Transcriptional Profiling and Biological Pathway(s) Analysis of Type 2 Diabetes Mellitus in a Pakistani Population †

 , ,
, , 
Abstract
:1. Introduction
2. Methods
2.1. Study Participants, Ethics, and Selection of Participants for Gene-Expression Assays
2.2. Blood Collection and RNA Preparation
2.3. cDNA Synthesis and Microarrays
2.4. Gene Expression Data Analysis
2.5. Identification of Cellular Processes and Pathways Involved by Ingenuity Pathways Analysis (IPA®)
2.6. Validation by High-Throughput TaqMan® Low Density Array (TLDA)
2.7. TLDA Data Analysis
3. Results
3.1. Differential Expression of Genes of T2DM Subjects
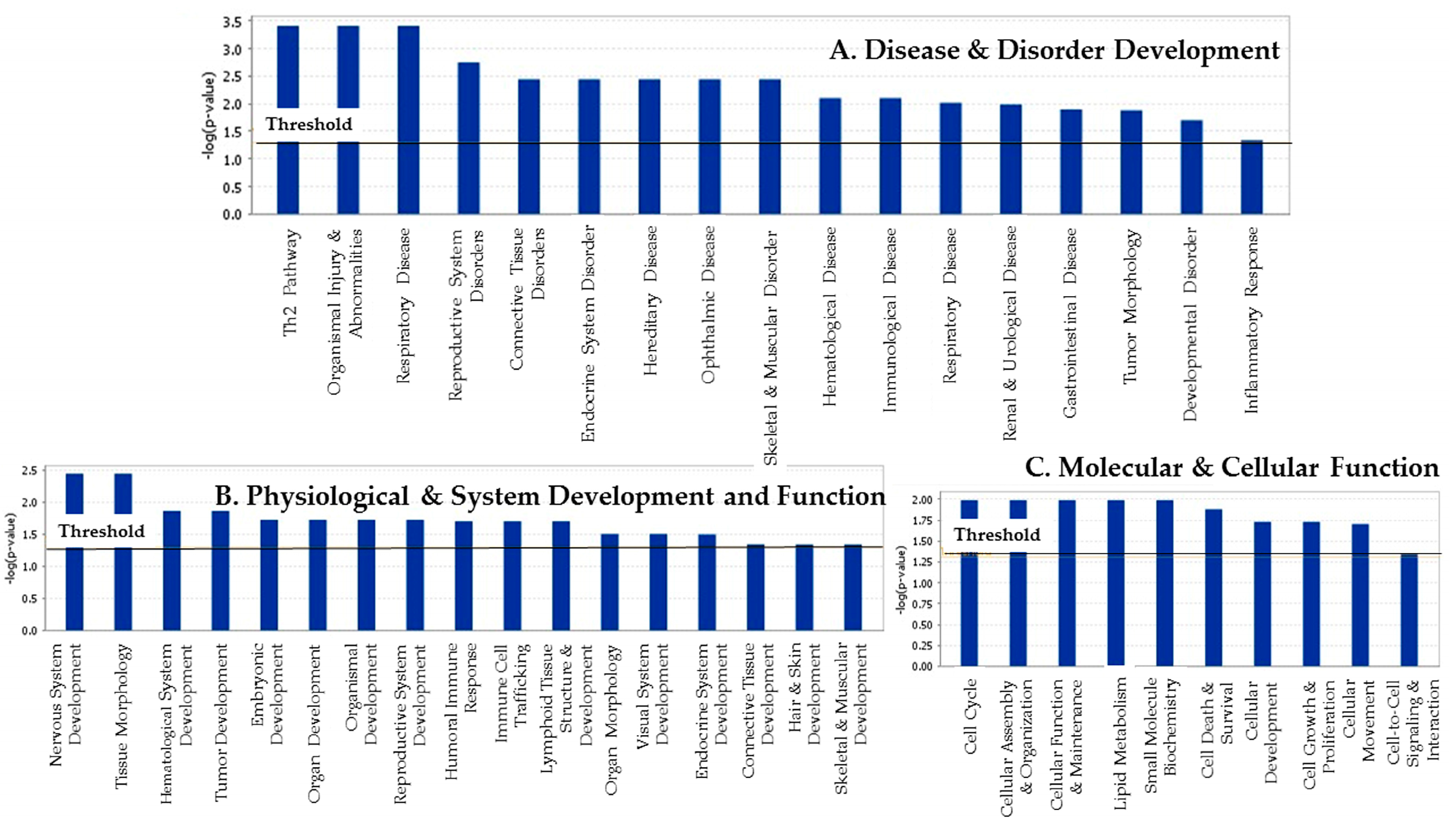
3.2. The Attribution of Differentially Expressed Genes to Their Biofunctions and Associated Diseases and Disorders
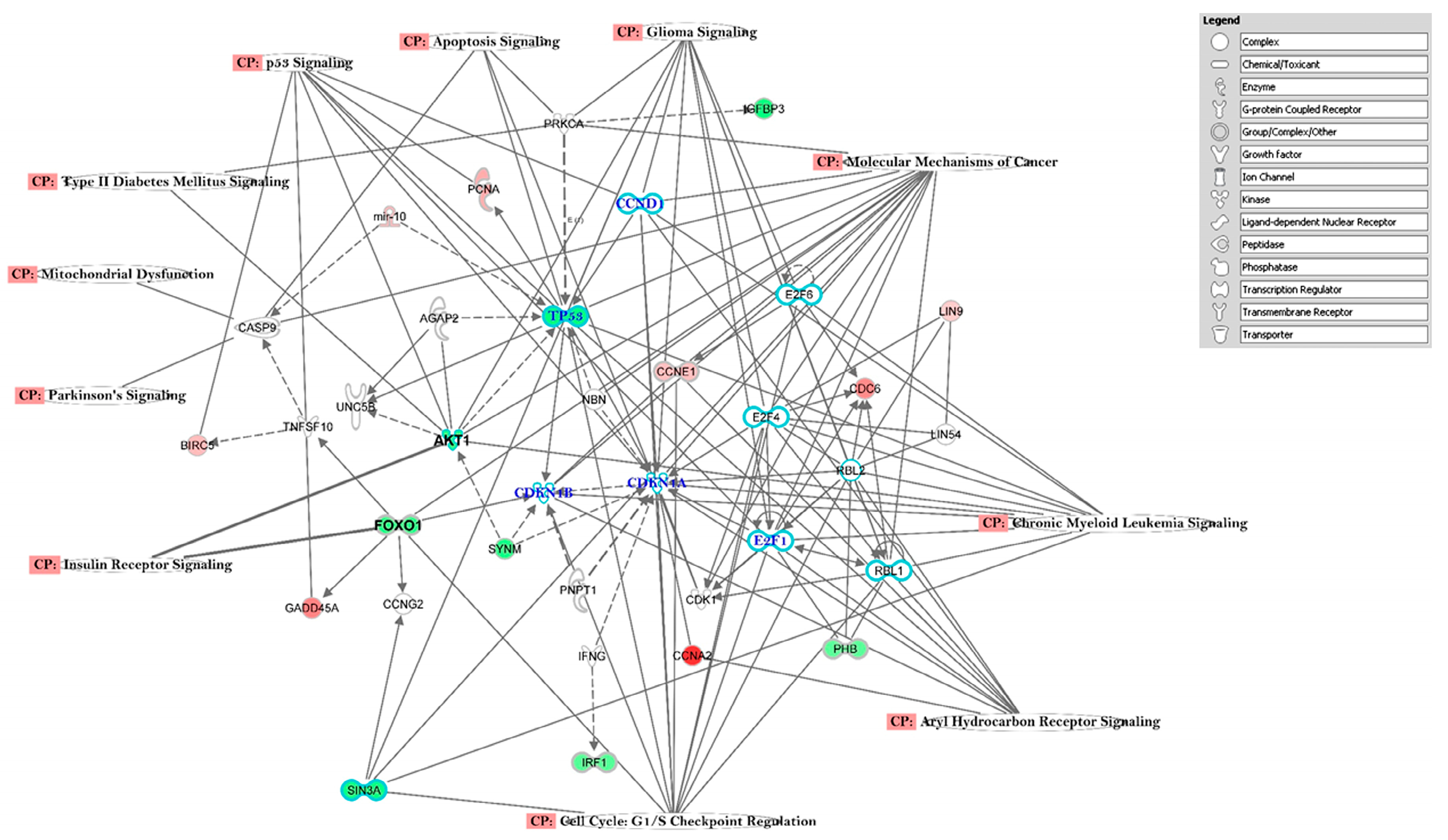
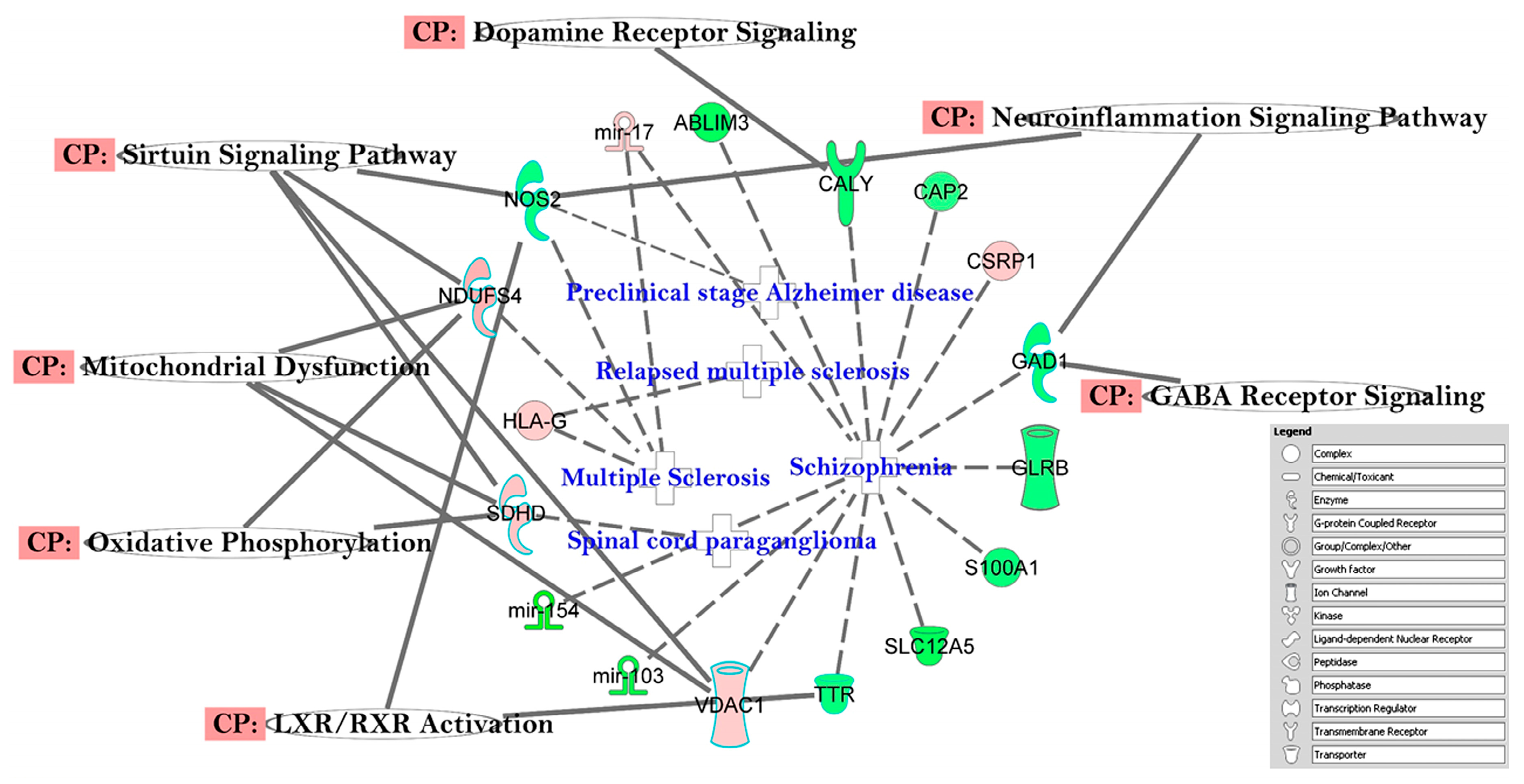
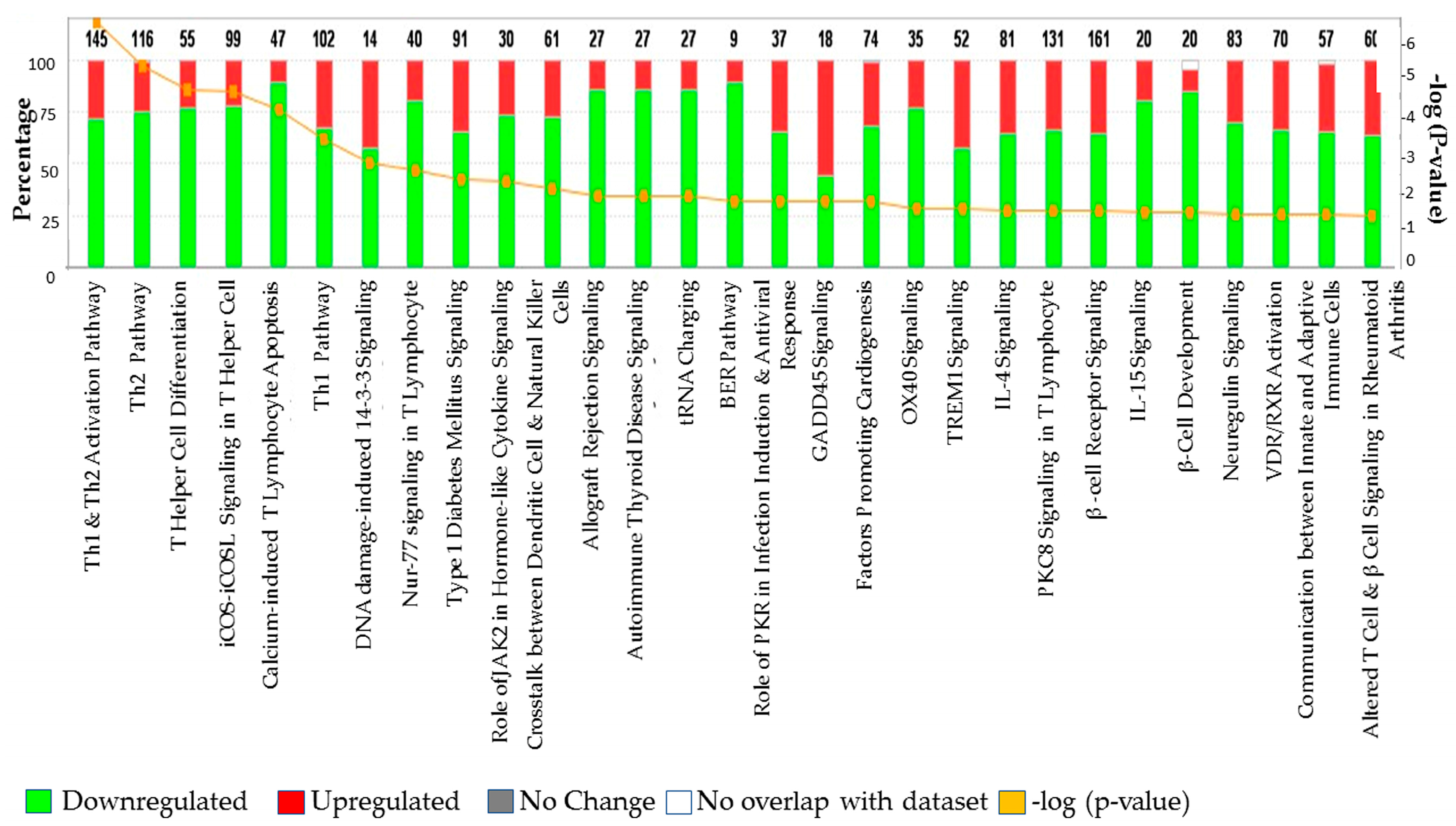
3.3. The Canonical Pathways (CP) and Gene Ontology (GO) Enrichment of Biological Processes
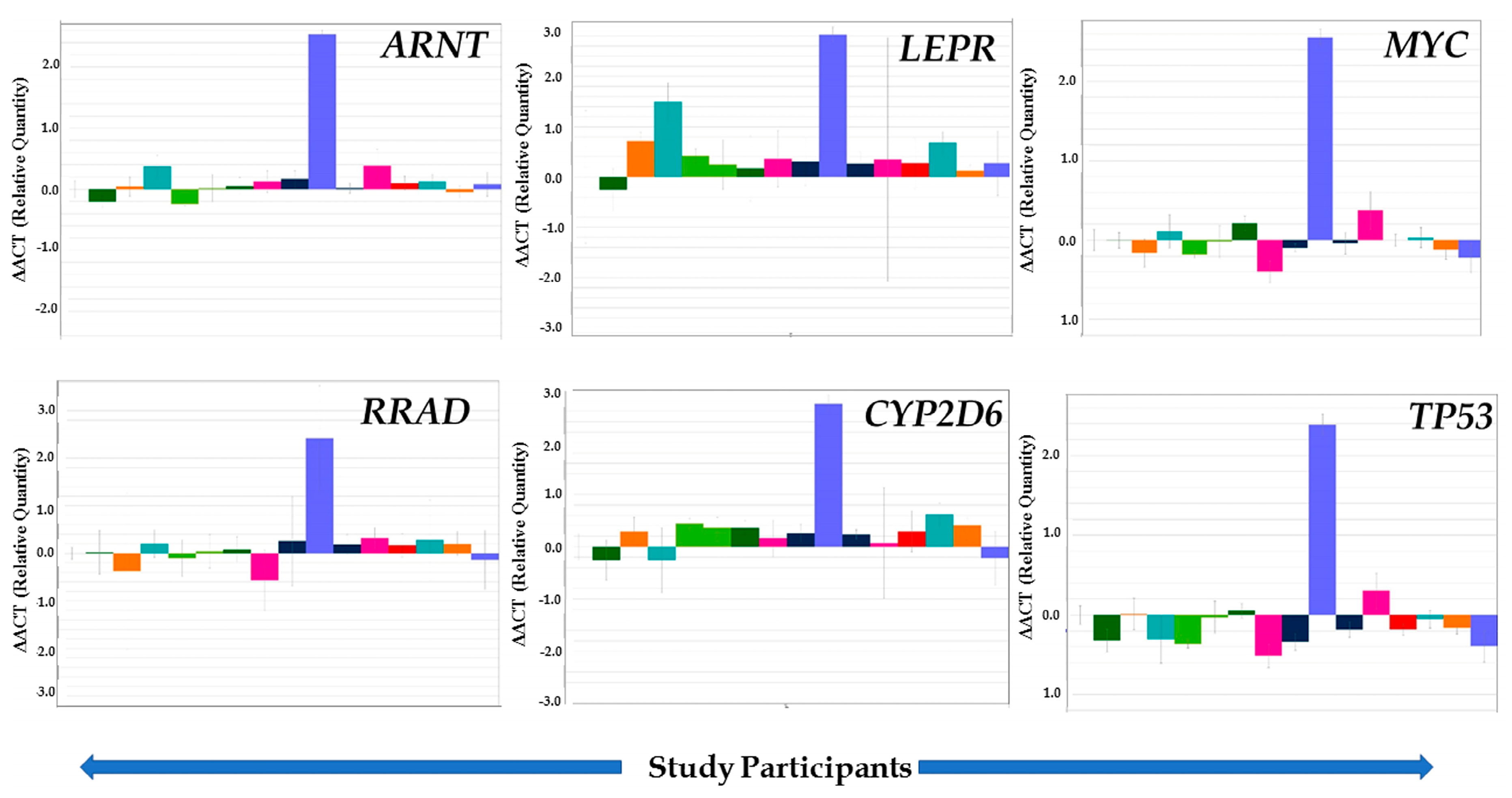
3.4. Validation of Selected Genes through TLDA
4. Discussion
4.1. Genes Associated with T2DM
4.2. Altered Pathways in T2DM
4.3. Strengths and Weaknesses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Noreen, Z.; DeJesus, J.; Bhatti, A.; Loffredo, C.A.; John, P.; Khan, J.S.; Ghosh, S. Epidemiological Investigation of Type 2 Diabetes and Alzheimer’s Disease in a Pakistani Population. Int. J. Environ. Res. Public Health 2018, 15, 1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar-Tana, J. Type 2 diabetes—Unmet need, unresolved pathogenesis, mTORC1-centric paradigm. Rev. Endocr. Metab. Disord. 2020, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.K.; Han, K.; Kim, M.K.; Koh, E.S.; Kim, E.S.; Nam, G.E.; Kwon, H.S. Changes in metabolic syndrome and its components and the risk of type 2 diabetes: A nationwide cohort study. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bonnefond, A.; Froguel, P. Rare and common genetic events in type 2 diabetes: What should biologists know? Cell Metab. 2015, 21, 357–368. [Google Scholar] [CrossRef] [Green Version]
- González, E.M.; Johansson, S.; Wallander, M.A.; Rodríguez, L.G. Trends in the prevalence and incidence of diabetes in the UK: 1996–2005. J. Epidemiol. Community Health 2009, 63, 332–336. [Google Scholar] [CrossRef]
- Cohrs, C.M.; Panzer, J.K.; Drotar, D.M.; Enos, S.J.; Kipke, N.; Chen, C.; Brennand, A. Dysfunction of persisting β cells is a key feature of early type 2 diabetes pathogenesis. Cell Rep. 2020, 31, 107469. [Google Scholar] [CrossRef]
- Abdul-Ghani, M.A.; Tripathy, D.; DeFronzo, R.A. Top of Form Contributions of beta-cell dysfunction and insulin resistance to the pathogenesis of impaired glucose tolerance and impaired fasting glucose. Diabetes Care. 2006, 29, 1130–1139. [Google Scholar] [CrossRef]
- Abedini, M.R.; Bijari, B.; Miri, Z.; Emampour, F.S.; Abbasi, A. The quality of life of the patients with diabetes type 2 using EQ-5D-5 L in Birjand. Health Qual. Life Outcomes 2020, 18, 18. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Torres, A.V.; Medina-Bravo, P.; Valerio-Pérez, B.E.; Mendoza-Salmeron, G.; Escobedo-dela-Peña, J.; Velázquez-López, L. Positive health beliefs are associated with improvement of glycated hemoglobin and lipid profiles in Mexican patients with type 2 diabetes mellitus: A cross-sectional study. BMC Public Health 2020, 20, 761. [Google Scholar] [CrossRef]
- Cho, N.H.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.D.; Ohlrogge, A.W.; Malanda, B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. 2018, 138, 271–281. [Google Scholar] [CrossRef]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88. [Google Scholar] [CrossRef] [PubMed]
- Uddin, F.; Ali, B.; Junaid, N. Prevalence of diabetic complications in newly diagnosed type 2 diabetes patients in Pakistan: Findings from national registry. J. Ayub Med. Coll. Abbottabad 2019, 30, S652–S658. [Google Scholar]
- Patel, C.J.; Chen, R.; Kodama, K.; Ioannidis, J.P.; Butte, A.J. Systematic identification of interaction effects between genome-and environment-wide associations in type 2 diabetes mellitus. Hum. Genet. 2013, 132, 495–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, R.B.; Groop, L. Genetics of type 2 diabetes—Pitfalls and possibilities. Genes 2015, 6, 87–123. [Google Scholar] [CrossRef] [Green Version]
- Sanghera, D.K.; Blackett, P.R. Type 2 diabetes genetics: Beyond GWAS. J. Diabetes Metab. 2012, 3, 6948. [Google Scholar] [CrossRef] [Green Version]
- Balk, E.M.; Earley, A.; Raman, G.; Avendano, E.A.; Pittas, A.G.; Remington, P.L. Combined Combined diet and physical activity promotion programs to prevent type 2 diabetes among persons at increased risk: A systematic review for the Community Preventive Services Task Force. Ann. Intern. Med. 2015, 163, 437–451. [Google Scholar] [CrossRef] [Green Version]
- Overvad, K.; Dahm, C.C. Association between dietary meat consumption and incident type 2 diabetes: The EPIC-InterAct study. Diabetologia 2013, 56, 47–59. [Google Scholar]
- Kohei, K. Pathophysiology of type 2 diabetes and its treatment policy. Jpn. Med. Assoc. J. 2010, 53, 41–46. [Google Scholar]
- Centers for Disease Control and Prevention. National Diabetes Statistics Report. 2020. Available online: https://www.cdc.gov/diabetes/pdfs/data/statistics/national-diabetes-statistics-report.pdf (accessed on 2 June 2020).
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Shaw, J.E. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas. Diabetes Res. Clin. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Ali, I. Diabetes mellitus in Pakistan: A major public health concern. Arch. Pharm. Pr. 2016, 7, 30–33. [Google Scholar]
- Jayawardena, R.; Ranasinghe, P.; Byrne, N.M.; Soares, M.J.; Katulanda, P.; Hills, A.P. Prevalence and trends of the diabetes epidemic in South Asia: A systematic review and meta-analysis. BMC Public Health 2012, 12, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.K.; Sharma, V. World Diabetes Day 2018: Battling the Emerging Epidemic of Diabetic Retinopathy. Indian J. Ophthalmol. 2018, 66, 1652–1653. [Google Scholar] [CrossRef] [PubMed]
- Mathers, C.D.; Loncar, D. Projections of global mortality and burden of disease from 2002 to 2030. PLoS Med. 2006, 3, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, P.; Kawar, B.; El-Nahas, M. Obesity and diabetes in the developing world—A growing challenge. N. Engl. J. Med. 2007, 356, 213–215. [Google Scholar] [CrossRef] [Green Version]
- Aamir, A.H.; Ul-Haq, Z.; Mahar, S.A.; Qureshi, F.M.; Ahmad, I.; Jawa, A.; Ishtiaq, O. Diabetes Prevalence Survey of Pakistan (DPS-PAK): Prevalence of type 2 diabetes mellitus and prediabetes using HbA1c: A population-based survey from Pakistan. BMJ. Open 2019, 9, e025300. [Google Scholar] [CrossRef]
- Forouhi, N.G.; Luan, J.; Hennings, S.; Wareham, N.J. Incidence of Type 2 diabetes in England and its association with baseline impaired fasting glucose: The Ely study 1990–2000. Diabet. Med. 2007, 24, 200–207. [Google Scholar] [CrossRef]
- Wild, S.; Roglic, G.; Green, A.; Sicree, R.; King, H. Global prevalence of diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care 2004, 27, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Basit, A.; Riaz, M.; Fawwad, A. Improving diabetes care in developing countries: The example of Pakistan. Diabetes Res. Clin. 2015, 107, 224–232. [Google Scholar] [CrossRef]
- Guariguata, L.; Whiting, D.; Hambleton, I.R.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. 2014, 103, 137–149. [Google Scholar] [CrossRef]
- Meo, S.A.; Zia, I.; Bukhari, I.A.; Arain, S.A. Type 2 diabetes mellitus in Pakistan: Current prevalence and future forecast. J. Pak. Med Assoc. 2016, 66, 1637–1642. [Google Scholar]
- Hu, F.B. Globalization of diabetes: The role of diet, lifestyle, and genes. Diabetes Care 2011, 34, 1249–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elobeid, M.A.; Padilla, M.A.; Brock, D.W.; Ruden, D.M.; Allison, D.B. Endocrine disruptors and obesity: An examination of selected persistent organic pollutants in the NHANES 1999–2002 data. Int. J. Environ. Res. Public Health 2010, 7, 2988–3005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Bertrand, K.A.; Choi, A.L.; Hu, F.B.; Laden, F.; Grandjean, P.; Sun, Q. Persistent organic pollutants and type 2 diabetes: A prospective analysis in the nurses’ health study and meta-analysis. Environ. Health Perspect. 2013, 121, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, G.; Valvi, D.; Coull, B.; Göen, T.; Hu, F.B.; Nielsen, F.; Sun, Q. Persistent organic pollutants and risk of type 2 diabetes: A prospective investigation among middle-aged women in Nurses’ Health Study II. Environ. Int. 2018, 114, 334–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Murinova, L.; Trnovec, T.; Loffredo, C.A.; Washington, K.; Mitra, P.S.; Dutta, S.K. Biomarkers linking PCB exposure and obesity. Curr. Pharm. Biotechnol. 2014, 15, 1058–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Mitra, P.S.; Loffredo, C.A.; Trnovec, T.; Murinova, L.; Sovcikova, E.; Dutta, S.K. Transcriptional profiling and biological pathway analysis of human equivalence PCB exposure in vitro: Indicator of disease and disorder development in humans. Environ. Res. 2015, 138, 202–216. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Trnovec, T.; Palkovicova, L.; Hoffman, E.P.; Washington, K.; Dutta, S.K. Status of LEPR gene in PCB-exposed population: A quick look. Int, J. Hum. Genet. 2013, 13, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Hoffman, E.P. Embryonic myogenesis pathways in muscle regeneration. Dev. Dyn. 2004, 229, 380–392. [Google Scholar] [CrossRef]
- Ghosh, S.; Loffredo, C.A.; Mitra, P.S.; Trnovec, T.; Murinova, L.P.; Sovcikova, E.; Dutta, S.K. PCB exposure and potential future cancer incidence in Slovak children: An assessment from molecular finger printing by Ingenuity Pathway Analysis (IPA®) derived from experimental and epidemiological investigations. Environ. Sci. Pollut. Res. 2018, 25, 16493–16507. [Google Scholar] [CrossRef]
- Wu, Z.; Irizarry, R.A. Preprocessing of oligonucleotide array data. Nat. Biotechnol. 2004, 22, 656–658. [Google Scholar] [CrossRef]
- Kaizer, E.C.; Glaser, C.L.; Chaussabel, D.; Banchereau, J.; Pascual, V.; White, P.C. Gene expression in peripheral blood mononuclear cells from children with diabetes. J. Clin. Endocrinol. Metab. 2007, 92, 3705–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, P.S.; Ghosh, S.; Zang, S.; Sonneborn, D.; Hertz-Picciotto, I.; Trnovec, T.; Dutta, S.K. Analysis of the toxicogenomic effects of exposure to persistent organic pollutants (POPs) in Slovakian girls: Correlations between gene expression and disease risk. Environ. Int. 2012, 39, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Aschner, P.; Gagliardino, J.J.; Ilkova, H.; Lavalle, F.; Ramachandran, A.; Mbanya, J.C.; Chan, J.C. Persistent poor glycaemic control in individuals with type 2 diabetes in developing countries: 12 years of real-world evidence of the International Diabetes Management Practices Study (IDMPS). Diabetologia 2020, 63, 711–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmanfarma, K.K.; Ansari-Moghaddam, A.; Zareban, I.; Adineh, H.A. Prevalence of type 2 diabetes in Middle–East: Systematic review& meta-analysis. Prim. Care Diabetes 2020, 14, 297–304. [Google Scholar]
- Das, S.K.; Sharma, N.K.; Chu, W.S.; Wang, H.; Elbein, S.C. Aryl hydrocarbon receptor nuclear translocator (ARNT) gene as a positional and functional candidate for type 2 diabetes and prediabetic intermediate traits: Mutation detection, case-control studies, and gene expression analysis. BMC Med. Genet. 2008, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Regulation of mammalian O2 homeostasis by hypoxia-inducible factor. Annu. Rev. Cell Dev. Biol. 1999, 15, 551–578. [Google Scholar] [CrossRef]
- Lau, S.M.; Cha, K.M.; Karunatillake, A.; Stokes, R.A.; Cheng, K.; McLean, M.; Gunton, J.E. Beta-cell ARNT is required for normal glucose tolerance in murine pregnancy. PLoS ONE 2013, 8, e77419. [Google Scholar] [CrossRef]
- Hu, Z.; Jiao, R.; Wang, P.; Zhu, Y.; Zhao, J.; De Jager, P.; Bennett, D.A.; Jin, L.; Xiong, M. Shared Causal Paths underlying Alzheimer’s dementia and Type 2 Diabetes. Sci. Rep. 2020, 10, 4107. [Google Scholar] [CrossRef] [Green Version]
- Abella, V.; Scotece, M.; Conde, J.; Pino, J.; Gonzalez-Gay, M.A.; Gomez-Reino, J.J.; Gualillo, O. Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat. Rev. Rheumatol. 2017, 13, 100. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, Y.; Heiman, M.; DiMarchi, R. Leptin: Structure, function and biology. Vitam. Horm. 2005, 71, 345–372. [Google Scholar]
- Denroche, H.C.; Huynh, F.K.; Kieffer, T.J. The role of leptin in glucose homeostasis. J. Diabetes Invest. 2012, 3, 115–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, W.L.; Chen, C.C.; Chang, C.T.; Wu, J.Y.; Chen, C.H.; Huang, Y.C.; Tsai, F.J. Gene polymorphisms of adiponectin and leptin receptor are associated with early onset of type 2 diabetes mellitus in the Taiwanese population. Int. J. Obes. 2012, 36, 790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadzadeh, G.; Nikzamir, A.; Mohammadi, J.; Pourdashti, S.; Shabazian, H.; Latifi, S.M. Association of the 223A/G LEPR polymorphism with serum leptin levels in Iranian subjects with type 2 diabetes. Arch. Iran. Med. 2013, 16, 636–641. [Google Scholar] [PubMed]
- Yang, M.M.; Wang, J.; Fan, J.J.; Ng, T.K.; Sun, D.J.; Guo, X.; Li, Y.B. Variations in the obesity gene “LEPR” contribute to risk of type 2 diabetes mellitus: Evidence from a meta-analysis. J. Diabetes Res. 2016, 2016, 5412084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherian, P.T.; Al-Khairi, I.; Sriraman, D.; Al-Enezi, A.; Al-Sultan, D.; Al-Otaibi, M.; Abu-Farha, M. Increased Circulation and Adipose Tissue Levels of DNAJC27/RBJ in Obesity and Type 2-Diabetes. Front. Endocrinol. 2018, 9, 423. [Google Scholar] [CrossRef]
- Hur, J.; Sullivan, K.A.; Pande, M.; Hong, Y.; Sima, A.A.; Jagadish, H.V.; Feldman, E.L. The identification of gene expression profiles associated with progression of human diabetic neuropathy. Brain 2011, 134, 3222–3235. [Google Scholar] [CrossRef]
- Liu, X.; Li, X.; Li, C.; Gong, C.; Liu, S.; Shi, Y. Study on regulation of adipokines on body fat distribution and its correlation with metabolic syndrome in type 2 diabetes mellitus. Minerva Endocrinol. 2019, 44, 259–263. [Google Scholar] [CrossRef]
- Qadir, M.I.; Ahmed, Z. lep Expression and its role in obesity and type-2 diabetes. Crit. Rev. Eukaryot. Gene 2017, 27, 47–51. [Google Scholar] [CrossRef]
- Dagdan, B.; Chuluun-Erdene, A.; Sengeragchaa, O.; Malchinkhuu, M.; Janlav, M. Leptin gene G2548A polymorphism among mongolians with metabolic syndrome. Med Sci. 2019, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Y.; Wang, H.; Yang, X.X.; Wu, J.J.; Geng, H.Y.; Kim, H.J.; Wang, L.S. LEPR gene Gln223Arg polymorphism and type 2 diabetes mellitus: A meta-analysis of 3,367 subjects. Oncotarget 2017, 8, 61927. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Zhang, C.; Zhang, F.; Li, H.; Yang, X.; Tang, X. The association between leptin receptor gene polymorphisms and type 2 diabetes mellitus: A systematic review and meta-analysis. Diabetes Res. Clin. 2016, 121, 49–58. [Google Scholar] [CrossRef]
- Zhang, L.; Qin, Y.; Liang, D.; Li, L.; Liang, Y.; Chen, L.; Zhang, H. Association of polymorphisms in LEPR with type 2 diabetes and related metabolic traits in a Chinese population. Lipids Health Dis. 2018, 17, 2. [Google Scholar] [CrossRef] [Green Version]
- Laville, M.; Auboeuf, D.; Khalfallah, Y.; Vega, N.; Riou, J.P.; Vidal, H. Acute regulation by insulin of phosphatidylinositol-3-kinase, Rad, Glut 4, and lipoprotein lipase mRNA levels in human muscle. J. Clin. Invest. 1996, 98, 43–49. [Google Scholar] [CrossRef]
- Moyers, J.S.; Bilan, P.J.; Reynet, C.; Kahn, C.R. Overexpression of Rad inhibits glucose uptake in cultured muscle and fat cells. J. Biol. 1996, 271, 23111–23116. [Google Scholar] [CrossRef] [Green Version]
- Sreekumar, R.; Halvatsiotis, P.; Schimke, J.C.; Nair, K.S. Gene expression profile in skeletal muscle of type 2 diabetes and the effect of insulin treatment. Diabetes 2002, 51, 1913–1920. [Google Scholar] [CrossRef] [Green Version]
- Ilany, J.; Bilan, P.J.; Kapur, S.; Caldwell, J.S.; Patti, M.E.; Marette, A.; Kahn, C.R. Overexpression of Rad in muscle worsens diet-induced insulin resistance and glucose intolerance and lowers plasma triglyceride level. Proc. Natl. Acad. Sci. USA 2006, 103, 4481–4486. [Google Scholar] [CrossRef] [Green Version]
- Reynet, C.; Kahn, C.R. Rad: A member of the Ras family overexpressed in muscle of type II diabetic humans. Science 1993, 262, 1441–1444. [Google Scholar] [CrossRef]
- Nelson, D.R.; Zeldin, D.C.; Hoffman, S.M.; Maltais, L.J.; Wain, H.M.; Nebert, D.W. Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes and alternative-splice variants. Pharmacogenetics 2004, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Bernard, S.; Neville, K.A.; Nguyen, A.T.; Flockhart, D.A. Interethnic differences in genetic polymorphisms of CYP2D6 in the US population: Clinical implications. Clin. Oncol. 2006, 11, 126–135. [Google Scholar]
- Nebert, D.W.; Dalton, T.P.; Okey, A.B.; Gonzalez, F.J. Role of aryl hydrocarbon receptor-mediated induction of the CYP1 enzymes in environmental toxicity and cancer. J. Biol. 2004, 279, 23847–23850. [Google Scholar] [CrossRef] [Green Version]
- Sissung, T.M.; Price, D.K.; Sparreboom, A.; Figg, W.D. Pharmacogenetics and regulation of human cytochrome P450 1B1: Implications in hormone-mediated tumor metabolism and a novel target for therapeutic intervention. Mol. Cancer Res. 2006, 4, 135–150. [Google Scholar] [CrossRef] [Green Version]
- Elfaki, I.; Almutairi, F.M.; Mir, R.; Khan, R.; Abu-Duhier, F. Cytochrome P450 Cyp1b1*2 Gene And Its Association with T2d In Tabuk Population, Northwestern Region Of Saudi Arabia. Asian J. Pharm. Clin. Res. 2018, 11, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Cohn, J.S.; Tremblay, M.; Boulet, L.; Jacques, H.; Davignon, J.; Roy, M.; Bernier, L. Plasma concentration and lipoprotein distribution of ApoC-I is dependent on ApoE genotype rather than the Hpa I ApoC-I promoter polymorphism. Atherosclerosis 2003, 169, 63–70. [Google Scholar] [CrossRef]
- Lauer, S.J.; Walker, D.; Elshourbagy, N.A.; Reardon, C.A.; Levy-Wilson, B.; Taylor, J.M. Two copies of the human apolipoprotein CI gene are linked closely to the apolipoprotein E gene. J. Biol. 1988, 263, 7277–7286. [Google Scholar]
- Hubacek, J.A.; Pitha, J.; Adamkova, V.; Skodova, Z.; Lanska, V.; Poledne, R. Apolipoprotein E and apolipoprotein CI polymorphisms in the Czech population: Almost complete linkage disequilibrium of the less frequent alleles of both polymorphisms. Physiol. Res. 2003, 52, 195–200. [Google Scholar]
- Xu, Y.; Berglund, L.; Ramakrishnan, R.; Mayeux, R.; Ngai, C.; Holleran, S.; Shachter, N.S. A common Hpa I RFLP of apolipoprotein CI increases gene transcription and exhibits an ethnically distinct pattern of linkage disequilibrium with the alleles of apolipoprotein E. J. Lipid Res. 1999, 40, 50–58. [Google Scholar]
- Hao, K.; Di Narzo, A.F.; Ho, L.; Luo, W.; Li, S.; Chen, R.; Pasinetti, G.M. Shared genetic etiology underlying Alzheimer’s disease and type 2 diabetes. Mol. Asp. Med. 2015, 43, 66–76. [Google Scholar] [CrossRef]
- Madhusudhanan, J.; Suresh, G.; Devanathan, V. Neurodegeneration in type 2 diabetes: Alzheimer’s as a case study. Brain Behav. 2020, 10, e01577. [Google Scholar] [CrossRef] [Green Version]
- Rad, S.K.; Arya, A.; Karimian, H.; Madhavan, P.; Rizwan, F.; Koshy, S.; Prabhu, G. Mechanism involved in insulin resistance via accumulation of β-amyloid and neurofibrillary tangles: Link between type 2 diabetes and Alzheimer’s disease. Drug Des. Dev. Ther. 2018, 12, 3999. [Google Scholar]
- Nasi, S.; Ciarapica, R.; Jucker, R.; Rosati, J.; Soucek, L. Making decisions through Myc. FEBS Lett. 2001, 490, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Kaneto, H.; Sharma, A.; Suzuma, K.; Laybutt, D.R.; Xu, G.; Bonner-Weir, S.; Weir, G.C. Induction of c-Myc expression suppresses insulin gene transcription by inhibiting NeuroD/BETA2-mediated transcriptional activation. J. Biol. 2002, 277, 12998–13006. [Google Scholar] [CrossRef] [Green Version]
- Pelengaris, S.; Khan, M.; Evan, G.I. Suppression of Myc-induced apoptosis in β cells exposes multiple oncogenic properties of Myc and triggers carcinogenic progression. Cell 2002, 109, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Laybutt, D.R.; Weir, G.C.; Kaneto, H.; Lebet, J.; Palmiter, R.D.; Sharma, A.; Bonner-Weir, S. Overexpression of c-Myc in β-cells of transgenic mice causes proliferation and apoptosis, downregulation of insulin gene expression, and diabetes. Diabetes 2002, 51, 1793–1804. [Google Scholar] [CrossRef] [Green Version]
- Riu, E.; Ferre, T.; Hidalgo, A.; Mas, A.; Franckhauser, S.; Otaegui, P.; Bosch, F. Overexpression Overexpression of c-myc in the liver prevents obesity and insulin resistance. FASEB J. 2003, 17, 1715–1717. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, Y.; Zhou, K.; Hong, A. Constructing regulatory networks to identify biomarkers for insulin resistance. Genes 2014, 539, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Lai, B.; Lai, Y.; Zhang, Y.; Zhou, M.; Sheng, L.; OuYang, G. The Solute Carrier Family 2 Genes Are Potential Prognostic Biomarkers in Acute Myeloid Leukemia. Technol. Cancer Res. Treat. 2020, 19, 1533033819894308. [Google Scholar] [CrossRef]
- Hong, C.T.; Chen, K.Y.; Wang, W.; Chiu, J.Y.; Wu, D.; Chao, T.Y.; Bamodu, O.A. Insulin Resistance Promotes Parkinson’s Disease through Aberrant Expression of α-Synuclein, Mitochondrial Dysfunction, and Deregulation of the Polo-Like Kinase 2 Signaling. Cells 2020, 9, 740. [Google Scholar] [CrossRef] [Green Version]
- De Felice, F.G.; Ferreira, S.T. Inflammation, defective insulin signaling, and mitochondrial dysfunction as common molecular denominators connecting type 2 diabetes to Alzheimer disease. Diabetes 2014, 63, 2262–2272. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Shimizu, I.; Minamino, T. The pathological role of adipose tissue aging in the progression of systemic insulin resistance. Inflamm. Regen. 2015, 35, 178–184. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Tu, B.; Wang, H.; Cao, Z.; Tang, M.; Zhang, C.; Zhao, Y. Tumor suppressor p53 cooperates with SIRT6 to regulate gluconeogenesis by promoting FoxO1 nuclear exclusion. Proc. Natl. Acad. Sci. USA 2014. [Google Scholar] [CrossRef] [Green Version]
- Kuricova, K.; Kuricova, L.; Dvorakova, V.; Kankova, K. Association of the Arg72Pro polymorphism in p53 with progression of diabetic nephropathy in T2DM subjects. J. Nephrol. Ther. 2014, 4. [Google Scholar] [CrossRef] [Green Version]
- Hager, K.M.; Gu, W. Understanding the non-canonical pathways involved in p53-mediated tumor suppression. Carcinogenesis 2013, 35, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Krook, A.; Roth, R.A.; Jiang, X.J.; Zierath, J.R.; Wallberg-Henriksson, H. Insulin-stimulated Akt kinase activity is reduced in skeletal muscle from NIDDM subjects. Diabetes 1998, 47, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Ambika, S.; Donna, S.; Edward, G.; Frank, B.H. Type 2 diabetes and risk of cancer. BMJ 2015, 350, g7707. [Google Scholar]
- Anastasi, E.; Filardi, T.; Tartaglione, S.; Lenzi, A.; Angeloni, A.; Morano, S. Linking type 2 diabetes and gynecological cancer: An introductory overview. Clin. Chem. Lab. Med. 2018, 56, 1413–1425. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, F.; Saito, E.; Lin, Y.; Song, M.; Luu, H.; Gupta, P.; Sawada, N.; Tamakoshi, A.; Shu, X.; et al. Association between type 2 diabetes and risk of cancer mortality: A pooled analysis of over 771,000 individuals in the Asia Cohort Consortium. Diabetologia 2017, 60, 1022–1032. [Google Scholar] [CrossRef]
- Pan, X.; He, M.; Yu, C.; Lv, J.; Guo, Y.; Bian, Z.; Yang, L.; Chen, Y.; Wu, T.; Chen, Z.; et al. Type 2 Diabetes and Risk of Incident Cancer in China: A Prospective Study Among 0.5 Million Chinese Adults. Am. J. Epidemiol. 2018, 187, 1380–1391. [Google Scholar] [CrossRef]
- Suh, S.; Kim, K. Diabetes and Cancer: Cancer Should Be Screened in Routine Diabetes Assessment. Diabetes Metab. J. 2019, 43, 733–743. [Google Scholar] [CrossRef]
- Vincent, E.E.; Yaghootkar, H. Using genetics to decipher the link between type 2 diabetes and cancer: Shared aetiology or downstream consequence? Diabetologia 2020. [Google Scholar] [CrossRef]
- Misra, A. Ethnic-Specific Criteria for Classification of Body Mass Index: A Perspective for Asian Indians and American Diabetes Association Position Statement. Diabetes Technol. Ther. 2015, 17, 667–671. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Top Functions and Disease | Significance (p-Value) |
|---|---|---|
| Top Canonical Pathways | ||
| B-Cell Development/Receptor Signaling | Immunological Disease/Cell Morphology/Immune Response | 8.17 × 10−6 |
| Altered B Cell Signaling | Hematological System development and Functions | 1.70 × 10−4 |
| Allograft Rejection Signaling | Cell to Cell Communication/Cellular Growth and Proliferation | 1.08 × 10−3 |
| Autoimmune Thyroid Disease Signaling | 1.08 × 10−3 | |
| Cyclins and Cell Cycle Regulations | Post-Translational Modification/Cell Cycle/ Connective Tissue Development | 1.77 × 102 |
| Role of NFAT in Regulation of Immune | Cellular Development, Growth and Proliferation | 1.86 × 10−2 |
| Response | ||
| PI3K/AKT Signaling | Cardiovascular Disease/Cardiovascular System Development and Function | 1.99 × 10−2 |
| NK-kB Activation | Cellular Development, Growth and Proliferation/ Hematological System Development and Function | 2.47 × 10−2 |
| PPAR Signaling | Gene Expression/Cardiovascular System Development and Function | 3.24 × 10−2 |
| Important Molecules | ||
| Disease and Disorders | ||
| Cancer | PPAR2, TP53, UBC, AKT1, CCL2, LRP6 | 3.87 × 10−4 |
| Organismal Injury and Abnormality | CCL2, CD48, PARP1, CCND2, TFP1, TP53, YUGB1 | 3.87 × 10−4 |
| Respiratory Disease | CCL2, CUBN, AKT1, KIF20A, CCR6, IGF20A | 3.87 × 10−4 |
| Reproductive System Disease | ATP78,BIRC5, CCL2, CITED2, PSMA1, RELA, TP53 | 1.97 × 10−3 |
| Connective Tissue Disorder | TERT, TP53 | 3.59 × 103 |
| Endocrine System Disorder | APCS, CALR, EZH2, PRDX6, TERT, TP53 | 3.59 × 10−3 |
| Ophthalmic Disease | TERT, TP53 | 3.59 × 10−3 |
| Immunological Disease | CD79A, CD79B,EZH2, LRP6, TNFRSF14, TP53 | 7.90 × 10−3 |
| Tumor Morphology | BIRC5, CD40, CD40LG, TP53 | 1.31 × 10−2 |
| Molecular and Cellular Functions | ||
| Cell Cycle | AURKA, TERT | 1.03 × 10−2 |
| Cellular Assembly and Organization | COL17A1, DST | 1.03 × 10−2 |
| Lipid Metabolism | ALG2, CD40LG | 1.03 × 10−2 |
| Small Molecule Biochemistry | ALG2, CD40LG | 1.03 × 10−2 |
| Cell Death and Survival | BIRC5, TP53, APOD, AKT1, SMARCA4, COL17A1 | 1.31 × 10−2 |
| Physiological System Development and Function | ||
| Nervous System Development and Function | CNTF,mir-196, OPN1MW | 3.59 × 10−3 |
| Tissue Morphology | CNTF, TP53 | 3.59 × 10−3 |
| Hematological System Development and Function | CCL2, FTL3LG, CD4, CD40 | 1.31 × 10−2 |
| Tissue Development | CCL2, DCM1, BMP15, ACVR1, ITGA5 | 1.37 × 10−2 |
| Endocrine System Development and Function | CCNE1, TCIM | 3.18 × 10−2 |
| Network ID | Genes in Network | Score | Focus Molecules | Functions |
|---|---|---|---|---|
| 1 | ADH1A, ADH4, APCS, CCNA2, CCNE1, CD40LG CDCA2, CDK2, CDK6, CDKN18, CENPE, CFHCKS2, CRP, FOXMI, HNF1A, IL6, KIF20A, MIF4GD PCK1, PCNA, TACR3, TGFB1, TMED10, AKT1 | 14 | 16 | Cell Cycle, DNA Replication, Recombination and Repair, Organismal Survival |
| 2 | ACTL6A, CLAR, CAV1, CITED2, COL1A1, COL1A2, FDXR, FUT1, HDAC1, HUWE1, JARID2, ENSA, EZH2 MCL1, MECP2, mir-196, NOTCH1, NTF4, PARP1, PRPF8 PIGER3, RCOR1, SERP1NB2, SF1, SIN3A, SMURF2, SNAI1, SP1, SSPN, TBX1, TP53, TP63, TRIM6, USP48, DNMT1 | 10 | 16 | Cellular Development, Organismal Development, Embryonic Development |
| 3 | ADAMTS3, ASPM, BIRC5, CCNB, CD44, DLGAP5 FOXM1, FOX01, HMMR, JAG1, JAK1, JUN, KIF18A, KLK6, LRP6, MAPK14, MITF, MYC, NCAPG, OSM, PBX1, PDX1, PGR, RBPJ, SERPINA3, SLC2A2, SPC25, TCF7L2, TFP1, TGFB1, TNC, TSPAN8, TXNIP, VEGA, EHF | 10 | 16 | Cancer, Cellular Growth and Proliferation Cellular Development |
| 4 | BID, CCL28, CCND2, CPT1A, CRCP, DSG2, HCAR3, ICAM1, IKBKE, IL18, ITGAV, JUN, MMD, MORF4L1, NAUK1, OAS1, PDPN, RRM2, TAP1, TGFB1, TGFBR2, TNF, VCAM1, VCL, WISP1, ZNF365 | 9 | 13 | Cell-To-Cell Signaling and Interaction, Hematological System Development and Function Inflammatory Response |
| 5 | ACTN4, ATF3, CCL2, CD40, CD80, CDC6, CDK6, CDKN2A, COL17A1, CXCL8, CXCL9, DST, E2F1, ESR2, FOS, HDAC3, IL13, 1L15, IL37, IL6, ITGA6, ITGB4, mir-181, NCOR2, NFKB1, NFKB1A, RAC1, RBI, RELA, SOD2, SP1 TERT, TLR9, TNSFL2, | 8 | 14 | Cancer, Cell Death and Survival Hematological Disease |
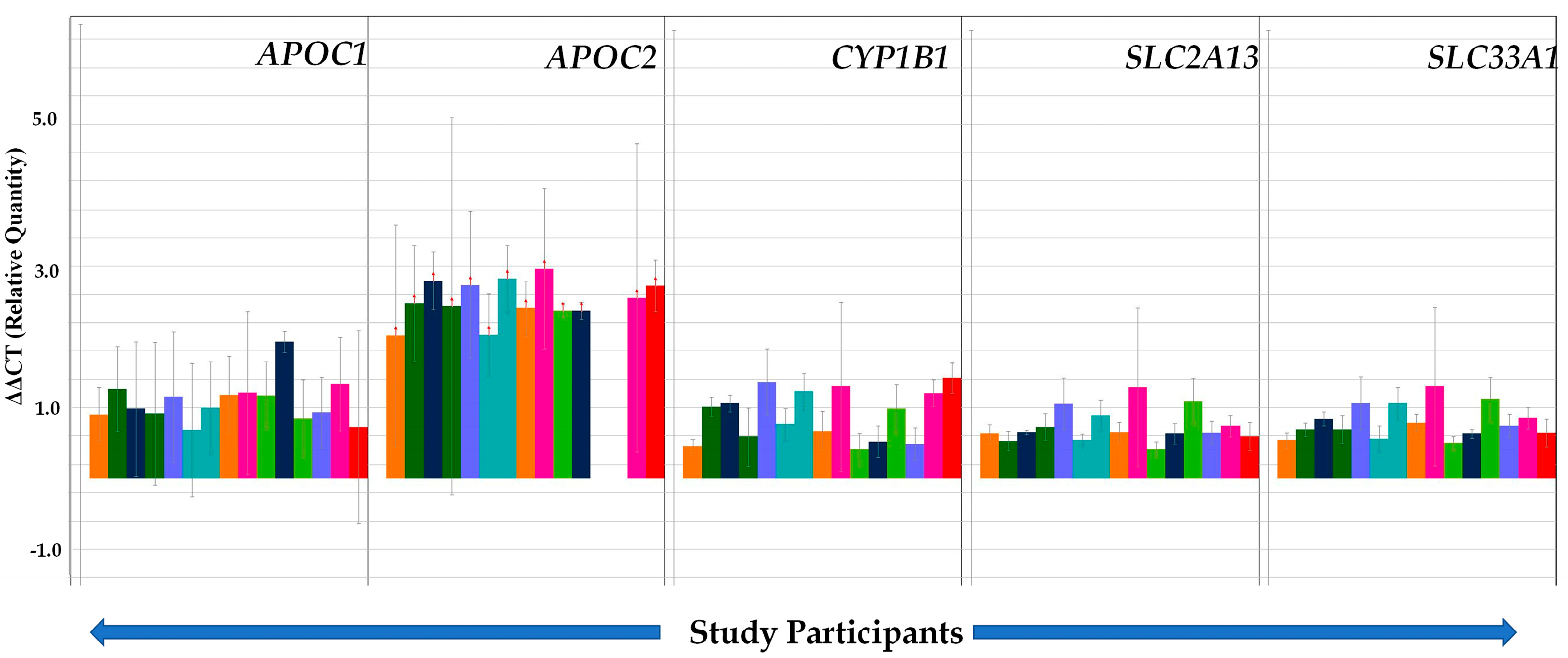
| Gene Name (Probe Sets) | Descriptions/Functions | Gene Regulation | % Change in Studied Subjects * (Number) | Average Relative Quantification ** |
|---|---|---|---|---|
| Metabolic Disease and Disorder | ||||
| LEPR (Hs00174492_m1) | Leptin receptor (Obesity) | Down | 7% (n = 1) | −0.35 |
| Up | 93% (n = 14) | +0.66 | ||
| RRAD (Hs00188163_m1) | Ras-related associated with diabetes | Down | 29% (n = 4) | −0.36+ 0.66 |
| Up | 71% (n = 11) | |||
| ARNT (Hs01121918_m1) | Encodes a protein that binds to ligand-bound aryl hydrocarbon receptor, involved in xenobiotic metabolism | Down | 20% (n = 3) | −0.25 |
| Up | 80% (n = 12) | +0.44 | ||
| Neurobehavioral | ||||
| CYP2D6 (Hs02576168_m1) | A member of Cytochrome P450 superfamily enzyme | Down | 20% (n = 3) | −0.27 |
| Up | 80% (n = 12) | +0.52 | ||
| APOC1 (Hs03037377_m1) | Apolipoprotein C1 Family; plays central role in HDL and VLDL metabolism | Up | 100% (n = 15) | +1.13 |
| APOC2(Hs000173442_m1) | Apolipoprotein C2 family that encodes a lipid-binding protein belonging to the apolipoprotein gene family, dysfunction or mutation results into hyperlipoproteinemia type IB, characterized by hypertriglyceridemia, xanthomas, and increased risk of pancreatitis and early atherosclerosis. | Up | 87% (n = 13) | +2.54 |
| ND | 13% (n = 2) | _ | ||
| CYP1B1(Hs00164385_m1) | Cytochrome P450 family 1 subfamily B member 1, which catalyzes many reactions involved in drug metabolism and synthesis of cholesterol, steroids, and other lipids. | Up | 100% (n = 15) | +1.12 |
| SLC2A13(Hs00369423_m1) | Solute carrier family 2 member 13, a member of mitochondrial carrier family | Up | 100% (n = 15) | +0.71 |
| SLC33A1(Hs00270469_m1) | Solute carrier family 33 member 1, required for the formation of O-acetylated (Ac) Gangliosides, disorder characterized by congenital cataracts, severe psychomotor retardation, and hearing loss | Up | 100% (n = 15) | +0.74 |
| Cancer | ||||
| MYC (Hs00153408_m1) | Proto-oncogene, cell cycle progression, apoptosis. | Down | 47% (n = 7) | −0.11 |
| Up | 47% (n = 7) | +0.54 | ||
| ND | 6% (n = 1) | - | ||
| TP53 (Hs01034249_m1) | Tumor suppressor protein p53 | Down | 73% (n = 11) | −0.27 |
| Up | 27% (n = 4) | +0.84 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noreen, Z.; Loffredo, C.A.; Bhatti, A.; Simhadri, J.J.; Nunlee-Bland, G.; Nnanabu, T.; John, P.; Khan, J.S.; Ghosh, S. Transcriptional Profiling and Biological Pathway(s) Analysis of Type 2 Diabetes Mellitus in a Pakistani Population. Int. J. Environ. Res. Public Health 2020, 17, 5866. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17165866
Noreen Z, Loffredo CA, Bhatti A, Simhadri JJ, Nunlee-Bland G, Nnanabu T, John P, Khan JS, Ghosh S. Transcriptional Profiling and Biological Pathway(s) Analysis of Type 2 Diabetes Mellitus in a Pakistani Population. International Journal of Environmental Research and Public Health. 2020; 17(16):5866. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17165866
Chicago/Turabian StyleNoreen, Zarish, Christopher A. Loffredo, Attya Bhatti, Jyothirmai J. Simhadri, Gail Nunlee-Bland, Thomas Nnanabu, Peter John, Jahangir S. Khan, and Somiranjan Ghosh. 2020. "Transcriptional Profiling and Biological Pathway(s) Analysis of Type 2 Diabetes Mellitus in a Pakistani Population" International Journal of Environmental Research and Public Health 17, no. 16: 5866. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17165866