Alterations in Serum-Free Amino Acid Profiles in Childhood Asthma

 , , and
, , and
Abstract
:1. Introduction
2. Methods
2.1. Chemicals and Reagents
2.2. Patients
2.3. Sample Collection and Preparation
2.4. LC-MS/MS Instrumentation
2.5. Statistical Analysis
3. Results
3.1. Patients’ Characteristics
3.2. Alterations in Serum-Free AA Profiles in Childhood Asthma
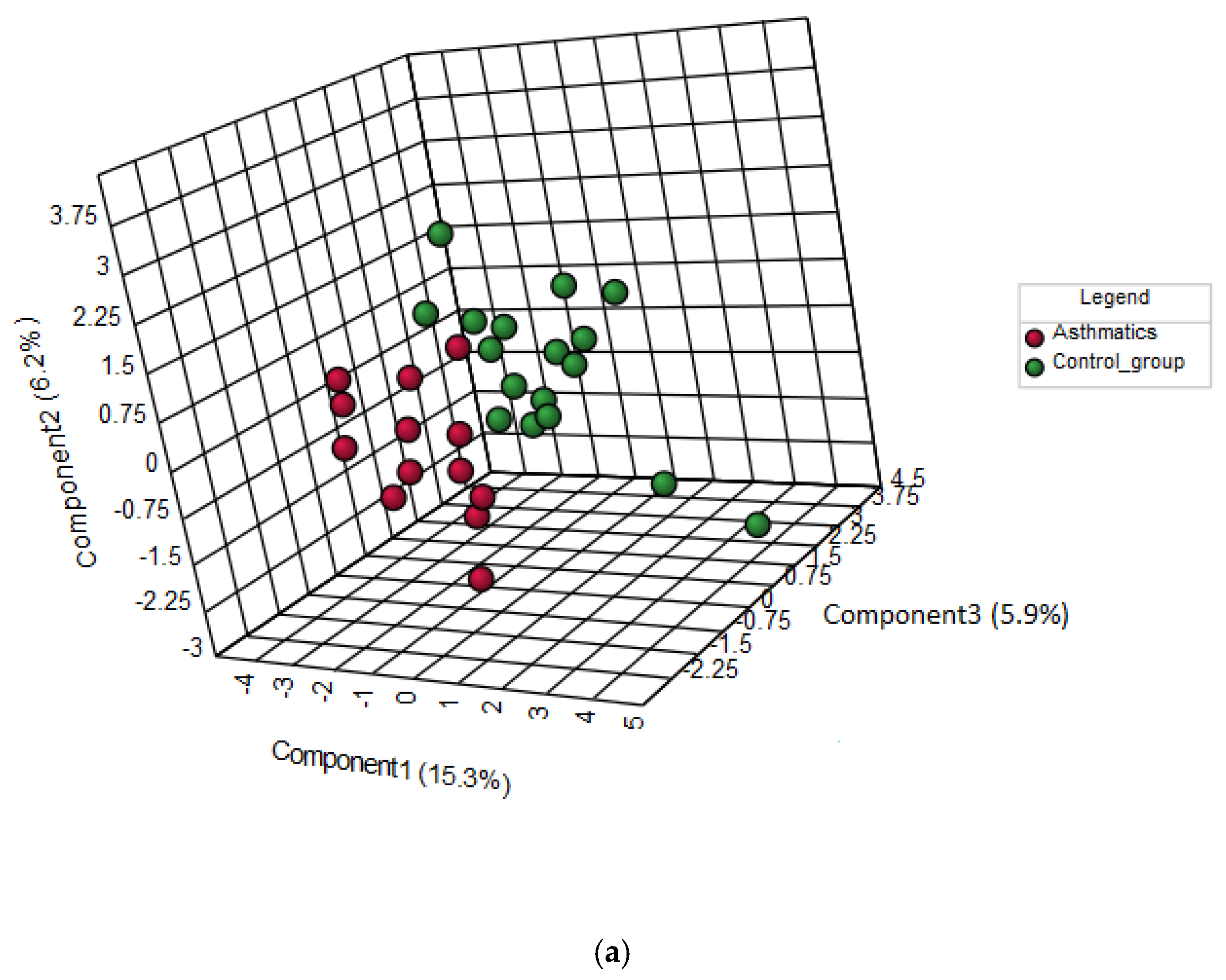
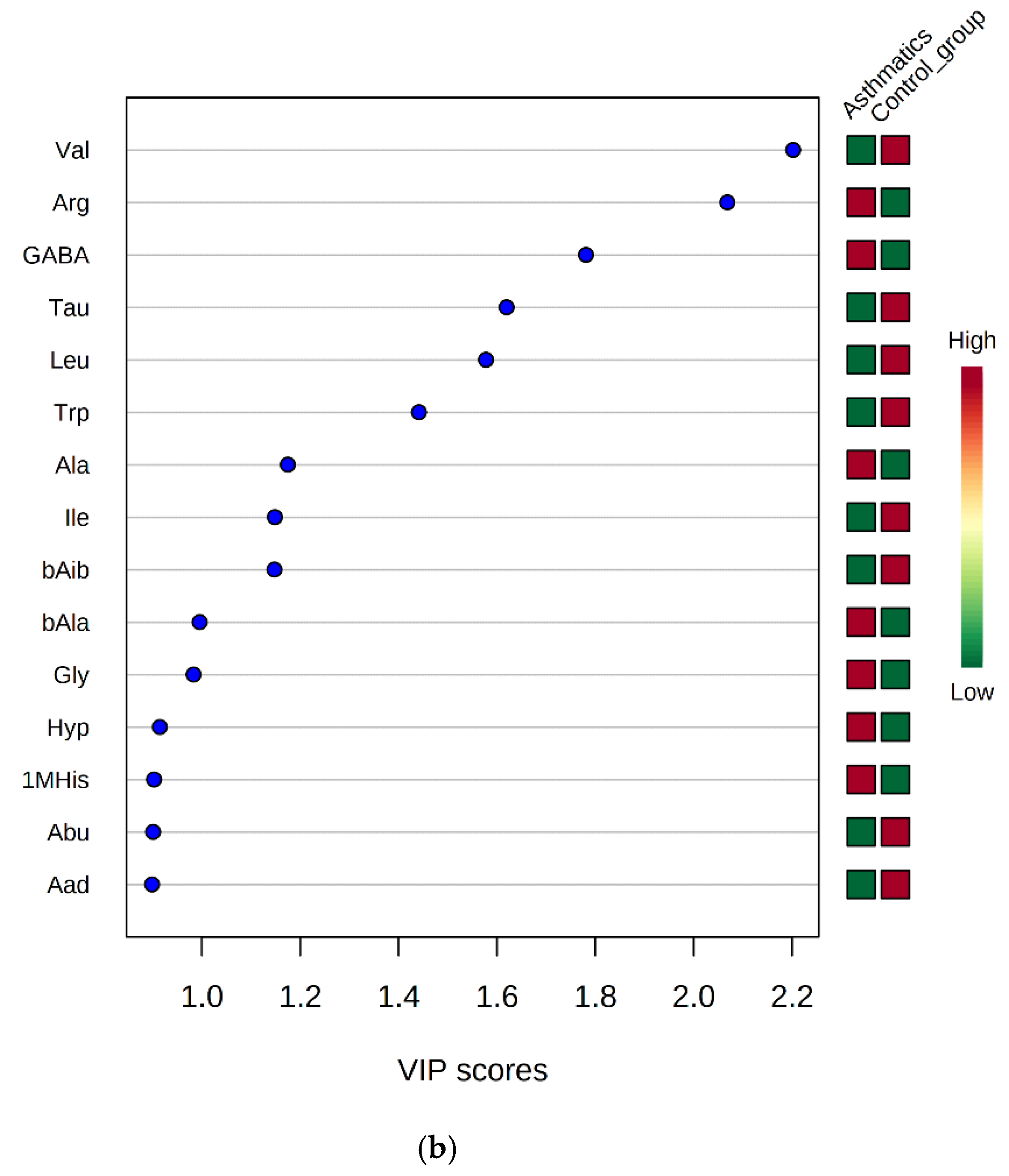
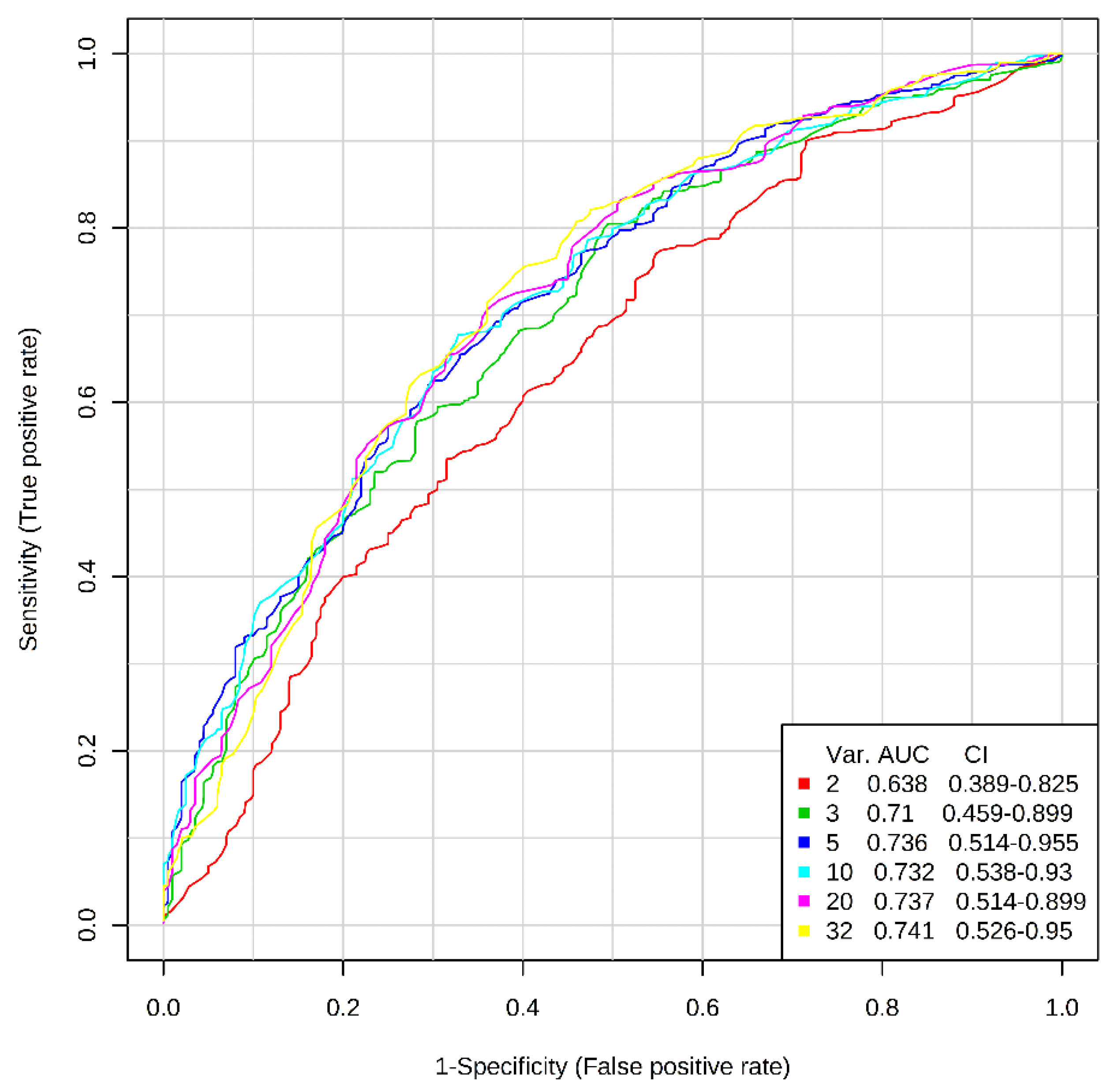
3.3. Discrimination between Asthmatic Children And Healthy Subjects by AA Profiles
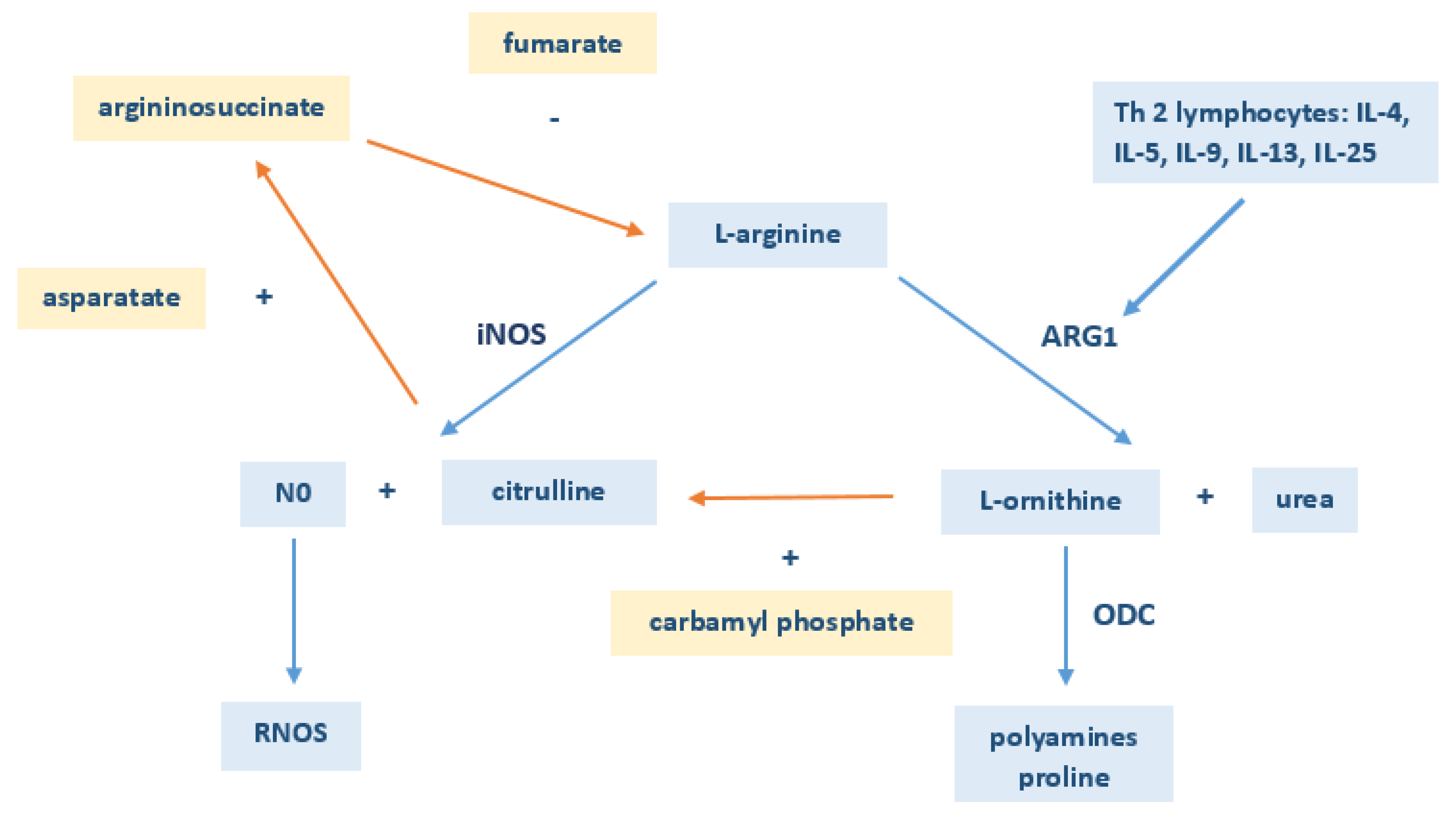
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Russell, R.J.; Brightling, C. Pathogenesis of asthma: Implications for precision medicine. Clin. Sci. (Lond. Engl. 1979) 2017, 131, 1723–1735. [Google Scholar] [CrossRef]
- Epidemiology of Asthma: Current Opinion in Otolaryngology & Head and Neck Surgery. Available online: https://journals.lww.com/co-otolaryngology/Abstract/2016/06000/Epidemiology_of_asthma.14.aspx (accessed on 2 January 2020).
- Adamko, D.J.; Sykes, B.D.; Rowe, B.H. The metabolomics of asthma: Novel diagnostic potential. Chest 2012, 141, 1295–1302. [Google Scholar] [CrossRef]
- Adamko, D.J.; Nair, P.; Mayers, I.; Tsuyuki, R.T.; Regush, S.; Rowe, B.H. Metabolomic profiling of asthma and chronic obstructive pulmonary disease: A pilot study differentiating diseases. J. Allergy Clin. Immunol. 2015, 136, 571–580. [Google Scholar] [CrossRef]
- Drexler, D.M.; Reily, M.D.; Shipkova, P.A. Advances in mass spectrometry applied to pharmaceutical metabolomics. Anal. Bioanal. Chem. 2011, 399, 2645–2653. [Google Scholar] [CrossRef]
- Kim, Y.M.; Heyman, H.M. Mass Spectrometry-Based Metabolomics. In Fungal Genomics; de Vries, R.P., Tsang, A., Grigoriev, I.V., Eds.; Part of the Methods in Molecular Biology Methods Book Series; Humana Press: New York, NY, USA, 2018; Volume 1775, pp. 107–118. [Google Scholar]
- Dettmer, K.; Hammock, B. Metabolomics—A new exciting field within the “omics” sciences. Environ. Health Perspect. 2004, 112, A396–A397. [Google Scholar] [CrossRef] [Green Version]
- Schuhmacher, R.; Krska, R.; Weckwerth, W.; Goodacre, R. Metabolomics and metabolite profiling. Anal. Bioanal. Chem. 2013, 405, 5003–5004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Charron, C.S.; Novotny, J.A.; Peng, B.; Yu, L.; Chen, P. Profiling glucosinolate metabolites in human urine and plasma after broccoli consumption using non-targeted and targeted metabolomic analyses. Food Chem. 2020, 309, 125660. [Google Scholar] [CrossRef] [PubMed]
- Saude, E.J.; Skappak, C.D.; Regush, S.; Cook, K.; Ben-Zvi, A.; Becker, A.; Moqbel, R.; Sykes, B.D.; Rowe, B.H.; Adamko, D.J. Metabolomic profiling of asthma: Diagnostic utility of urine nuclear magnetic resonance spectroscopy. J. Allergy Clin. Immunol. 2011, 127, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Mattarucchi, E.; Baraldi, E.; Guillou, C. Metabolomics applied to urine samples in childhood asthma; differentiation between asthma phenotypes and identification of relevant metabolites. Biomed. Chromatogr. BMC 2012, 26, 89–94. [Google Scholar] [CrossRef]
- Lara, A.; Khatri, S.B.; Wang, Z.; Comhair, S.A.A.; Xu, W.; Dweik, R.A.; Bodine, M.; Levison, B.S.; Hammel, J.; Bleecker, E.; et al. Alterations of the arginine metabolome in asthma. Am. J. Respir. Crit. Care Med. 2008, 178, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Fogarty, A.; Broadfield, E.; Lewis, S.; Lawson, N.; Britton, J. Amino acids and asthma: A case-control study. Eur. Respir. J. 2004, 23, 565–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, C.R.; Poljakovic, M.; Lavrisha, L.; Machado, L.; Kuypers, F.A.; Morris, S.M. Decreased arginine bioavailability and increased serum arginase activity in asthma. Am. J. Respir. Crit. Care Med. 2004, 170, 148–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.; Kim, S.-H.; Lee, H.-S.; Choi, G.S.; Jung, Y.-S.; Ryu, D.H.; Park, H.-S.; Hwang, G.-S. Serum metabolomics reveals pathways and biomarkers associated with asthma pathogenesis. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2013, 43, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Ried, J.S.; Baurecht, H.; Stückler, F.; Krumsiek, J.; Gieger, C.; Heinrich, J.; Kabesch, M.; Prehn, C.; Peters, A.; Rodriguez, E.; et al. Integrative genetic and metabolite profiling analysis suggests altered phosphatidylcholine metabolism in asthma. Allergy 2013, 68, 629–636. [Google Scholar] [CrossRef]
- Checkley, W.; Deza, M.P.; Klawitter, J.; Romero, K.M.; Klawitter, J.; Pollard, S.L.; Wise, R.A.; Christians, U.; Hansel, N.N. Identifying biomarkers for asthma diagnosis using targeted metabolomics approaches. Respir. Med. 2016, 121, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Carraro, S.; Giordano, G.; Reniero, F.; Carpi, D.; Stocchero, M.; Sterk, P.J.; Baraldi, E. Asthma severity in childhood and metabolomic profiling of breath condensate. Allergy 2013, 68, 110–117. [Google Scholar] [CrossRef]
- Kelly, R.S.; Dahlin, A.; McGeachie, M.J.; Qiu, W.; Sordillo, J.; Wan, E.S.; Wu, A.C.; Lasky-Su, J. Asthma Metabolomics and the Potential for Integrative Omics in Research and the Clinic. Chest 2017, 151, 262–277. [Google Scholar] [CrossRef] [Green Version]
- Turi, K.N.; Romick-Rosendale, L.; Ryckman, K.K.; Hartert, T.V. A review of metabolomics approaches and their application in identifying causal pathways of childhood asthma. J. Allergy Clin. Immunol. 2018, 141, 1191–1201. [Google Scholar] [CrossRef] [Green Version]
- Rodnina, M.V.; Beringer, M.; Wintermeyer, W. How Ribosomes Make Peptide Bonds. Trends Biochem. Sci. 2007, 32, 20–26. [Google Scholar] [CrossRef]
- Curis, E.; Nicolis, I.; Moinard, C.; Osowska, S.; Zerrouk, N.; Bénazeth, S.; Cynober, L. Almost All about Citrulline in Mammals. Amino Acids 2005, 29, 177–205. [Google Scholar] [CrossRef]
- Jin, R.; Banton, S.; Tran, V.T.; Konomi, J.V.; Li, S.; Jones, D.P.; Vos, M.B. Amino Acid Metabolism Is Altered in Adolescents with Nonalcoholic Fatty Liver Disease—An Untargeted, High Resolution Metabolomics Study. J. Pediatr. 2016, 172, 14–19.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadel, F.I.; Elshamaa, M.F.; Essam, R.G.; Elghoroury, E.A.; El-Saeed, G.S.M.; El-Toukhy, S.E.; Ibrahim, M.H. Some Amino Acids Levels: Glutamine, Glutamate, and Homocysteine, in Plasma of Children with Chronic Kidney Disease. Int. J. Biomed. Sci. 2014, 10, 36–42. [Google Scholar] [PubMed]
- Sawy, M.E.; Zaki, M.M.; EL-Hakim, I.Z.; Mowafy, M.E.; Al-Abd, H.S. Serum amino acid abnormalities in pediatric patients with chronic renal failure with and without history of thromboembolic manifestations. Egypt. J. Med. Hum. Genet. 2012, 13, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Klupczyńska, A.; Dereziński, P.; Dyszkiewicz, W.; Pawlak, K.; Kasprzyk, M.; Kokot, Z.J. Evaluation of serum amino acid profiles’ utility in non-small cell lung cancer detection in Polish population. Lung Cancer 2016, 100, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Matysiak, J.; Dereziński, P.; Klupczyńska, A.; Hajduk, J.P.; Swiatly, A.; Plewa, S.; Horała, A.; Jaźwiński, P.; Banach, P.; Nowak-Markwitz, E.; et al. Proteomic and metabolomic strategy of searching for biomarkers of genital cancer diseases using mass spectrometry methods. J. Med. Sci. 2016, 85, 330. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Wang, Z.; Huang, Y.; Duan, Y.; Wang, X. Investigation of salivary free amino acid profile for early diagnosis of breast cancer with ultra performance liquid chromatography-mass spectrometry. Clin. Chim. Acta Int. J. Clin. Chem. 2015, 447, 23–31. [Google Scholar] [CrossRef]
- Hasim, A.; Aili, A.; Maimaiti, A.; Mamtimin, B.; Abudula, A.; Upur, H. Plasma-free amino acid profiling of cervical cancer and cervical intraepithelial neoplasia patients and its application for early detection. Mol. Biol. Rep. 2013, 40, 5853–5859. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, M.; Liu, X.; Sun, H.; Guo, Z.; Tang, X.; Wang, Z.; Li, J.; Li, H.; Sun, W.; et al. Urine Metabolomics for Renal Cell Carcinoma (RCC) Prediction: Tryptophan Metabolism as an Important Pathway in RCC. Front. Oncol. 2019, 9, 663. [Google Scholar] [CrossRef] [Green Version]
- Takashina, C.; Tsujino, I.; Watanabe, T.; Sakaue, S.; Ikeda, D.; Yamada, A.; Sato, T.; Ohira, H.; Otsuka, Y.; Oyama-Manabe, N.; et al. Associations among the plasma amino acid profile, obesity, and glucose metabolism in Japanese adults with normal glucose tolerance. Nutr. Metab. 2016, 13, 5. [Google Scholar] [CrossRef]
- Yamakado, M.; Nagao, K.; Imaizumi, A.; Tani, M.; Toda, A. Plasma Free Amino Acid Profiles Predict Four-Year Risk of Developing Diabetes, Metabolic Syndrome, Dyslipidemia, and Hypertension in Japanese Population. Sci. Rep. 2015, 5, 11919. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, W.; Wang, Y.; Pedram, P.; Cahill, F.; Zhai, G.; Randell, E.; Gulliver, W.; Sun, G. Serum metabolic biomarkers distinguish metabolically healthy peripherally obese from unhealthy centrally obese individuals. Nutr. Metab. 2016, 13, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tumor-Dependent Increase of Serum Amino Acid Levels in Breast Cancer Patients Has Diagnostic Potential and Correlates with Molecular Tumor Subtypes. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC3835137/ (accessed on 25 June 2020).
- Shi, T.; Tang, T.; Qian, K.; Wang, F.; Li, J.; Cao, Y. High-performance liquid chromatographic method for determination of amino acids by precolumn derivatization with 4-chloro-3,5-dinitrobenzotrifluoride. Anal. Chim. Acta 2009, 654, 154–161. [Google Scholar] [CrossRef]
- Deng, C.; Li, N.; Zhang, X. Rapid determination of amino acids in neonatal blood samples based on derivatization with isobutyl chloroformate followed by solid-phase microextraction and gas chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 2558–2564. [Google Scholar] [CrossRef]
- Kraj, L.; Krawiec, M.; Koter, M.; Graboń, W.; Kraj, G.; Chołojczyk, M.; Kulus, M.; Barańczyk-Kuźma, A. Altered L-arginine metabolism in children with controlled asthma. Allergy Asthma Proc. 2014, 35, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Matysiak, J.; Dereziński, P.; Klupczyńska, A.; Matysiak, J.; Kaczmarek, E.; Kokot, Z.J. Effects of a Honeybee Sting on the Serum Free Amino Acid Profile in Humans. PLoS One 2014, 9, e103533. [Google Scholar] [CrossRef] [Green Version]
- Held, P.K.; White, L.; Pasquali, M. Quantitative urine amino acid analysis using liquid chromatography tandem mass spectrometry and aTRAQ reagents. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 2695–2703. [Google Scholar] [CrossRef]
- Filee, R.; Schoos, R.; Boemer, F. Evaluation of Physiological Amino Acids Profiling by Tandem Mass Spectrometry. JIMD Rep. 2014, 13, 119–128. [Google Scholar]
- Hogenboom, J.A.; D’Incecco, P.; Fuselli, F.; Pellegrino, L. Ion-Exchange Chromatographic Method for the Determination of the Free Amino Acid Composition of Cheese and Other Dairy Products: An Inter-Laboratory Validation Study. Food Anal. Methods 2017, 10, 3137–3148. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [Green Version]
- Maarsingh, H.; Zaagsma, J.; Meurs, H. Arginine homeostasis in allergic asthma. Eur. J. Pharmacol. 2008, 585, 375–384. [Google Scholar] [CrossRef]
- Zimmermann, N.; Rothenberg, M.E. The arginine-arginase balance in asthma and lung inflammation. Eur. J. Pharmacol. 2006, 533, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.R. Arginine and asthma. Nestle Nutr. Inst. Workshop Ser. 2013, 77, 1–15. [Google Scholar] [PubMed] [Green Version]
- Kelly, R.S.; McGeachie, M.J.; Lee-Sarwar, K.A.; Kachroo, P.; Chu, S.H.; Virkud, Y.V.; Huang, M.; Litonjua, A.A.; Weiss, S.T.; Lasky-Su, J. Partial Least Squares Discriminant Analysis and Bayesian Networks for Metabolomic Prediction of Childhood Asthma. Metabolites 2018, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, I.H.; Kristensen, D.M.; Holm, J.B.; Mortensen, O.H. Physiological role of taurine—From organism to organelle. Acta Physiol. (Oxf. Engl.) 2015, 213, 191–212. [Google Scholar] [CrossRef]
- Ripps, H.; Shen, W. Review: Taurine: A “very essential” amino acid. Mol. Vis. 2012, 18, 2673–2686. [Google Scholar]
- Mongin, A.A. Potential link between cysteinyl-leukotriene receptors and release of bioactive amino acids in regulation of lung function. Focus on “Volume-sensitive release of organic osmolytes in the human lung epithelial cell line A549: Role of the 5-lipoxygenase”. Am. J. Physiol. Cell Physiol. 2013, 305, C24–C25. [Google Scholar] [CrossRef] [Green Version]
- Gallos, G.; Yim, P.; Chang, S.; Zhang, Y.; Xu, D.; Cook, J.M.; Gerthoffer, W.T.; Emala, C.W. Targeting the restricted α-subunit repertoire of airway smooth muscle GABAA receptors augments airway smooth muscle relaxation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L248–L256. [Google Scholar] [CrossRef] [Green Version]
- Comhair, S.A.A.; McDunn, J.; Bennett, C.; Fettig, J.; Erzurum, S.C.; Kalhan, S.C. Metabolomic Endotype of Asthma. J. Immunol. (Baltim. Md. 1950) 2015, 195, 643–650. [Google Scholar] [CrossRef] [Green Version]
- Bridges, C.C.; Ola, M.S.; Prasad, P.D.; El-Sherbeny, A.; Ganapathy, V.; Smith, S.B. Regulation of taurine transporter expression by NO in cultured human retinal pigment epithelial cells. Am. J. Physiol. Cell Physiol. 2001, 281, C1825–C1836. [Google Scholar] [CrossRef] [Green Version]
- Zinellu, A.; Fois, A.G.; Sotgia, S.; Zinellu, E.; Bifulco, F.; Pintus, G.; Mangoni, A.A.; Carru, C.; Pirina, P. Plasma protein thiols: An early marker of oxidative stress in asthma and chronic obstructive pulmonary disease. Eur. J. Clin. Investig. 2016, 46, 181–188. [Google Scholar] [CrossRef]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metab. Off. J. Metab. Soc. 2010, 6, 78–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, F.; He, X.; Deng, H.; Chen, Q.; Lu, J.; Spraul, M.; Yu, Y. Discrimination of metabolic profiles of pancreatic cancer from chronic pancreatitis by high-resolution magic angle spinning 1H nuclear magnetic resonance and principal components analysis. Cancer Sci. 2007, 98, 1678–1682. [Google Scholar] [CrossRef] [PubMed]
- Ascha, M.; Wang, Z.; Ascha, M.S.; Dweik, R.; Zein, N.N.; Grove, D.; Brown, J.M.; Marshall, S.; Lopez, R.; Hanouneh, I.A. Metabolomics studies identify novel diagnostic and prognostic indicators in patients with alcoholic hepatitis. World J. Hepatol. 2016, 8, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Kinny-Köster, B.; Bartels, M.; Becker, S.; Scholz, M.; Thiery, J.; Ceglarek, U.; Kaiser, T. Plasma Amino Acid Concentrations Predict Mortality in Patients with End-Stage Liver Disease. PLoS ONE 2016, 11, e0159205. [Google Scholar] [CrossRef]
- Budhathoki, S.; Iwasaki, M.; Yamaji, T.; Yamamoto, H.; Kato, Y.; Tsugane, S. Association of plasma concentrations of branched-chain amino acids with risk of colorectal adenoma in a large Japanese population. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2017, 28, 818–823. [Google Scholar] [CrossRef]
- Yu, D.; Moore, S.C.; Matthews, C.E.; Xiang, Y.-B.; Zhang, X.; Gao, Y.-T.; Zheng, W.; Shu, X.-O. Plasma metabolomic profiles in association with type 2 diabetes risk and prevalence in Chinese adults. Metab. Off. J. Metab. Soc. 2016, 12, 3. [Google Scholar] [CrossRef] [Green Version]
- Motta, A.; Paris, D.; D’Amato, M.; Melck, D.; Calabrese, C.; Vitale, C.; Stanziola, A.A.; Corso, G.; Sofia, M.; Maniscalco, M. NMR metabolomic analysis of exhaled breath condensate of asthmatic patients at two different temperatures. J. Proteome Res. 2014, 13, 6107–6120. [Google Scholar] [CrossRef]
- Ghosh, N.; Choudhury, P.; Subramani, E.; Saha, D.; Sengupta, S.; Joshi, M.; Banerjee, R.; Roychowdhury, S.; Bhattacharyya, P.; Chaudhury, K. Metabolomic signatures of asthma-COPD overlap (ACO) are different from asthma and COPD. Metab. Off. J. Metab. Soc. 2019, 15, 87. [Google Scholar] [CrossRef]
- Chiu, C.-Y.; Cheng, M.-L.; Chiang, M.-H.; Wang, C.-J.; Tsai, M.-H.; Lin, G. Metabolomic Analysis Reveals Distinct Profiles in the Plasma and Urine Associated with IgE Reactions in Childhood Asthma. J. Clin. Med. 2020, 9, 887. [Google Scholar] [CrossRef] [Green Version]
- Smith, I.C.P.; Blandford, D.E. Nuclear magnetic resonance spectroscopy. Anal. Chem. 1995, 67, 509–518. [Google Scholar] [CrossRef]
- Xiang, Y.-Y.; Wang, S.; Liu, M.; Hirota, J.A.; Li, J.; Ju, W.; Fan, Y.; Kelly, M.M.; Ye, B.; Orser, B.; et al. A GABAergic system in airway epithelium is essential for mucus overproduction in asthma. Nat. Med. 2007, 13, 862–867. [Google Scholar] [CrossRef] [PubMed]
- Tanianskii, D.A.; Jarzebska, N.; Birkenfeld, A.L.; O’Sullivan, J.F.; Rodionov, R.N. Beta-Aminoisobutyric Acid As a Novel Regulator of Carbohydrate and Lipid Metabolism. Nutrients 2019, 11, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifonova, O.P.; Lokhov, P.G.; Archakov, A.I. Metabolic profiling of human blood. Biomeditsinskaia Khimiia 2014, 60, 281–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Asthma Patients | Control Subjects | |
|---|---|---|---|
| No. of Subjects | 13 | 17 | |
| Sex | |||
| Male | 7 (53.8%) | 11 (64.7%) | |
| Female | 6 (46.2%) | 6 (35.3%) | |
| Age | |||
| Median | 12 | 10 | |
| Range | 4–16 | 3–18 | |
| Asthma Severity | |||
| Mild | 7 (53.8%) | ||
| Moderate | 4 (30.8%) | ||
| Severe | 2 (15.4%) | ||
| The Daily dose of Corticosteroids (Budesonide or Equivalent) | |||
| 100–200 μg | 6 (46.2%) | ||
| 250–350 μg | 2 (15.4%) | ||
| 400–500 μg | 3 (23.0%) | ||
| >500 μg | 1 (7.7%) | ||
| Unknown | 1 (7.7%) | ||
| Comorbidities | |||
| Hypoacusia | 1 | ||
| Atopic Dermatitis | 1 | ||
| Allergic Rhinitis | 3 | ||
| Coeliac Disease | 1 | ||
| Cholecystitis | 1 | ||
| Lung Function | |||
| FEV1/VC | Mean | 85.25% | |
| Range | 67–94% | ||
| FEV1 | Mean | 82.5% | |
| Range | 40–97% | ||
| FVC EX | Mean | 87.33% | |
| Range | 49–95% | ||
| Total IgE | |||
| Mean | 278.61 kU/I | 96.29 kU/I | |
| Range | 15.5–1035 kU/I | 12.2–797 kU/I | |
| Amino Acid | Abbreviation | Children with Asthma (n = 13) | Control Group (n = 17) | ||||
|---|---|---|---|---|---|---|---|
| Median | Mean | SD | Median | Mean | SD | ||
| 1-Methyl-L-Histidine | 1MHis | 3.09 | 4.42 | 4.4 | 1.74 | 1.93 | 1.73 |
| 3-Methyl-L-Histidine | 3MHis | 2.38 | 2.52 | 0.84 | 2.04 | 2.32 | 1.06 |
| L-A-Aminoadipic Acid | Aad | 0.64 | 0.66 | 0.2 | 0.66 | 0.78 | 0.29 |
| L-A-Amino-N-Butyric Acid | Abu | 15.56 | 16.72 | 5.24 | 18.98 | 19.53 | 6.16 |
| L-Alanine | Ala | 443.71 | 423.32 | 89.17 | 368.45 | 396.24 | 106.97 |
| L-Arginine | Arg | 112.59 | 109.97 | 12.66 | 87.67 | 93.15 | 22.77 |
| L-Asparagine | Asn | 46.87 | 49.32 | 5.23 | 51.71 | 52.24 | 11.23 |
| L-Aspartic Acid | Asp | 11.71 | 11.94 | 3.65 | 9.89 | 11.65 | 7.89 |
| D,L-Β-Aminoisobutyric Acid | bAib | 1.13 | 1.11 | 0.31 | 1.26 | 1.47 | 0.61 |
| Β-Alanine | bAla | 16.51 | 13.84 | 6.61 | 8.95 | 9.64 | 3.9 |
| L-Citrulline | Cit | 24.37 | 23.77 | 5.46 | 24.71 | 24.31 | 3.79 |
| Ethanolamine | EtN | 8.94 | 8.96 | 2.17 | 8.35 | 8.96 | 1.65 |
| Ƴ-Amino-N-Butyric Acid | GABA | 1.25 | 1.18 | 0.49 | 0.67 | 0.75 | 0.31 |
| L-Glutamine | Gln | 424.51 | 429.37 | 56.74 | 459.64 | 458.41 | 71.53 |
| L-Glutamic Acid | Glu | 52.97 | 58 | 19.77 | 64.58 | 62.66 | 18.27 |
| Glycine | Gly | 278.17 | 283.25 | 32.45 | 255.44 | 268.74 | 46.85 |
| L-Histidine | His | 69.11 | 68.38 | 7.93 | 61.62 | 66.83 | 13.1 |
| Hydroxy-L-Proline | Hyp | 22.02 | 22.44 | 14.17 | 13.32 | 15.94 | 6.71 |
| L-Isoleucine | Ile | 63.9 | 61.95 | 12.54 | 63.32 | 68.1 | 16.01 |
| L-Leucine | Leu | 92.42 | 97.89 | 18.21 | 104.96 | 111.25 | 25.25 |
| L-Lysine | Lys | 142.42 | 147.4 | 25.22 | 150.06 | 157.79 | 40.29 |
| L-Methionine | Met | 21.32 | 22.06 | 5.39 | 22.45 | 23.44 | 7.17 |
| L-Ornithine | Orn | 59.45 | 64.63 | 15.76 | 61.35 | 65.67 | 18.49 |
| L-Phenylalanine | Phe | 55.46 | 52.55 | 8.97 | 54.67 | 54.85 | 9.84 |
| L-Proline | Pro | 197.03 | 181.4 | 67.24 | 171.13 | 177.63 | 51.23 |
| Sarcosine | Sar | 1.28 | 1.24 | 0.74 | 1.26 | 1.26 | 0.51 |
| L-Serine | Ser | 133.87 | 140.51 | 14.25 | 140.57 | 139.22 | 22.81 |
| Taurine | Tau | 66.95 | 70.75 | 17.75 | 86.33 | 85.95 | 16.55 |
| L-Threonine | Thr | 94.75 | 101.29 | 27.81 | 97.24 | 96.91 | 20.15 |
| L-Tryptophan | Trp | 51.47 | 51.61 | 7.49 | 57.34 | 59.19 | 13.14 |
| L-Tyrosine | Tyr | 52.31 | 52.57 | 11.82 | 51.51 | 55.81 | 18.07 |
| L-Valine | Val | 165.8 | 173.07 | 29.58 | 195.68 | 204.36 | 38.34 |
| Amino Acid | p Value | |||||
|---|---|---|---|---|---|---|
| Shapiro–Wilk Test | Levene’s Test | Mann–Whitney U Test | Student’s t-Test | Welch’s t-Test | ||
| Children with Asthma (n = 13) | Control Group (n = 17) | |||||
| 1MHis | 0.052421 | 0.003404 | 0.276532 | |||
| 3MHis | 0.875277 | 0.234623 | 0.327524 | 0.591304 | ||
| Aad | 0.496739 | 0.112430 | 0.07644 | 0.216463 | ||
| Abu | 0.621549 | 0.972163 | 0.644789 | 0.198044 | ||
| Ala | 0.178613 | 0.371844 | 0.669371 | 0.467315 | ||
| Arg | 0.288521 | 0.071223 | 0.043268 | 0.016265 | ||
| Asn | 0.469455 | 0.584648 | 0.009499 | 0.353783 | ||
| Asp | 0.753162 | 0.000012 | 0.241259 | |||
| bAib | 0.694310 | 0.326845 | 0.020441 | 0.045451 | ||
| bAla | 0.385620 | 0.326401 | 0.034083 | 0.056388 | ||
| Cit | 0794372 | 0.951293 | 0.206713 | 0.753887 | ||
| EtN | 0.128147 | 0.172044 | 0.617651 | 0.997393 | ||
| GABA | 0.227097 | 0.609400 | 0.099223 | 0.006383 | ||
| Gln | 0.389567 | 0.267469 | 0.442514 | 0.239702 | ||
| Glu | 0.572162 | 0.911833 | 0.656593 | 0.509076 | ||
| Gly | 0.437805 | 0.194367 | 0.324685 | 0.348727 | ||
| His | 0.813939 | 0.357040 | 0.015545 | 0.690912 | ||
| Hyp | 0.093057 | 0.017077 | 0.167247 | |||
| Ile | 0.738028 | 0.211595 | 0.309006 | 0.263792 | ||
| Leu | 0.212462 | 0.279189 | 0.302909 | 0.118475 | ||
| Lys | 0.752616 | 0.270535 | 0.367141 | 0.422490 | ||
| Met | 0.792258 | 0.308677 | 0.289639 | 0.566910 | ||
| Orn | 0.026378 | 0.288794 | 0.769551 | |||
| Phe | 0.185401 | 0.541752 | 0.844958 | 0.515613 | ||
| Pro | 0.173926 | 0.379691 | 0.219423 | 0.862817 | ||
| Sar | 0.547304 | 0.953359 | 0.354722 | 0.953930 | ||
| Ser | 0.132290 | 0.220079 | 0.300592 | 0.859334 | ||
| Tau | 0.011918 | 0.966747 | 0.012036 | |||
| Thr | 0.615151 | 0.330271 | 0.439175 | 0.619944 | ||
| Trp | 0.301341 | 0.046520 | 0.131898 | |||
| Tyr | 0.562556 | 0.220185 | 0.094828 | 0.580349 | ||
| Val | 0.773498 | 0.170362 | 0.652018 | 0.021465 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matysiak, J.; Klupczynska, A.; Packi, K.; Mackowiak-Jakubowska, A.; Bręborowicz, A.; Pawlicka, O.; Olejniczak, K.; Kokot, Z.J.; Matysiak, J. Alterations in Serum-Free Amino Acid Profiles in Childhood Asthma. Int. J. Environ. Res. Public Health 2020, 17, 4758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17134758
Matysiak J, Klupczynska A, Packi K, Mackowiak-Jakubowska A, Bręborowicz A, Pawlicka O, Olejniczak K, Kokot ZJ, Matysiak J. Alterations in Serum-Free Amino Acid Profiles in Childhood Asthma. International Journal of Environmental Research and Public Health. 2020; 17(13):4758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17134758
Chicago/Turabian StyleMatysiak, Joanna, Agnieszka Klupczynska, Kacper Packi, Anna Mackowiak-Jakubowska, Anna Bręborowicz, Olga Pawlicka, Katarzyna Olejniczak, Zenon J. Kokot, and Jan Matysiak. 2020. "Alterations in Serum-Free Amino Acid Profiles in Childhood Asthma" International Journal of Environmental Research and Public Health 17, no. 13: 4758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17134758