione, M.; Palmerini, M.G.; Bianchi, S.; Macchiarelli, G.; Nottola, S.A. Ultrastructure of human mature oocytes after vitrification. Eur. J. Histochem. 2012, 56, 38. [Google Scholar] [CrossRef]
- Bianchi, S.; Macchiarelli, G.; Micara, G.; Aragona, C.; Maione, M.; Nottola, S.A. Ultrastructural and morphometric evaluation of aged cumulus-oocyte-complexes. Ital. J. Anat. Embryol. 2013, 118, 28. [Google Scholar]
- Bianchi, S.; Nottola, S.; Torge, D.; Palmerini, M.; Necozione, S.; Macchiarelli, G. Association between Female Reproductive Health and Mancozeb: Systematic Review of Experimental Models. Int. J. Environ. Res. Public Health 2020, 17, 2580. [Google Scholar] [CrossRef] [Green Version]
- Palmerini, M.G.; Nottola, S.A.; Tunjung, W.A.; Kadowaki, A.; Bianchi, S.; Cecconi, S.; Sato, E.; Macchiarelli, G. EGF-FSH supplementation reduces apoptosis of pig granulosa cells in co-culture with cumulus-oocyte complexes. Biochem. Biophys. Res. Commun. 2016, 481, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Zhurabekova, G.; Balmagambetova, A.; Bianchi, S.; Belli, M.; Bekmukhambetov, Y.; Macchiarelli, G. The toxicity of lindane in the female reproductive system: A review on the aral sea. Euromediterranean Biomed. J. 2018, 13, 104–108. [Google Scholar]
- Giusti, I.; Bianchi, S.; Nottola, S.A.; Macchiarelli, G.; Dolo, V. Clinical electron microscopy in the study of human ovarian tissues. EuroMediterranean Biomed. J. 2019, 14, 145–151. [Google Scholar]
- Bianchi, S.; Fantozzi, G.; Bernardi, S.; Antonouli, S.; Continenza, M.A.; Macchiarelli, G. Commercial oral hygiene products and implant collar surfaces: Scanning electron microscopy observations. Can. J. Dent. Hyg. 2020, 54, 26–31. [Google Scholar]
- Bianchi, S.; Bernardi, S.; Continenza, M.A.; Vincenti, E.; Antonouli, S.; Torge, D.; Macchiarelli, G. Scanning Electron Microscopy Approach for Evaluation of Hair Dyed with Lawsonia inermis Powder: In vitro Study. Int. J. Morphol. 2020, 38, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Ducibella, T.; Ukena, T.; Karnovsky, M.; Anderson, E. Changes in cell surface and cortical cytoplasmic organization during early embryogenesis in the preimplantation mouse embryo. J. Cell. Biol. 1977, 74, 153–167. [Google Scholar] [CrossRef]
- Dvorák, M.; Stastná, J.; Cech, S.; Trávnik, P.; Horký, D. The differentiation of rat ova during cleavage. Adv. Anat. Embryol. Cell Biol. 1978, 55, 3–131. [Google Scholar]
- Glujovsky, D.; Blake, D.; Farquhar, C.; Bardach, A. Cleavage stage versus blastocyst stage embryo transfer in assisted reproductive technology. Cochrane Database Syst. Rev. 2012, 7, CD002118. [Google Scholar]
- Fernández-Shaw, S.; Cercas, R.; Braña, C.; Villas, C.; Pons, I. Ongoing and cumulative pregnancy rate after cleavage-stage versus blastocyst-stage embryo transfer using vitrification for cryopreservation: Impact of age on the results. J. Assist. Reprod. Genet. 2015, 32, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Dvorák, M. Submicroscopic differentiation of nucleolus during cleavage of rat ovum. Scr. Med. 1974, 47, 497–502. [Google Scholar]
- Izquierdo, L.; Ebensperger, C. Cell membrane regionalization in early mouse embryos as demonstrated by 5’-nucleotidase activity. J. Embryol. Exp. Morphol. 1982, 69, 115–126. [Google Scholar] [PubMed]
- Harvey, A.; Gibson, T.; Lonergan, T.; Brenner, C. Dynamic regulation of mitochondrial function in preimplantation embryos and embryonic stem cells. Mitochondrion 2011, 11, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Belli, M.; Antonouli, S.; Nottola, S.A. Mitochondria in mammalian oocytes and early embryos. A review on morphological and functional studies. EuroMediterranean Biomed. J. 2018, 13, 114–117. [Google Scholar]
- Ding, W.X.; Yin, X.M. Mitophagy: Mechanisms, pathophysiological roles, and analysis. Biol. Chem. 2012, 393, 547–564. [Google Scholar] [CrossRef] [Green Version]
- Sathananthan, A.H.; Ng, S.C.; Bongso, A.; Trounson, A.; Ratnam, S.S. Visual Atlas of Early Human Development for Assisted Reproductive Technology; National University of Singapore: Singapore, 1993; p. 209. [Google Scholar]
- El Shafie, M.; Sousa, M.; Windt, M.L.; Kruger, T.F. An Atlas of the Ultrastructure of Human Oocytes; Parthenon Publishing: New York, NY, USA, 2000; p. 187. [Google Scholar]
- Van blerkom, J. Mitochondria in early mammalian development. Semin. Cell. Dev. Biol. 2009, 20, 354–364. [Google Scholar] [CrossRef]
- Van Blerkom, J. Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion 2011, 11, 797–813. [Google Scholar] [CrossRef]
- Fu, Z.; Wang, B.; Wang, S.; Wu, W.; Wang, Q.; Chen, Y.; Kong, S.; Lu, J.; Tang, Z.; Ran, H.; et al. Integral Proteomic Analysis of Blastocysts Reveals Key Molecular Machinery Governing Embryonic Diapause and Reactivation for Implantation in Mice. Biol. Reprod. 2014, 90, 52. [Google Scholar] [CrossRef]
- Saksouk, N.; Simboeck, E.; Déjardin, J. Constitutive heterochromatin formation and transcription in mammals. Epigenetics Chromatin 2015, 15, 3. [Google Scholar] [CrossRef] [Green Version]
- Reik, W. Stability and flexibility of epigenetic gene regulation in mammalian development. Nature 2007, 447, 425–432. [Google Scholar] [CrossRef]
- Popken, J.; Graf, A.; Krebs, S.; Blum, H.; Schmid, V.J.; Strauss, A.; Guengoer, T.; Zakhartchenko, V.; Wolf, E.; Cremer, T. Remodeling of the Nuclear Envelope and Lamina during Bovine Preimplantation Development and Its Functional Implications. PLoS ONE 2015, 10, e0124619. [Google Scholar] [CrossRef] [Green Version]
- Cech, S.; Sedlackova, M. Ultrastructure and morphometric an analysis of preimplantation mouse embryos. Cell Tissue Res. 1983, 230, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Crosier, A.E.; Farin, P.W.; Dykstra, M.J.; Alexander, J.E.; Farin, C.E. Ultrastructural morphometry of bovine compact morulae produced in vivo or in vitro. Biol. Reprod. 2000, 62, 1459–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thandavan, S.P.; Jiang, M.; Schoenlein, P.; Dong, Z. Autophagy: Molecular machinery, regulation, and implications for renal pathophysiology. Am. J. Physiol. Renal. Physiol. 2009, 297, F244–F256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherz-Shouval, R.; Elazar, Z. ROS, mitochondria and the regulation of autophagy. Trends Cell Biol. 2007, 17, 422–427. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control: In Vivo Fertilized Mouse Embryos Flushed from the Uterus after Natural Fertilization | In Vitro Fertilization (IVF) 5%: IVF Performed in Optimal Conditions (Potassium Simplex Optimized Medium-KSOM with Amino Acids and 5% Oxygen) | IVF 20%: IVF Performed in High Oxygen (KSOM with Amino Acids and 20% Oxygen) |
|---|---|---|
| 2-cell: 24–28 h | 2-cell: 38 h | 2-cell: 38-h |
| 4-cell: 36–40 h | 4-cell: 48–50 h | 4-cell: 48–50 h |
| Morula: 60–70 h | Morula: 75–85 h | Morula: 75–85 h |
| Blastocyst: 82–84 h | Blastocyst: 108–110 h | Blastocyst: 108–110 h |
| Blastocyst Control | IVF 5% Blastocyst | IVF 20% Blastocyst | |
|---|---|---|---|
| Nuclear Shape | Oval or irregular shape. The surface showed minor irregularities. | Oval or irregular shape. The surface showed minor irregularities. | Oval, round or irregular shape. The surface presented minor irregularities and rare deep invaginations. |
| Nuclear envelope | Common structure formed by two intact membrane. Perinuclear space is often interrupted by nuclear pores. | N.D. (no differences) | N.D. |
| Hetero Chromatin | Regularly distributed in the nucleus with minor condensation close the nuclear envelope. | N.D. | Lower content of heterochromatin. |
| Nucleoli | One or two per nucleoli with irregular shape and reticular aspect. | N.D. | N.D. |
| Cell membrane | TE cell membrane appeared smooth with few and short microvilli. ICM cell surface presented numerous and longer microvilli. | N.D. | Interruption of the TE or ICM cell membrane were occasionally seen. |
| Mitochondria | Elongated and tubular in shape with long cristae and a medium dense matrix. Vacuolated mitochondria were present. | Higher density of vacuolated mitochondria. | Higher density of vacuolated mitochondria. |
| Vacuoles | Vacuoles were present but with smaller dimension if compared with those of the previous developmental stages. | Higher density of vacuoles. | Higher density of vacuoles. |
| Residual bodies | Occasionally seen in the cytoplasm. | Frequently seen in the cytoplasm. | Frequently seen in the cytoplasm. Smaller than the ones of the previous group. |
| Multi vesicular bodies | No presence of MVB in this group. | Occasionally found in the cytoplasm. | Occasionally found in the cytoplasm. |
| RER | Usually found in form of isolated cisternae. | N.D. | N.D. |
| SER | Present in the cytoplasm in form of small and smooth vesicles. Vesicles organized in cisternae are occasionally seen. | Cytoplasm showed lower density of SER vesicle. | Cytoplasm showed lower density of SER vesicle. |
| Golgi | Occasionally seen in the perinuclear region. | N.D. | N.D. |
| Glycogen | Present in the cytoplasm in mono-particulate form. | Higher density of glycogen granules in the cytoplasm. | Higher density of glycogen granules in the cytoplasm. |
| Lipid droplets | Lipid droplets were present quite regularly in the cytoplasm. | N.D. | N.D. |
| 5% O2 Concentration | 20% O2 Concentration | References | |
|---|---|---|---|
| Mitochondrial numerical density | ~100–200 less mitochondria/area detected. | ~300 less mitochondria/area detected. | Belli et al., 2019 [5] |
| Vacuolization | No differences. | Nearly 200 more vacuoles/area detected. | Belli et al., 2019 [5] |
| mtDNA copy number | ~107 less copied detected. | ~1.5 (107) less copies detected. | Belli et al., 2019 [5] |
| Alteration of the global gene expression | The expression of 264 genes is altered compared to in vivo control. | The expression of 2133 genes is altered compared to in vivo control | Feuer et al., 2017 [37] |
| Cell number | 25 more blastomeres than in vivo control. | 10 less blastomeres than in vivo control. | Rinaudo et al., 2006 [31] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belli, M.; Rinaudo, P.; Palmerini, M.G.; Ruggeri, E.; Antonouli, S.; Nottola, S.A.; Macchiarelli, G. Pre-Implantation Mouse Embryos Cultured In Vitro under Different Oxygen Concentrations Show Altered Ultrastructures. Int. J. Environ. Res. Public Health 2020, 17, 3384. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17103384
Belli M, Rinaudo P, Palmerini MG, Ruggeri E, Antonouli S, Nottola SA, Macchiarelli G. Pre-Implantation Mouse Embryos Cultured In Vitro under Different Oxygen Concentrations Show Altered Ultrastructures. International Journal of Environmental Research and Public Health. 2020; 17(10):3384. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17103384
Chicago/Turabian StyleBelli, Manuel, Paolo Rinaudo, Maria Grazia Palmerini, Elena Ruggeri, Sevastiani Antonouli, Stefania Annarita Nottola, and Guido Macchiarelli. 2020. "Pre-Implantation Mouse Embryos Cultured In Vitro under Different Oxygen Concentrations Show Altered Ultrastructures" International Journal of Environmental Research and Public Health 17, no. 10: 3384. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17103384