A Review of Maternal Nutrition during Pregnancy and Impact on the Offspring through Development: Evidence from Animal Models of Over- and Undernutrition



Abstract
:1. Introduction
2. Evidence of Impacts of Maternal Nutrition on Epigenetic Marks in the Progeny
3. Impact of Maternal Nutrition on Fetal Programming in Animal Models
4. Summary and Implications for Human Health Disparities
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.; James, W.P.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the metabolic syndrome: A joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillen, I.C.; Robinson, J.S. Developmental origins of the metabolic syndrome: Prediction, plasticity, and programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef] [PubMed]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Bedogni, G.; Brambilla, B.; Cianfarani, S.; Nobili, V.; Pietrobelli, A.; Agostoni, C. Epigenetic mechanisms elicited by nutrition in early life. Nutr. Res. Rev. 2011, 24, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Lillycrop, K.A.; Burdge, G.C. Maternal diet as a modifier of offspring epigenetics. J. Dev. Orig. Health Dis. 2015, 6, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S. Impact of Maternal Diet on the Epigenome during In Utero Life and the Developmental Programming of Diseases in Childhood and Adulthood. Nutrients 2015, 7, 9492–9507. [Google Scholar] [CrossRef] [Green Version]
- Richards, E.J. Inherited epigenetic variation-revisiting soft inheritance. Nat. Rev. Genet. 2006, 2, 21–32. [Google Scholar] [CrossRef]
- Ulrey, C.L.; Liu, L.; Andrews, L.G.; Tollefsbol, T.O. The impact of metabolism on DNA methylation. Hum. Mol. Genet. 2005, 14 (Suppl. 1), R139–R147. [Google Scholar] [CrossRef]
- Waterland, R.A.; Lin, J.R.; Smith, C.A.; Jirtle, R.L. Postweaning diet affects genomic imprinting at the insulin-like growth factor 2 (Igf2) locus. Hum. Mol. Genet. 2006, 15, 705–716. [Google Scholar] [CrossRef] [Green Version]
- Waterland, R.A.; Cutbert, G. Potential mechanisms of metabolic imprinting that lead to chronic disease. Am. J. Clin. Nutr. 1999, 69, 179–197. [Google Scholar] [CrossRef]
- Ford, S.P.; Long, N.M. Evidence for similar changes in offspring phenotype following either maternal undernutrition or overnutrition: Potential impact on fetal epigenetic mechanisms. Reprod. Fertil. Dev. 2012, 24, 105–111. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A. Developmental origins of disease paradigm: A mechanistic and evolutionary perspective. Pediatr. Res. 2004, 56, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Dolinoy, D.C.; Wiedman, J.; Waterland, R.; Jirtle, R.L. Maternal genistein alters coat color and protects Avy mouse offspring from obesity by modifying the fetal epigenome. Environ. Health Perspect. 2006, 114, 567–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakyan, V.K.; Blewitt, M.E.; Druker, R.; Preis, J.I.; Whitelaw, E. Metastable epialleles in mammals. Trends Genet. 2002, 18, 348–351. [Google Scholar] [CrossRef]
- Waterland, R.A.; Jirtle, R.L. Transposable elements: Targets for early nutritional effects on epigenetic gene regulation. Mol. Cell. Biol. 2003, 23, 5293–5300. [Google Scholar] [CrossRef] [Green Version]
- Waterland, R.A.; Dolinoy, D.C.; Lin, J.R.; Smith, C.A.; Shi, X.; Tahiliani, K.G. Maternal methyl supplements increase offspring DNA methylation at axin fused. Genesis 2006, 44, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Wolff, G.L. Influence of maternal phenotype on metabolic differentiation of agouti locus mutants in the mouse. Genetics 1978, 88, 529–539. [Google Scholar]
- Morgan, H.D.; Sutherland, H.G.; Martin, D.I.; Whitelaw, E. Epigenetic inheritance at the agouti locus in the mouse. Nat. Genet. 1999, 23, 314–318. [Google Scholar] [CrossRef]
- Reed, S.C. The inheritance and expression of fused, a new mutation in the house mouse. Genetics 1937, 22, 1–13. [Google Scholar]
- Belyaev, D.K.; Ruvinsky, A.O.; Trut, L.N. Inherited activation–inactivation of the star gene in foxes: Its bearing on the problem of domestication. J. Hered. 1981, 72, 107–112. [Google Scholar] [CrossRef]
- Wolff, G.L.; Kodell, R.L.; Moore, S.R.; Cooney, C.A. Maternal epigenetics and methyl supplements affect agouti gene expression in Avy/a mice. FASEB J. 1998, 12, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Rakyan, V.K.; Chong, S.; Champ, M.E.; Cuthbert, P.C.; Morgan, H.D.; Luu, K.V.; Whitelaw, E. Transgenerational inheritance of epigenetic states at the murine AxinFu allele occurs after maternal and paternal transmission. Proc. Natl. Acad. Sci. USA 2003, A100, 2538–2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J. Fetal origins of coronary heart disease. BMJ 1993, 331, 171–174. [Google Scholar]
- Godfrey, K.; Robinson, S. Maternal nutrition, placental growth and fetal programming. Proc. Nutr. Soc. 1998, 57, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boney, C.M.; Verma, A.; Tucher, R.; Vohr, B.R. Metabolic syndrome in childhood: Association with birth weight, maternal obesity, and gestational diabetes mellitus. Pediatrics 2005, 115, e290–e296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mingrone, G.; Manco, M.; Mora, M.E.; Guidone, C.; Iaconelli, A.; Gnuili, D.; Leccesi, L.; Chiellini, C.; Ghirlanda, G. Influence of maternal obesity on insulin sensitivity and secretion in offspring. Diabetes Care 2008, 31, 1872–1876. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, R.C. Predicting preschooler obesity at birth: The role of maternal obesity in early pregnancy. Pediatrics 2004, 114, e29–e36. [Google Scholar] [CrossRef] [Green Version]
- Ford, S.P.; Zhang, L.; Zhu, M.; Miller, M.M.; Smith, D.T.; Hess, B.W.; Moss, G.E.; Nathanielsz, P.W.; Nijland, M.J. Maternal obesity accelerates fetal pancreatic beta-cell but not alpha-cell development in sheep: Prenatal consequences. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R835–R843. [Google Scholar] [CrossRef]
- Long, N.M.; George, L.A.; Uthlaut, A.B.; Smith, D.T.; Nijland, S.M.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity and increased nutrient intake before and during gestation in the ewe results in altered growth, adiposity, and glucose tolerance in adult offspring. J. Anim. Sci. 2010, 88, 3546–3553. [Google Scholar] [CrossRef] [Green Version]
- Vonnahme, K.A.; Hess, B.W.; Hansen, T.R.; McCormick, R.J.; Rule, D.C.; Moss, G.E.; Murdock, W.J.; Skinner, D.C.; Nathanielsz, P.W.; Ford, S.P. Maternal undernutrition from early to midgestation leads to growth retardation, cardiac ventricular hypertrophy, and increased liver weight in the fetal sheep. Biol. Reprod. 2003, 69, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Bouret, S.G. Early life origins of obesity: Role of hypothalamic programming. J. Pediatr. Gastroenterol. Nutr. 2009, 48, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Ergaz, Z.; Avgil, M.; Ornoy, A. Intrauterine growth restriction: Etiology and consequences: What do we know about the human situation and experimental animal models. Reprod. Toxicol. 2005, 20, 301–322. [Google Scholar] [CrossRef] [PubMed]
- Hales, C.N.; Barker, D.J. The thrifty phenotype hypothesis. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, S.P.; Hess, B.W.; Schwope, M.M.; Nijland, M.J.; Gilbert, J.S.; Vonnahme, K.A.; Means, W.J.; Han, H.; Nathanielsz, P.W. Maternal undernutrition during early to mid-gestation in the ewe results in altered growth, adiposity, and glucose tolerance in male offspring. J. Anim. Sci. 2007, 85, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Pankey, C.L.; Odhiambo, J.F.; Ghnenis, A.B.; Nathanielsz, P.W.; Ford, S.P. Rapid communication: Reduced maternal nutrition during early-to mid-gestation elevates newborn lamb plasma cortisol concentrations and eliminates the neonatal leptin surge. J. Anim. Sci. 2018, 96, 2640–2645. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhu, M.J.; Uthlaut, A.B.; Nijland, M.J.; Nathanielsz, P.W.; Hess, B.W.; Ford, S.P. Upregulation of growth signaling and nutrient transporters in the cotyledons of early to mid-gestational nutrient restricted ewes. Placenta 2011, 32, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Effect of maternal nutrient restriction in sheep on the development of fetal skeletal muscle. Biol. Reprod. 2004, 71, 1968–1973. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Ford, S.P.; Nijland, M.J.; Nathanielsz, P.W.; Ren, J. Influence of maternal undernutrition and overfeeding on cardiac ciliary neurotrophic factor receptor and ventricular size in fetal sheep. J. Nutr. Biochem. 2008, 19, 409–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Zhu, M.J.; Zhang, L.; Hein, S.M.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity and overnutrition alter fetal growth rate and cotyledonary vascularity and angiogenic factor expression in the ewe. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R249–R258. [Google Scholar] [CrossRef]
- Zhang, L.; George, L.A.; Ford, S.P.; Nathanielsz, P.W. Maternal nutrient restriction (NR) from early to mid-gestation increases pancreatic b-cell numbers at midgestation, but pancreatic weight and b-cell numbers are reduced by late gestation. J. Anim. Sci. 2010, 40, 30–39. [Google Scholar]
- Zhang, L.; Long, N.M.; Hein, S.M.; Ma, Y.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity in ewes results in reduced fetal pancreatic b-cell numbers in late gestation and decreased circulating insulin concentration at term. Domest. Anim. Endocrinol. 2011, 40, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Ford, S.P.; Fang, C.X.; Nijland, M.J.; Nathanielsz, P.W.; Ren, J. Maternal nutrient restriction during early to mid gestation up-regulates cardiac insulin-like growth factor (IGF) receptors associated with enlarged ventricular size in fetal sheep. Grow. Horm. IGF Res. 2005, 4, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.J.; Ma, Y.; Long, N.M.; Du, M.; Ford, S.P. Maternal obesity markedly increases placental fatty acid transporter expression and fetal blood triglycerides at midgestation in the ewe. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2010, 299, R1224–R1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ma, H.; Tong, C.; Zhang, H.; Lawlis, G.B.; Li, Y.; Zang, M.; Ren, J.; Nijland, M.J.; Ford, S.P.; et al. Overnutrition and maternal obesity in sheep pregnancy alter the JNK IRS-1 signaling cascades and cardiac function in the fetal heart. FASEB J. 2010, 24, 2066–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, N.M.; Ford, S.P.; Nathanielsz, P.W. Maternal obesity eliminates the neonatal lamb plasma leptin peak. J. Physiol. 2011, 589, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- George, L.A.; Zhang, L.; Tuersunjiang, N.; Ma, Y.; Long, N.M.; Uthlaut, A.B.; Smith, D.T.; Nathanielsz, P.W.; Ford, S.P. Early maternal undernutrition programs increased feed intake, altered glucose metabolism and insulin secretion, and liver function in aged female offspring. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2012, 302, R795–R804. [Google Scholar] [CrossRef] [Green Version]
- Long, N.M.; Rule, D.C.; Tuersunjiang, N.; Nathanielsz, P.W.; Ford, S.P. Maternal Obesity in Sheep Increases Fatty Acid Synthesis, Upregulates Nutrient Transporters, and Increases Adiposity in Adult Male Offspring after a Feeding Challenge. PLoS ONE 2015, 10, e0122152. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ford, S.P.; Means, W.J.; Hess, B.W.; Nathanielsz, P.W.; Du, M. Maternal nutrient restriction affects properties of skeletal muscle in offspring. J. Physiol. 2006, 575, 241–250. [Google Scholar] [CrossRef]
- Yan, X.; Huang, Y.; Zhao, J.X.; Long, N.M.; Uthlaut, A.B.; Zhu, M.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Maternal obesity-impaired insulin signaling in sheep and induced lipid accumulation and fibrosis in skeletal muscle of offspring. Biol. Reprod. 2011, 85, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Zhu, M.J.; Xu, W.; Tong, J.F.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Up-regulation of Toll-like receptor 4/nuclear factor-kappa B signaling is associated with enhanced adipogenesis and insulin resistance in fetal skeletal muscle of obese sheep at late gestation. Endocrinology 2010, 151, 380–387. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, J.X.; Yan, X.; Zhu, M.J.; Long, N.M.; McCormick, R.J.; Ford, S.P. Maternal obesity enhances collagen accumulation and cross-linking in skeletal muscle of ovine offspring. PLoS ONE 2012, 7, e31691. [Google Scholar] [CrossRef] [Green Version]
- Moisiadis, V.G.; Matthews, S.G. Glucocorticoids and fetal programming part 2: Mechanisms. Nat. Rev. Endocrinol. 2014, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Ghnenis, A.B.; Odhiambo, J.F.; McCormick, R.J.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity in the ewe increases cardiac ventricular expression of glucocorticoid receptors, proinflammatory cytokines and fibrosis in adult male offspring. PLoS ONE 2017, 12, 12. [Google Scholar] [CrossRef]
- Ghnenis, A.B.; Odhiambo, J.F.; Smith, A.M.; Pankey, C.L.; Nathanielsz, P.W.; Ford, S.P. A heretical view: Rather than a solely placental protective function, placental 11β hydroxysteroid dehydrogenase 2 also provides substrate for fetal peripheral cortisol synthesis in obese pregnant ewes. J. Dev. Orig. Health Dis. 2020, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Busada, J.T.; Cidlowski, J.A. Mechanisms of glucocorticoid action during development. In Current Topics in Developmental Biology; Forrest, D., Tsai, S., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 125, pp. 147–170. [Google Scholar]
- Shasa, D.R.; Odhiambo, J.F.; Long, N.M.; Tuersunjiang, N.; Nathanielsz, P.W.; Ford, S.P. Multigenerational impact of maternal overnutrition/obesity in the sheep on the neonatal leptin surge in granddaughters. Int. J. Obes. 2015, 39, 695–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Zhu, C.; Sun, M.; Maimaiti, R.; Ford, S.P.; Nathanielsz, P.W.; Ren, J.; Guo, W. Maternal obesity impairs fetal cardiomyocyte contractile function in sheep. FASEB J. 2019, 33, 2587–2598. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Yan, X.; Zhao, J.X.; Zhu, M.J.; McCormick, R.J.; Ford, S.P.; Nathanielsz, P.W.; Ren, J.; Du, M. Maternal obesity induces fibrosis in fetal myocardium of sheep. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E968–E975. [Google Scholar] [CrossRef]
- Fan, X.; Turdi, S.; Ford, S.P.; Hua, Y.; Nijland, M.J.; Zhu, M.; Nathanielsz, P.W.; Ren, J. Influence of gestational overfeeding on cardiac morphometry and hypertrophic protein markers in fetal sheep. J. Nutr. Biochem. 2011, 22, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Pankey, C.L.; Walton, M.W.; Odhiambo, J.F.; Smith, A.M.; Ghnenis, A.B.; Nathanielsz, P.W.; Ford, S.P. Intergenerational impact of maternal overnutrition and obesity throughout pregnancy in sheep on metabolic syndrome in grandsons and granddaughters. Domest. Anim. Endocrinol. 2017, 60, 67–74. [Google Scholar] [CrossRef]
- Deputy, N.P.; Dub, B.; Sharma, A.J. Prevalence and Trends in Prepregnancy Normal Weight—48 States, New York City, and District of Columbia, 2011–2015. MMWR Morb. Mortal. Wkly. Rep. 2018, 66, 1402–1407. [Google Scholar] [CrossRef] [Green Version]



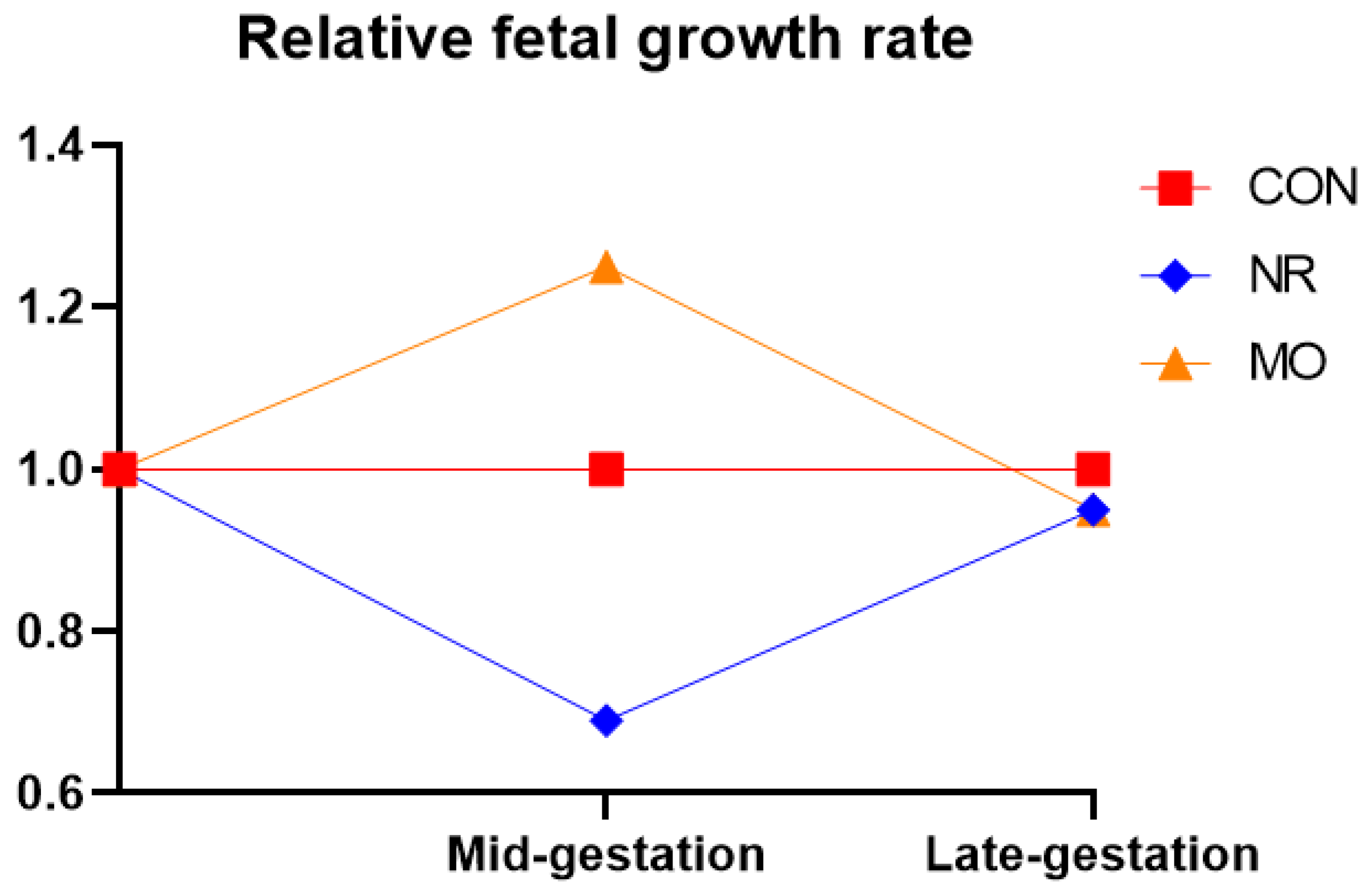{kind=link}
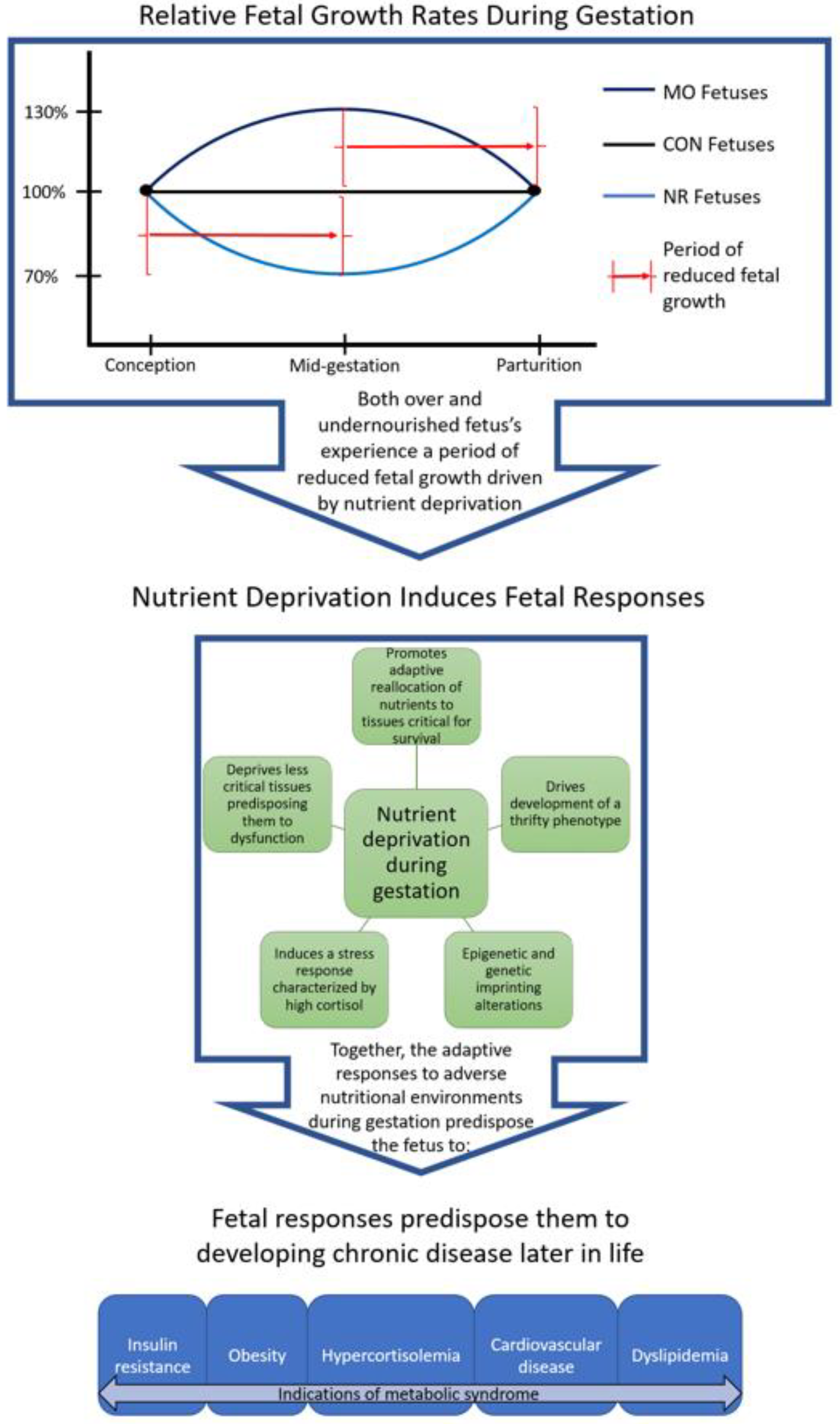{kind=link}
| Mid−Gestation (Day 78) Fetus | NR Offspring | MO Offspring | Reference(s) |
|---|---|---|---|
| − fetal weight | − | + | [29,35,36,37],[37,38] |
| − crown rump length | − | + | [29,35,37],[37,38] |
| − liver wt | + | + | [29],[27] |
| − pancreatic β−cell numbers | + | + | [39],[27,40] |
| − cardiac ventricular wt/fetal wt | + | + | [41],[37] |
| − plasma glucose | − | + | [29],[27] |
| − plasma insulin | ND | + | [N/A],[27] |
| − plasma cortisol | + | + | [37],[27,37] |
| − plasma cholesterol | + | + | [35],[42] |
| − plasma triglycerides | − | + | [35],[42] |
| Late−gestation (Day 135) fetus | |||
| − fetal wt | ND | ND | [38],[38] |
| − crown rump length | ND | ND | [38],[38] |
| − liver wt | ND | ND | [41],[40] |
| − pancreatic β−cell numbers | − | − | [39],[40] |
| − cardiac ventricular wt./fetal wt | + | + | [41],[43] |
| − plasma glucose | ND | [N/A],[40] | |
| − plasma insulin | ND | [N/A],[40] | |
| − plasma cortisol | ND | [N/A],[40] | |
| − plasma cholesterol | ND | + | [35],[42] |
| − plasma triglycerides | − | + | [35],[42] |
| Newborn lambs | |||
| − birth weight | ND | ND | [41],[27,44] |
| − crown rump length | ND | [N/A],[27] | |
| − plasma glucose | + | + | [34],[27] |
| − plasma insulin | − | − | [34],[44] |
| − plasma cortisol | + | + | [34],[44] |
| − plasma leptin | − | − | [34],[44] |
| Adult offspring | |||
| − appetite | + | + | [45],[46] |
| − growth rate | + | + | [33],[46] |
| − insulin resistance | + | + | [33,45],[46] |
| − plasma leptin | + | + | [45],[46] |
| − adiposity to ad. lib. feeding | + | + | [45],[46] |
| − left ventricular wall thickness | + | + | [45],[46] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Odhiambo, J.F.; Pankey, C.L.; Ghnenis, A.B.; Ford, S.P. A Review of Maternal Nutrition during Pregnancy and Impact on the Offspring through Development: Evidence from Animal Models of Over- and Undernutrition. Int. J. Environ. Res. Public Health 2020, 17, 6926. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17186926
Odhiambo JF, Pankey CL, Ghnenis AB, Ford SP. A Review of Maternal Nutrition during Pregnancy and Impact on the Offspring through Development: Evidence from Animal Models of Over- and Undernutrition. International Journal of Environmental Research and Public Health. 2020; 17(18):6926. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17186926
Chicago/Turabian StyleOdhiambo, John F., Christopher L. Pankey, Adel B. Ghnenis, and Stephen P. Ford. 2020. "A Review of Maternal Nutrition during Pregnancy and Impact on the Offspring through Development: Evidence from Animal Models of Over- and Undernutrition" International Journal of Environmental Research and Public Health 17, no. 18: 6926. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17186926