Algicidal Efficiency and Genotoxic Effects of Phanerochaete chrysosporium against Microcystis aeruginosa



Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Strains and Cultivation
2.2. Fungal Strains
2.3. Animal and Experimental Design
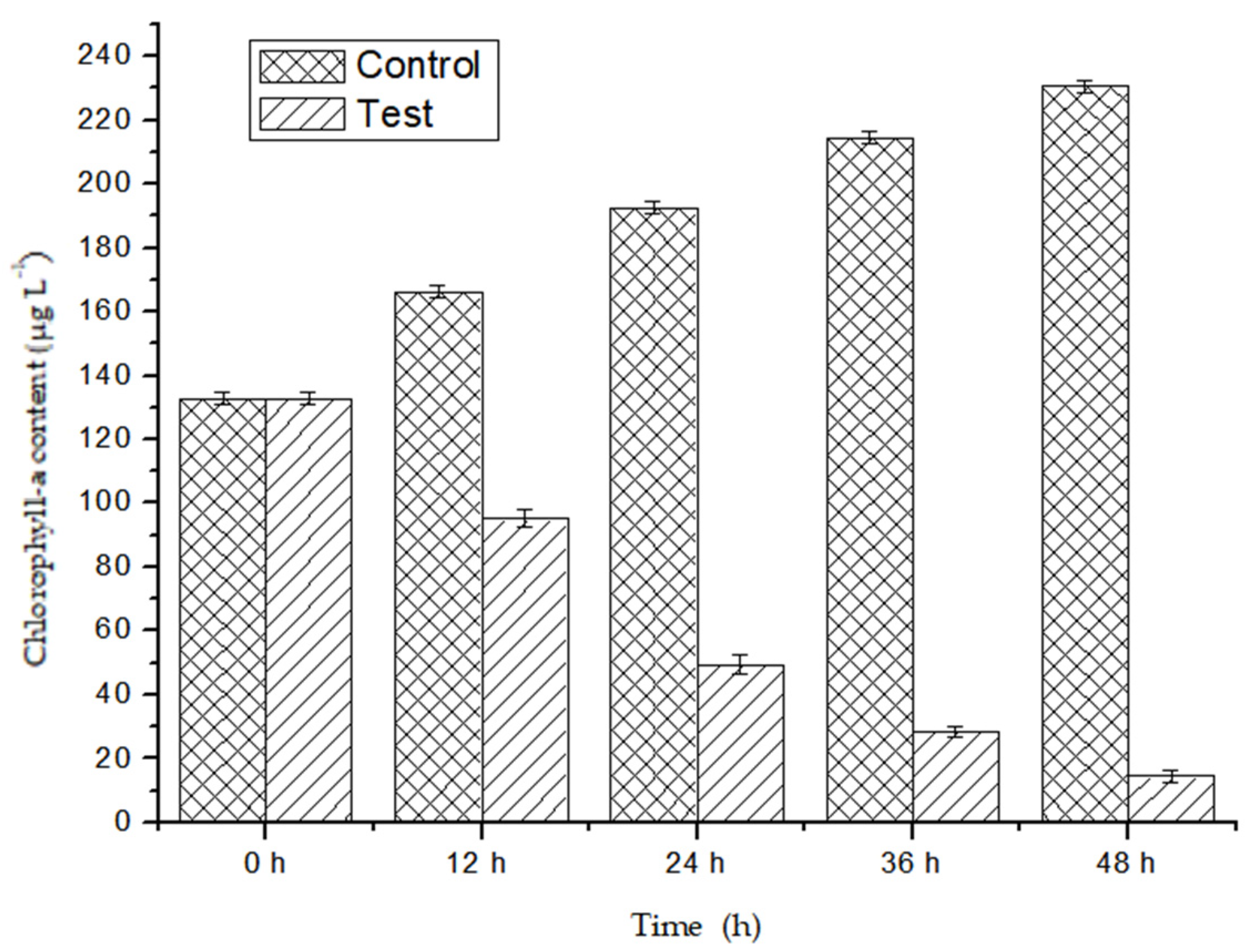
2.4. Chlorophyll—A Content Test
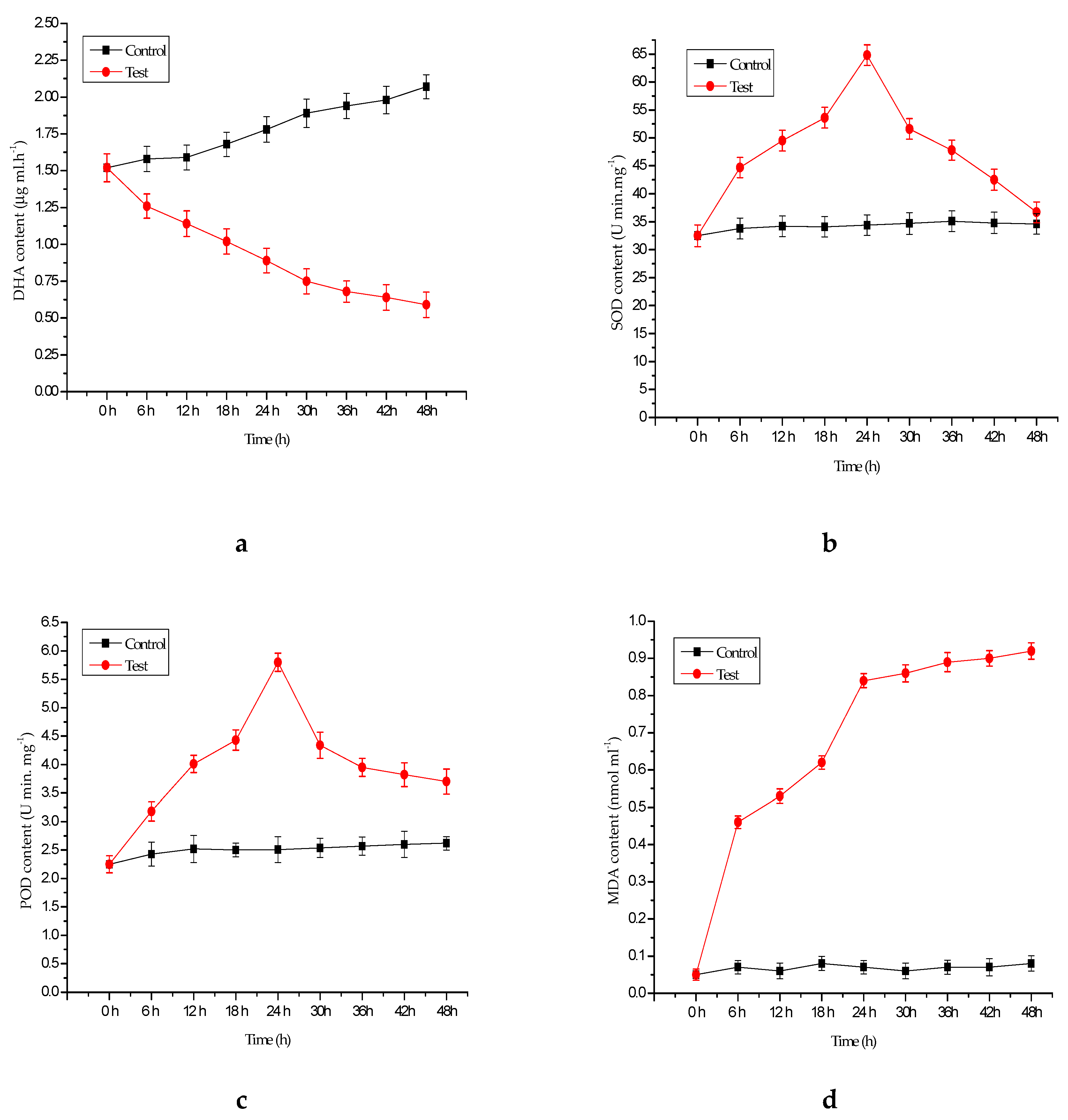
2.5. Antioxidant Enzyme Assays
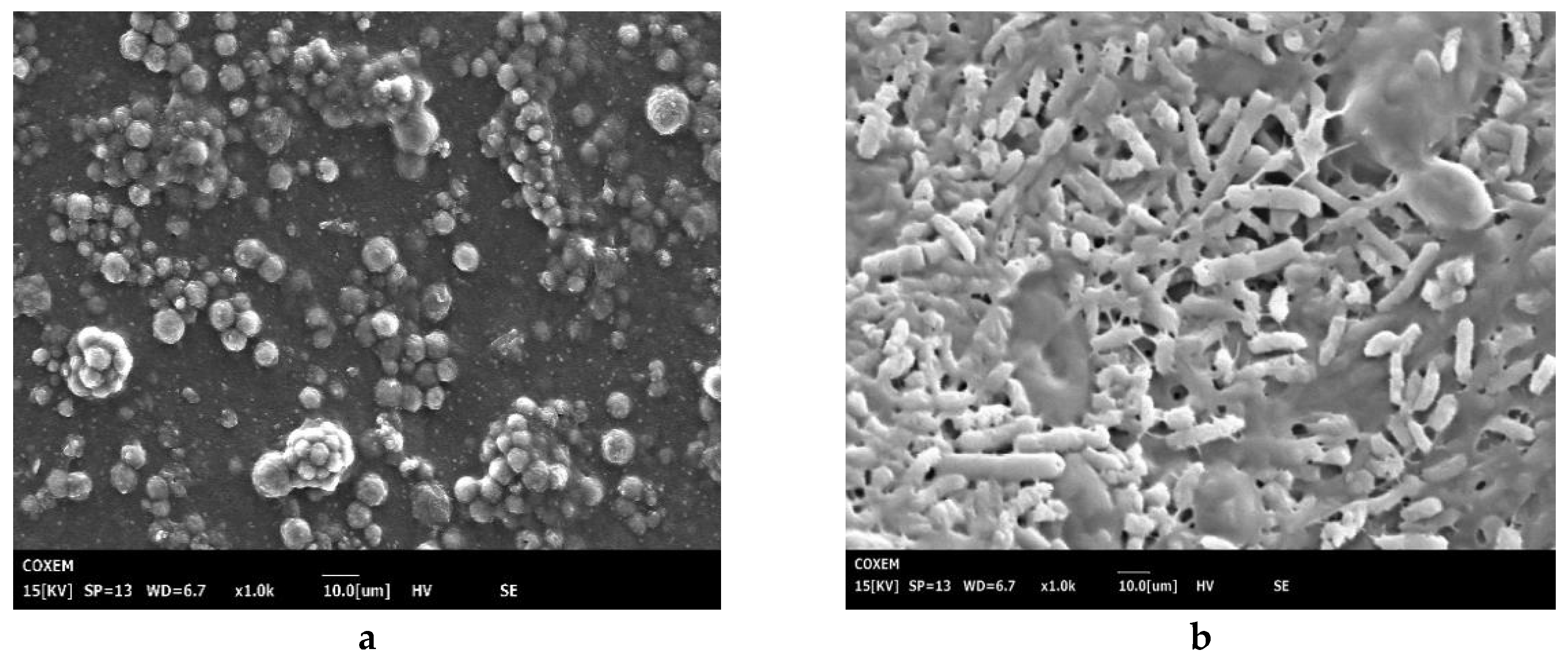
2.6. Scanning Electron Microscopy
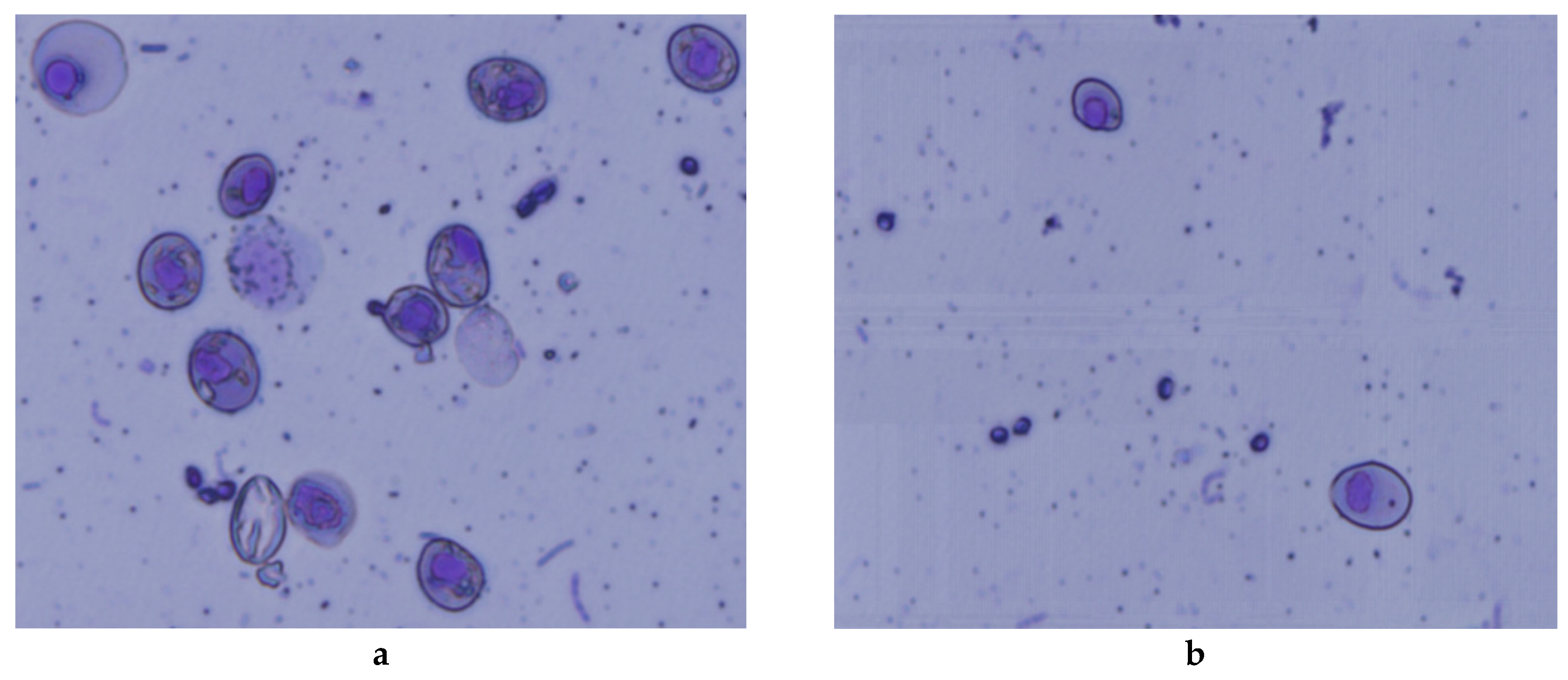
2.7. Micronucleus Assay
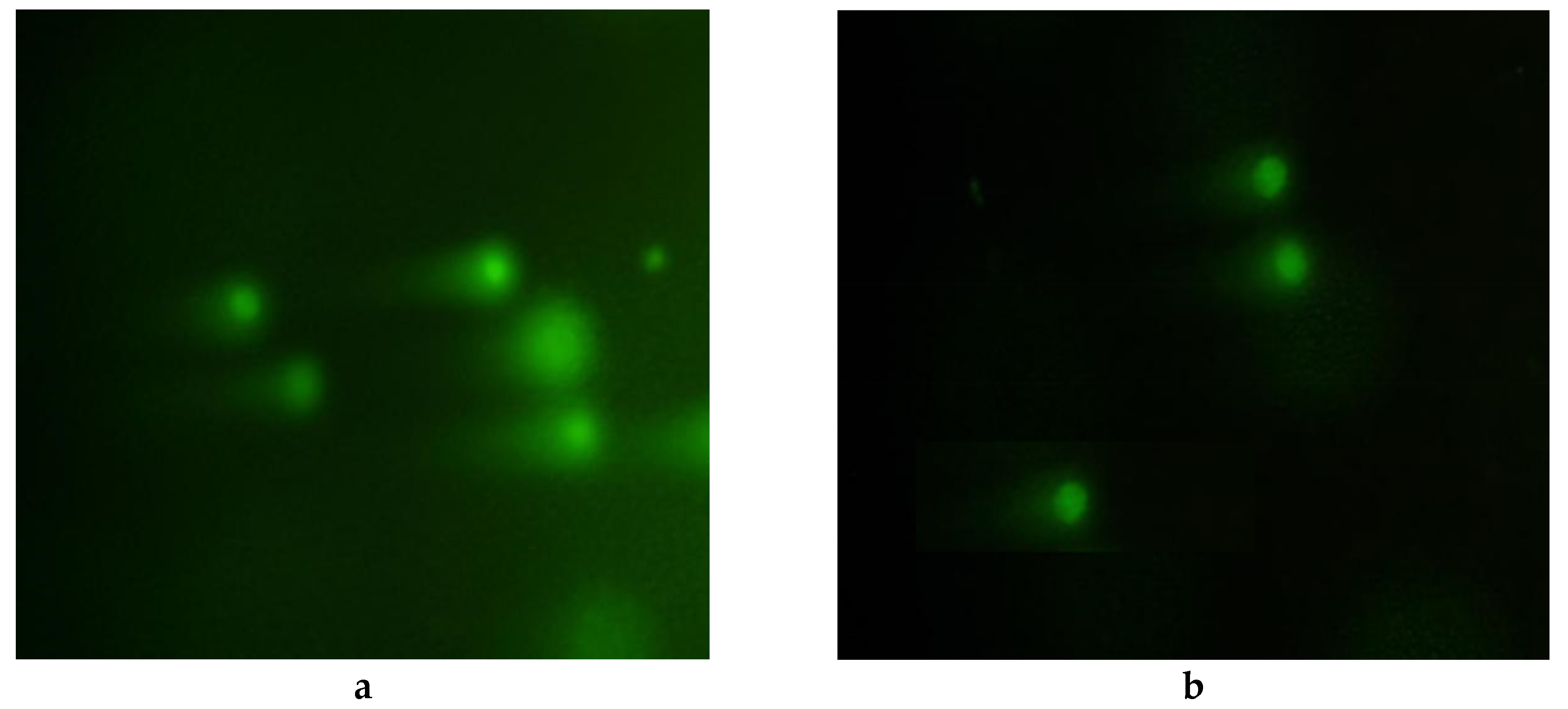
2.8. Comet Assay
2.9. Statistical Analysis
3. Results and Discussion
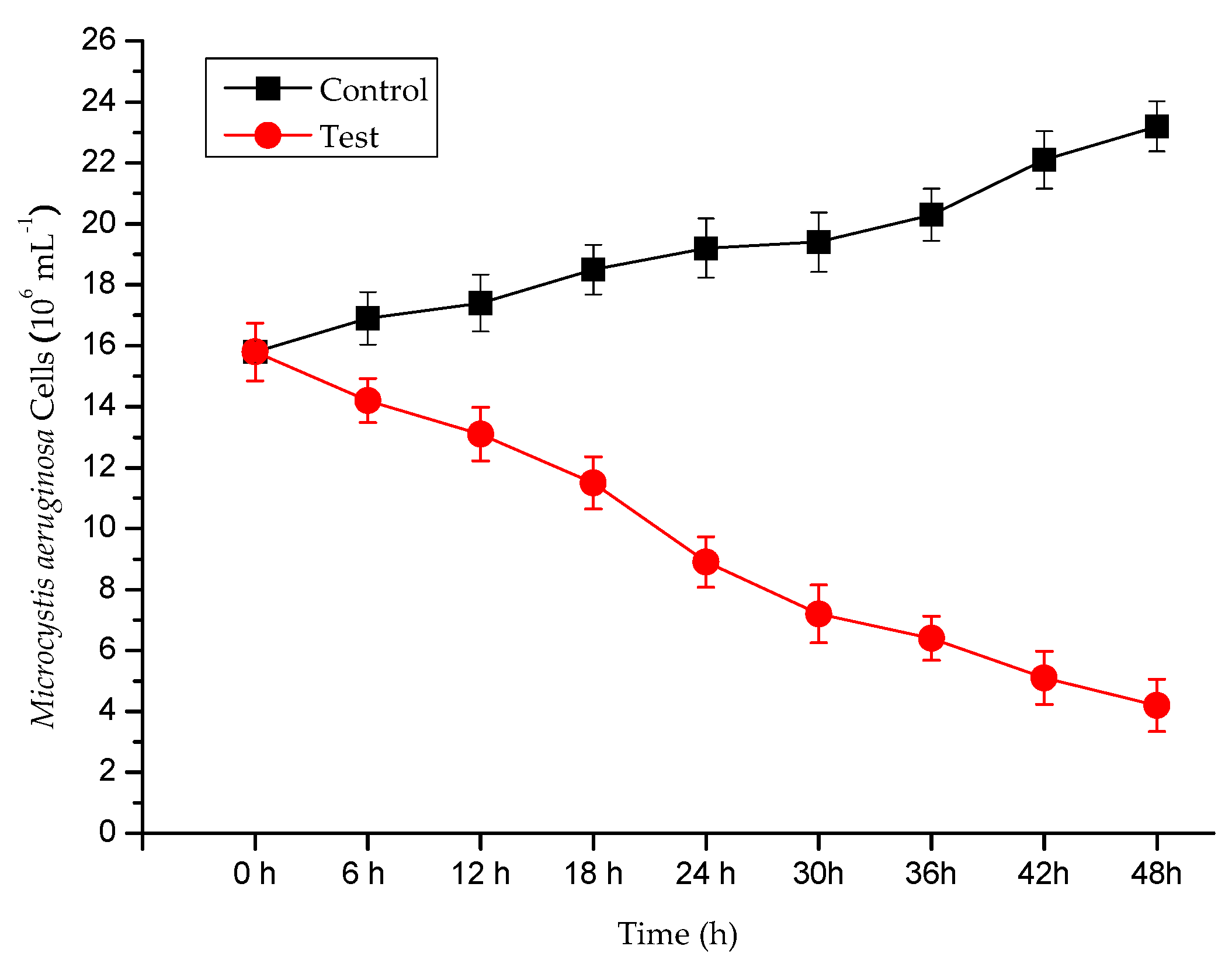
3.1. Algicidal Efficiency
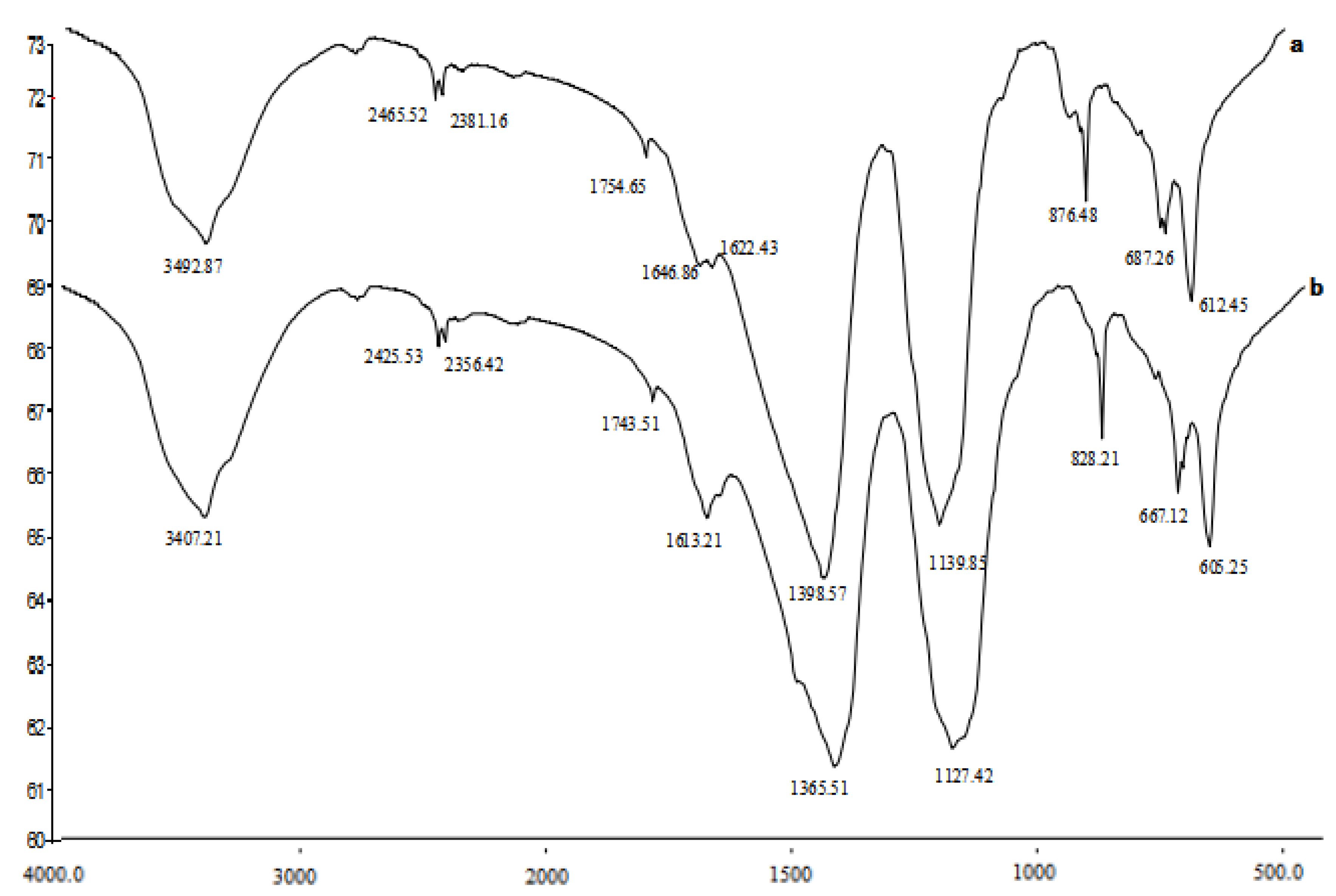
3.2. Fourier Transform Infrared Spectroscopy
3.3. Scanning Electron Microscopy
3.4. Micronuclei Assay
3.5. Comet Assay
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, J.G.; Daniels, V. Algal blooms of the 18th and 19th centuries. Toxicon 2018, 142, 42–44. [Google Scholar] [CrossRef]
- Codd, G.A. Cyanobacterial toxins, the perception of water quality, and the prioritisation of eutrophication control. Ecol. Eng. 2000, 16, 51–60. [Google Scholar] [CrossRef]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful algal blooms and eutrophication: Nutrient sources, composition and consequences. Estuaries 2002, 25, 562–584. [Google Scholar] [CrossRef]
- Dillon, A.; Parry, J.D. Characterization of temperate cyanophages active against freshwater phycocyanin-rich Synechococcus species. Freshw. Biol. 2008, 53, 1253–1261. [Google Scholar] [CrossRef]
- Hare, C.E.; Demir, E.; Coyne, K.J.; Cary, S.C.; Kirchman, D.L.; Hutchins, D.A. A bacterium that inhibits the growth of Pfiesteria piscicida and other dinoflagellates. Harmful Algae 2005, 4, 221–234. [Google Scholar] [CrossRef]
- Dirk, W.; Irene, K.; Spiros, A. White-rot fungi and their enzymes for the treatment of industrial dye effluence. Biotechnol. Adv. 2003, 22, 161–187. [Google Scholar]
- Han, G.M.; Feng, X.G.; Jia, Y.; Wang, C.Y.; He, X.B.; Zhou, Q.Y.; Tian, X.J. Isolation and evaluation of terrestrial fungi with algicidal ability from Zijin Mountain, Nanjing, China. J. Microbiol. 2011, 49, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Wang, Q.; Chen, Z.H.; Jiang, W.X.; Zhang, P.; Tian, X.J. Inhibition of phytoplankton species by co-culture with a fungus. Ecol. Eng. 2010, 36, 1389–1391. [Google Scholar] [CrossRef]
- Zeng, G.M.; Wang, P.; Wang, Y. Algicidal efficiency and mechanism of Phanerochaete chrysosporium against harmful algal bloom species. Algal Res. 2015, 12, 182–190. [Google Scholar] [CrossRef]
- Bai, Y.; Sun, Z.Z.; Zeng, B.; Long, J.Y.; Li, L.; José, V.O.; Li, C. A comparison of dimension reduction techniques for support vector machine modeling of multi-parameter manufacturing quality prediction. J. Intell. Manuf. 2019, 30, 2245–2256. [Google Scholar] [CrossRef]
- Zeng, G.M.; Zhang, M.L.; Wang, P.; Li, X.; Wu, P.; Sun, D. Genotoxicity effects of Phanerochaete chrysosporium against harmful algal bloom species by micronucleus test and comet assay. Chemosphere 2019, 218, 1031–1041. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravind, L.; Walker, D.R.; Koonin, E.V. Conserved domains in DNA repair proteins and evolution of repair systems. Nucleic Acids Res. 1999, 27, 1223–1242. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.Y.; Huang, Y.; Liu, G.X. Using DNA damage to monitor water environment. Chin. J. Oceanol. Limnol. 2005, 23, 340–348. [Google Scholar]
- Frenzilli, G.; Nigro, M.; Lyons, B. The Comet assay for the evaluation of genotoxic impact in aquatic environments. Mutat. Res. Rev. Mutat. Res. 2009, 681, 80–92. [Google Scholar] [CrossRef]
- Ma, F.F.; Carbone, F.; Forti, G.C.; Buschini, A.; Poli, P.; Rossi, C.; Marabini, L.; Radice, S.; Chiesara, E.; Hrelia, P. Drinking water quality: An in vitro approach for the assessment of cytotoxic and genotoxic load in water sampled along distribution system. Environ. Int. 2009, 35, 1053–1061. [Google Scholar]
- Çavas, T. In vivo genotoxicity evaluation of atrazine and atrazine-based herbicide on fish Carassius auratus using the micronucleus test and the comet assay. Food Chem. Toxicol. 2011, 49, 1431–1435. [Google Scholar] [CrossRef]
- Crasta, K.; Ganem, N.J.; Dagher, R.; Lantermann, A.B.; Ivanova, E.V.; Pan, Y.; Nezi, L.; Protopopov, A.; Chowd-hury, D.; Pellman, D. DNA breaks and chromosome pulverization from errors in mitosis. Nature 2012, 482, 53–58. [Google Scholar] [CrossRef]
- Koehler, C.; Ginzkey, C.; Iriehs, C.; Hackenberg, S.; Iroelich, K.; Scherzed, A.; Burghartz, M.; Kessler, M.; Kleinsasser, N. Aspects of nitrogen dioxide toxicity in environmental urban concentrations in hunan nasal epithelium. Toxicol. Appl. Pharm. 2010, 245, 219–225. [Google Scholar] [CrossRef]
- Birder, L.A.; Groat, W.C. Mechanisms of disease: Involvement of the urothelium in bladder dysfunction. Nat. Clin. Pract. Urol. 2007, 4, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Teng, J.; Wang, Z.Y.; Jarrard, D.F.; Bjorling, D.E. Roles of estrogen receptor alpha and beta in modulating urothelial cell proliferation. Endocr. Relat. Cancer 2008, 15, 351–364. [Google Scholar] [CrossRef]
- Lavelle, J.; Meyers, S.; Ramage, R.; Bastacky, S.; Doty, D.; Apodaca, G.; Zeidel, M.L. Bladder permeability barrier: Recovery from selective injury of surface epithelial cells. Am. J. Physiol. Renal Physiol. 2002, 283, F242–F253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, G.M.; Zhou, J.; Huang, T.; Liu, S.Y.; Ji, F.F.; Wang, P. Extraction of Chlorophyll-a from Eutrophic Water by Repeated Freezing and Thawing-Extraction Method. Asian J. Chem. 2014, 26, 2289–2292. [Google Scholar] [CrossRef]
- Xie, J.; Hu, W.R.; Pei, H.Y.; Dun, M.; Qi, F. Detection of amount and activity of living algae in fresh water by dehydrogenase activity (DHA). Environ. Monit. Assess. 2008, 146, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Mitsuta, K. The evaluation theory of free radical scavenging and the application to xanthine oxidase. Bull. Chem. Soc. Jpn. 2010, 83, 351–363. [Google Scholar] [CrossRef]
- Andrews, J.; Malone, M.; Thompson, D.S.; Ho, L.C.; Burton, K.S. Peroxidase isozyme patterns in the skin of maturing tomato fruit. Plant Cell Environ. 2000, 23, 415–422. [Google Scholar] [CrossRef]
- Uchimaya, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Uribe, P.; Espejo, R.T. Effect of associated bacteria on the growth and toxicity of Alexandrium catenella. Appl. Environ. Microbiol. 2003, 69, 659–662. [Google Scholar] [CrossRef] [Green Version]
- Lovejoy, C.; Bowman, J.P.; Hallegraeff, G.M. Algicidal effects of a novel marine Pseudoalteromonas isolate (class Proteobacteria, gamma subdivision) on harmful algal bloom species of the genera Chattonella, Gymnodinium, and Heterosigma. Appl. Environ. Microbiol. 1998, 64, 2806–2813. [Google Scholar] [CrossRef] [Green Version]
- Shedbalkar, U.; Dhanve, R.; Jadhav, J. Biodegradation of triphenylmethane dye cotton blue by Penicillium ochrochloron MTCC 517. J. Hazard. Mater. 2008, 157, 472–479. [Google Scholar] [CrossRef]
- Stoica, B.A.; Bordeianu, G.; Stanescu, R.; Serban, D.N.; Nechifor, M. A new method for the quantification of superoxide dismutase mimics with an allopurinol-xanthine oxidase-lucigenin enhanced system. J. Biol. Inorg. Chem. 2011, 16, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Oncel, L.; Yurdakulol, E.; Keles, Y.; Kurt, L.; Yildiz, A. Role of antioxidant defense system and bio-chemical adaptation on stress tolerance of high mountain and steppe plants. Acta Oecol. 2004, 26, 211–218. [Google Scholar] [CrossRef]
- Shi, S.Y.; Tang, D.S.; Liu, Y.D. Effects of an algicidal bacterium Pseudomonas mendocina on the growth and antioxidant system of Aphanizomenon flos-aquae. Curr. Microbiol. 2009, 59, 107–112. [Google Scholar] [CrossRef]
- Ren, J.; Ma, Q.W.; Huang, H.H. Oxidative degradation of microcystin-LR by combination of UV/H2O2. Fresen. Environ. Bull. 2010, 12, 3037–3044. [Google Scholar]
- Çelik, A.; Kanık, A. Genotoxicity of occupational exposure to wood dust: Micronucleus frequency and nuclear changes in exfoliated buccal mucosa cells. Environ. Mol. Mutagen. 2006, 47, 693–698. [Google Scholar] [CrossRef]
- Djomo, J.E.; Ferrier, V.; Gauthier, L.; Zoll, M.C.; Marty, J. Amphibian micronucleus test in vivo: Evaluation of the genotoxicity of some major polycyclic aromatic hydrocarbons found in crude oil. Mutagenesis 1995, 10, 223–226. [Google Scholar] [CrossRef]
- Lee, R.F.; Steinert, S. Use of the single cell gel electrophoresis/Comet assay for detecting DNA damage in aquatic (marine and freshwater) animals. Mutat. Res. 2003, 544, 43–64. [Google Scholar] [CrossRef]
- Da, S.J.; Freitas, T.R.O.; Marinho, J.R.; Speit, G.; Erdtmann, B. An alkaline single-cell gel eletrophoresis (comet) assay for environmental biomonitoring with native rodents. Genet. Mol. Biol. 2000, 23, 241–245. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | 0 h | 24 h | 48 h | ||||
|---|---|---|---|---|---|---|---|
| Sample | |||||||
| MCN ‰ | ONA ‰ | MCN ‰ | ONA ‰ | MCN ‰ | ONA ‰ | ||
| Control | 1.05 ± 0.09 | 2.45 ± 0.06 | 4.31 ± 0.08 | 10.28 ± 0.10 | 4.52 ± 0.07 | 13.21 ± 0.08 | |
| Test | 1.05 ± 0.09 | 2.45 ± 0.06 | 2.13 ± 0.05 ** | 4.64 ± 0.07 ** | 1.99 ± 0.05 ** | 5.83 ± 0.07 ** | |
| Time (h) | Tail Length (%) | Comet Length (%) | ||
|---|---|---|---|---|
| 0 | 1.12 ± 0.21 a | 1.12 ± 0.21 b | 6.45 ± 0.09 a | 6.45 ± 0.09 b |
| 24 | 18.22 ± 0.41 a | 10.56 ± 0.45 b ** | 37.12 ± 0.24 a | 21.35 ± 0.32 b ** |
| 48 | 36.41 ± 0.65 a | 21.68 ± 0.34 b ** | 52.36 ± 0.86 a | 36.45 ± 0.67 b * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, G.; Zhang, M.; Gao, P.; Wang, J.; Sun, D. Algicidal Efficiency and Genotoxic Effects of Phanerochaete chrysosporium against Microcystis aeruginosa. Int. J. Environ. Res. Public Health 2020, 17, 4029. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17114029
Zeng G, Zhang M, Gao P, Wang J, Sun D. Algicidal Efficiency and Genotoxic Effects of Phanerochaete chrysosporium against Microcystis aeruginosa. International Journal of Environmental Research and Public Health. 2020; 17(11):4029. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17114029
Chicago/Turabian StyleZeng, Guoming, Maolan Zhang, Pei Gao, Jiale Wang, and Da Sun. 2020. "Algicidal Efficiency and Genotoxic Effects of Phanerochaete chrysosporium against Microcystis aeruginosa" International Journal of Environmental Research and Public Health 17, no. 11: 4029. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17114029