Enhancement of Chromium (VI) Reduction in Microcosms Amended with Lactate or Yeast Extract: A Laboratory-Scale Study


 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Matrices
2.2. Amendments
2.3. Experimental Design and Microcosms Setup
2.4. Controls
2.5. Chemical Analysis
2.6. Molecular Analysis
2.6.1. Evaluation of Microbial Abundance
2.6.2. PCR Assays
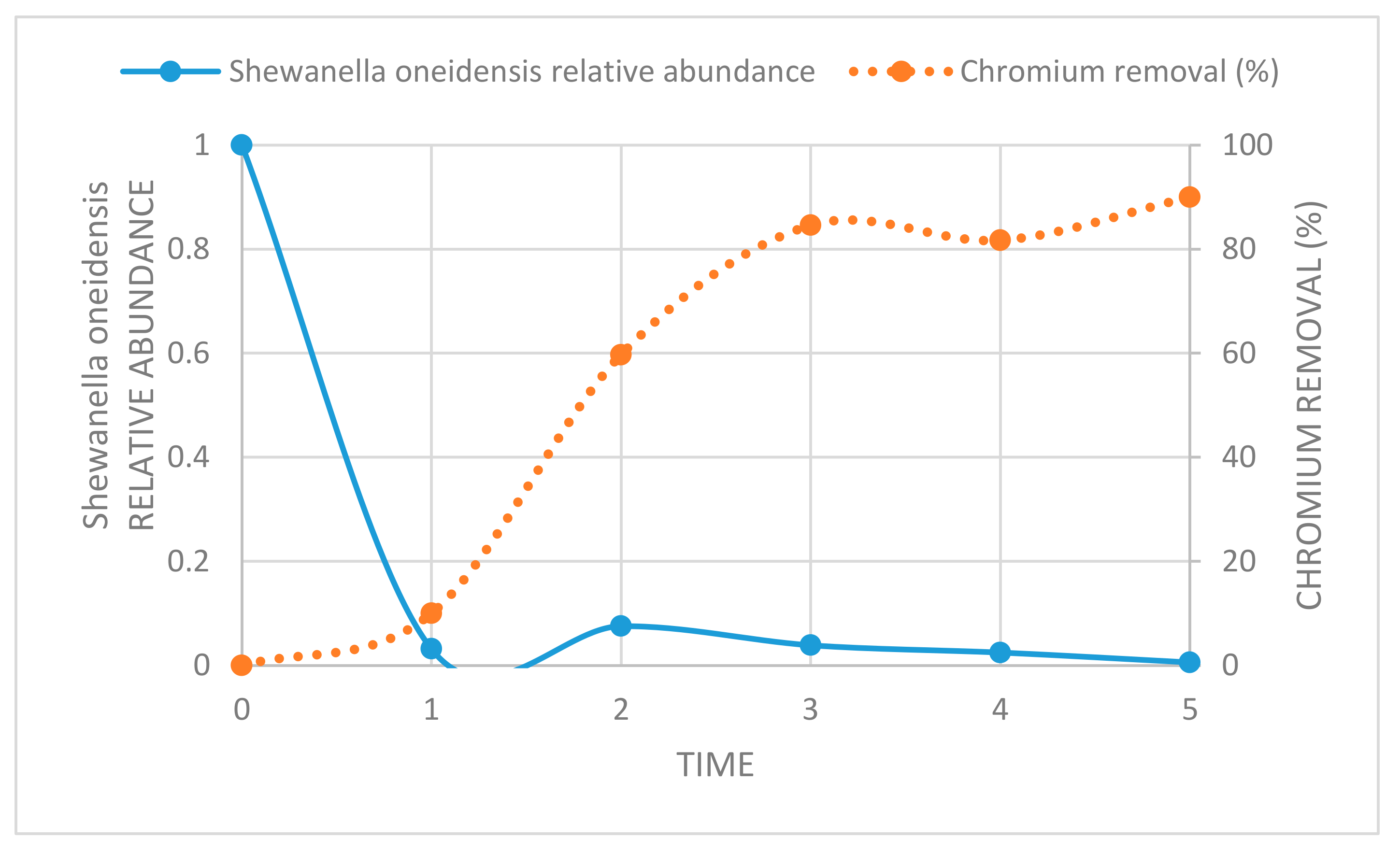
2.6.3. Assessment of Shewanella Oneidensis in Amended Microcosms
3. Results
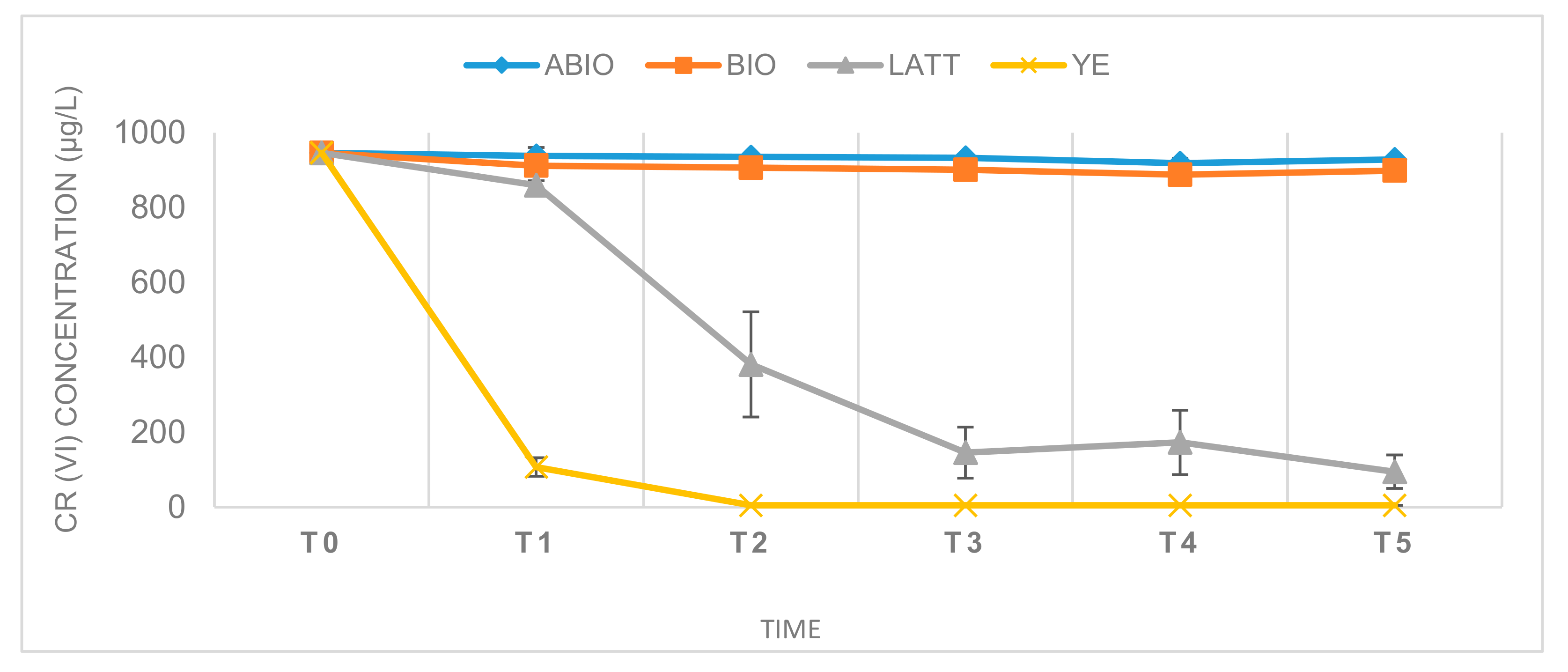
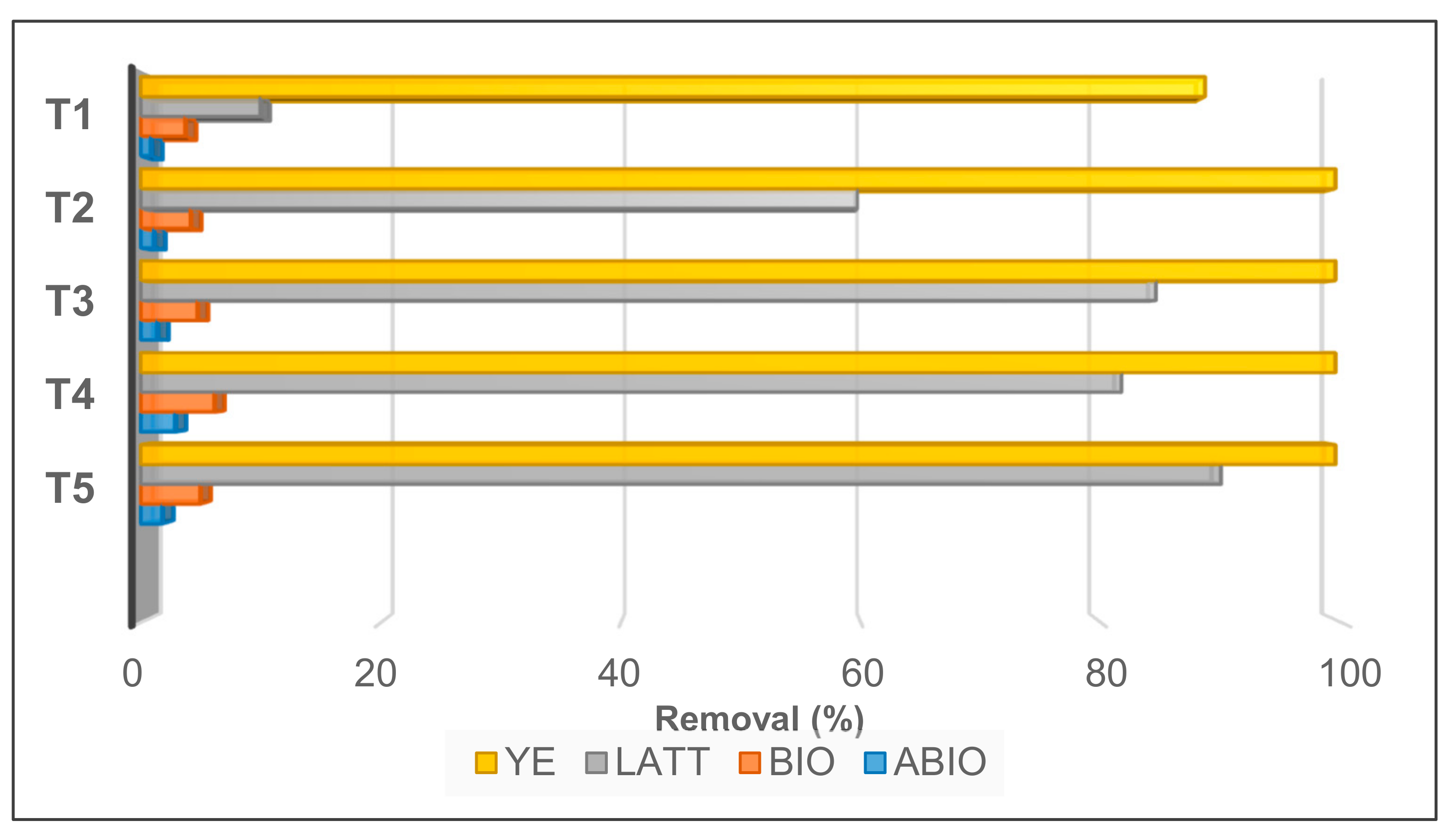
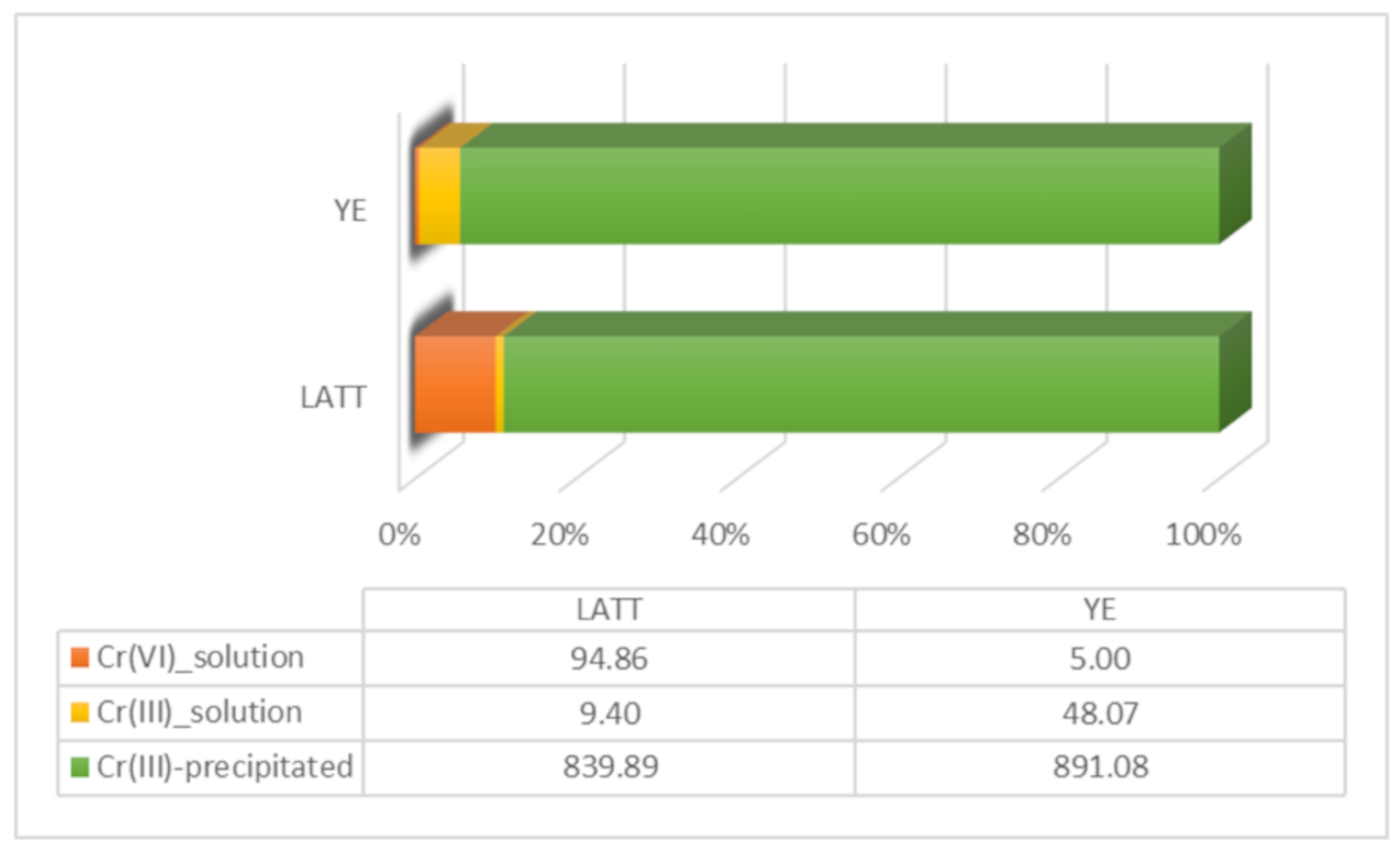
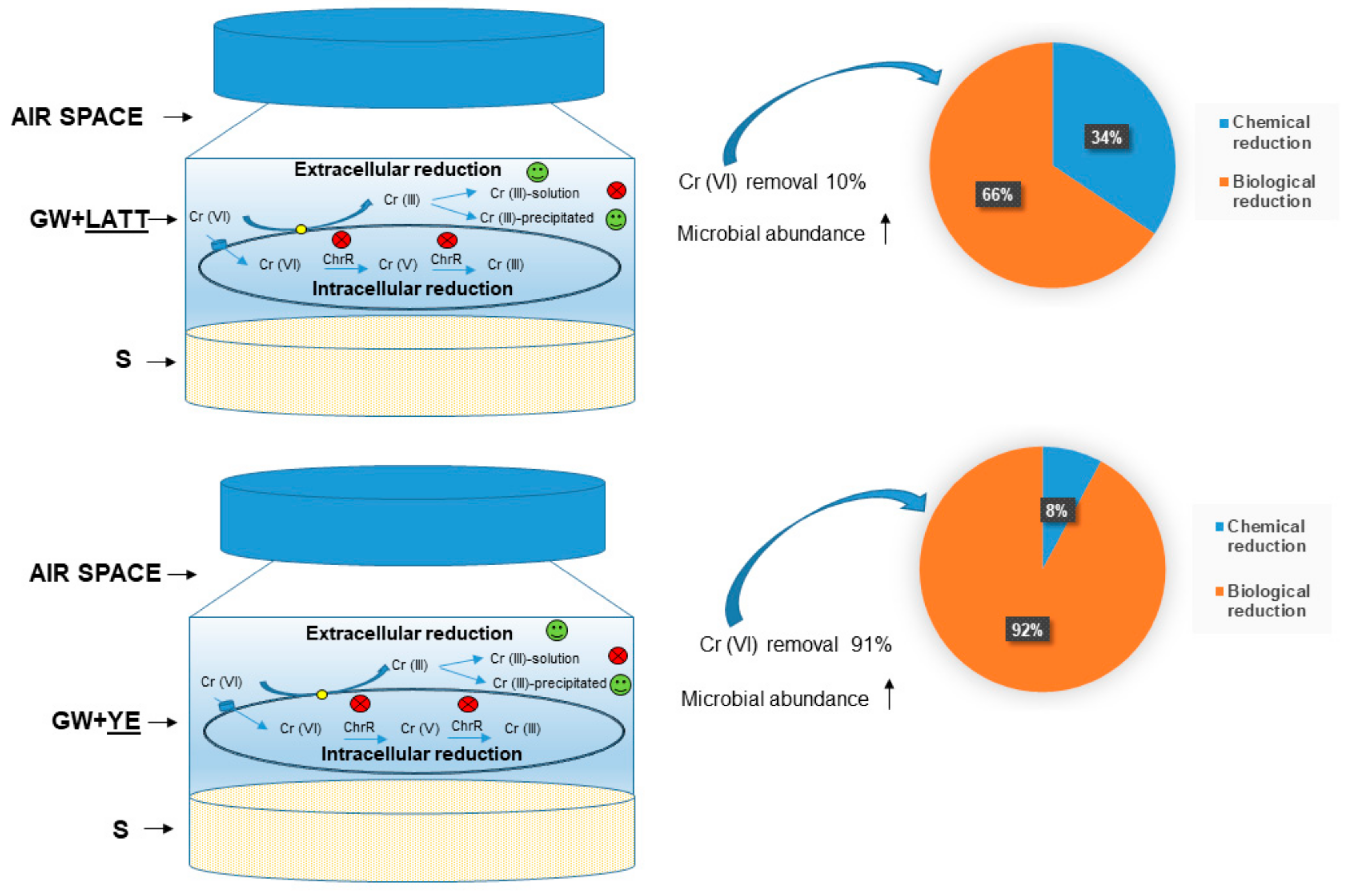
3.1. Cr(VI) Reduction and Distribution Mass-Balance
3.2. Control Tests
3.3. The Amendments’ Effect on Microbial Abundance
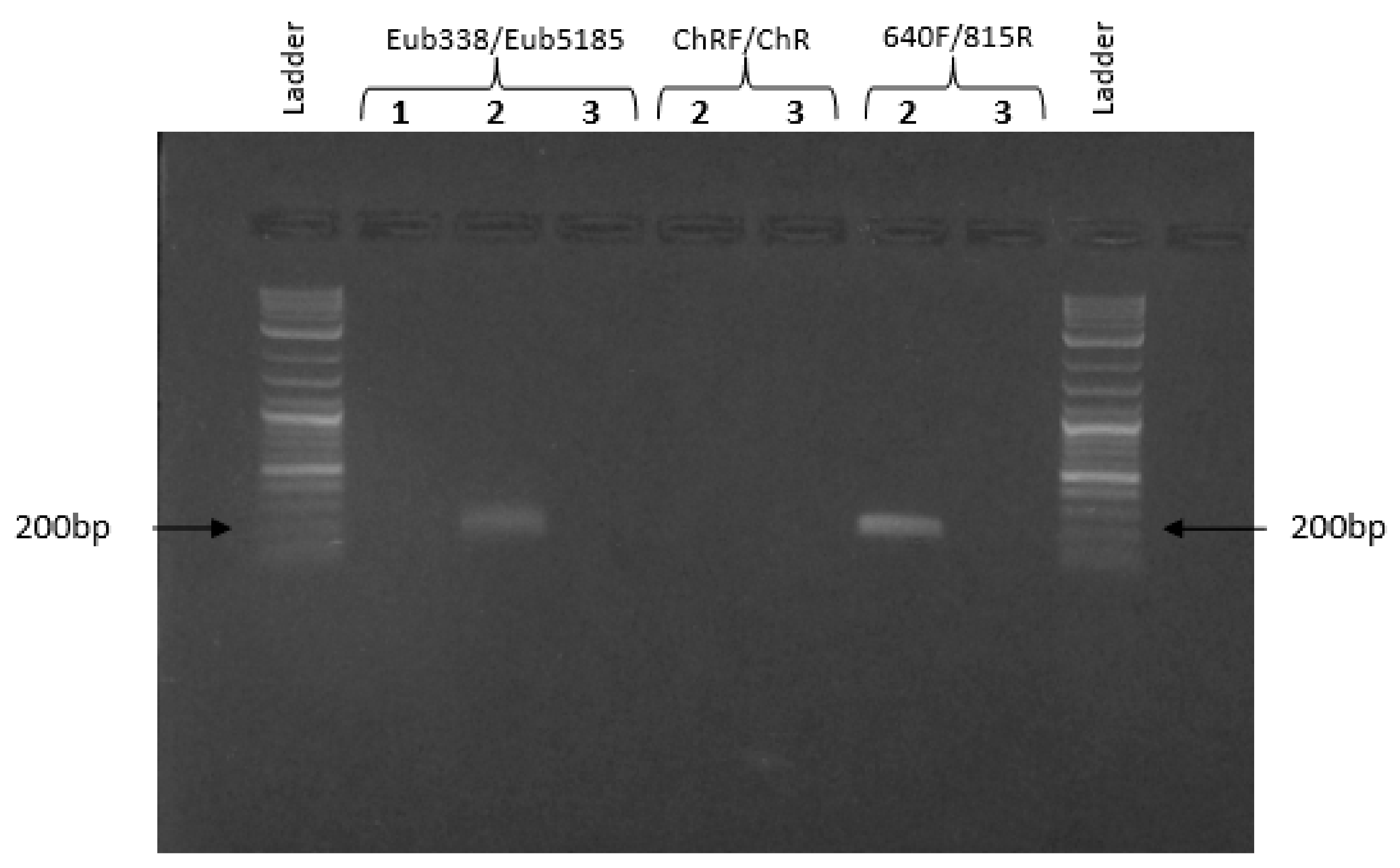
3.4. PCR Assays
3.5. Assessment of Shewanella Oneidensis in Amended Biological Microcosm
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kimbrough, D.E.; Cohen, Y.; Winer, A.M.; Creelman, L.; Mabuni, C. A critical assessment of chromium in the environment. Crit. Rev. Environ. Sci. Technol. 1999, 29, 1–46. [Google Scholar] [CrossRef]
- Saha, B.; Orvig, C. Biosorbents for hexavalent chromium elimination from industrial and municipal effluents. Coord. Chem. Rev. 2010, 254, 2959–2972. [Google Scholar] [CrossRef]
- Codd, R.; Dillon, C.T.; Levina, A.; Lay, P.A. Studies on the genotoxicity of chromium: From the test tube to the cell. Coord. Chem. Rev. 2001, 217, 537–582. [Google Scholar] [CrossRef]
- Focardi, S.; Pepi, M.; Focardi, E.S. Microbial Reduction of Hexavalent Chromium as a Mechanism of Detoxification and Possible Bioremediation Applications. In Biodegradation—Life of Science; InTechOpen: London, UK, 2013. [Google Scholar]
- Koleli, N.; Demir, A. Chromite. In Environmental Materials and Waste: Resource Recovery and Pollution Prevention; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 245–263. [Google Scholar]
- Liu, H.; Wang, Y.; Zhang, H.; Huang, G.; Yang, Q.; Wang, Y. Synchronous detoxification and reduction treatment of tannery sludge using Cr (VI) resistant bacterial strains. Sci. Total Environ. 2019, 687, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Mtimunye, P.J.; Lutsinge, T.B.; Molokwane, P.E.; Chirwa, E.M.N. Cr(VI) remediation in groundwater aquifer media using natural organic matter as carbon source. Chem. Eng. Trans. 2017, 61, 1831–1836. [Google Scholar]
- Belchik, S.M.; Kennedy, D.W.; Dohnalkova, A.C.; Wang, Y.; Sevinc, P.C.; Wu, H.; Lin, Y.; Lu, H.P.; Fredrickson, J.K.; Shi, L. Extracellular reduction of hexavalent chromium by cytochromes MtrC and OmcA of Shewanella oneidensis MR-1. Appl. Environ. Microbiol. 2011, 77, 4035–4041. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, M.A.; Bar, H.Y.; Beg, Q.K.; Segrè, D.; Booth, J.; Cotta, M.A.; Angenent, L.T. Transcriptional analysis of shewanella oneidensis MR-1 with an electrode compared to Fe(III)citrate or oxygen as terminal electron acceptor. PLoS ONE 2012, 7, e30827. [Google Scholar] [CrossRef]
- Baaziz, H.; Gambari, C.; Boyeldieu, A.; Chaouche, A.A.; Alatou, R.; Méjean, V.; Jourlin-Castelli, C.; Fons, M. ChrASO, the chromate efflux pump of Shewanella oneidensis, improves chromate survival and reduction. PLoS ONE 2017, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Chen, N.; Feng, C.; Li, M.; Gao, Y.; Hu, Y. Coupling enhancement of Chromium (VI) bioreduction in groundwater by phosphorus minerals. Chemosphere 2020, 240, 124896. [Google Scholar] [CrossRef]
- Thatoi, H.; Das, S.; Mishra, J.; Rath, B.P.; Das, N. Bacterial chromate reductase, a potential enzyme for bioremediation of hexavalent chromium: A review. J. Environ. Manag. 2014, 146, 383–399. [Google Scholar] [CrossRef]
- Zhu, W.; Chai, L.; Ma, Z.; Wang, Y.; Xiao, H.; Zhao, K. Anaerobic reduction of hexavalent chromium by bacterial cells of Achromobacter sp. Strain Ch1. Microbiol. Res. 2008, 163, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Li, X. Bioreduction of chromium (VI) by Bacillus sp. isolated from soils of iron mineral area. Eur. J. Soil Biol. 2009, 45, 483–487. [Google Scholar] [CrossRef]
- Bharagava, R.N.; Mishra, S. Hexavalent chromium reduction potential of Cellulosimicrobium sp. isolated from common effluent treatment plant of tannery industries. Ecotoxicol. Environ. Saf. 2018, 147, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chai, L.; Liao, Q.; Tang, C.; Liao, Y.; Peng, B.; Yang, Z. Structural and genetic diversity of hexavalent chromium-resistant bacteria in contaminated soil. Geomicrobiol. J. 2016, 33, 222–229. [Google Scholar] [CrossRef]
- Barra Caracciolo, A.; Bottoni, P.; Grenni, P. Microcosm studies to evaluate microbial potential to degrade pollutants in soil and water ecosystems. Microchem. J. 2013, 107, 126–130. [Google Scholar] [CrossRef]
- Uricchio, V.F.; Calabrese, A.; Massarelli, C.; Campanale, C.; De Paola, D.; Rapanà, N.; Ciannarella, R.; Casale, B.; Tumolo, M.; Mauro, R. Protocollo D’intesa per la Realizzazione di un Monitoraggio Ambientale nel Territorio Della Città di Barletta. Relazione Tecnica Conclusiva; IRSA-CNR Internal Report within the Project: “Environmental Monitoring of Barletta Land.”; IRSA: Roma, Italia, 2017; pp. 1–139. (In Italian) [Google Scholar]
- McLean, J.; Beveridge, T.J. Chromate Reduction by a Pseudomonad Isolated from a Site Contaminated with Chromated Copper Arsenate Chromate Reduction by a Pseudomonad Isolated from a Site Contaminated with Chromated Copper Arsenate. Appl. Environ. Microbiol. 2001, 67, 1076–1084. [Google Scholar] [CrossRef] [Green Version]
- Somenahally, A.C.; Mosher, J.J.; Yuan, T.; Podar, M.; Phelps, T.J.; Brown, S.D.; Yang, Z.K.; Hazen, T.C.; Arkin, A.P.; Palumbo, A.V.; et al. Hexavalent chromium reduction under fermentative conditions with lactate stimulated native microbial communities. PLoS ONE 2013, 8, 1–11. [Google Scholar] [CrossRef]
- Cifuentes, F.R.; Lindemann, W.C.; Barton, L.L. Chromium sorption and reduction in soil with implications to bioremediation. Soil Sci. 1996, 161, 233–241. [Google Scholar] [CrossRef]
- Matteucci, F.; Ercole, C.; Del Gallo, M. A study of chlorinated solvent contamination of the aquifers of an industrial area in central Italy: A possibility of bioremediation. Front. Microbiol. 2015, 6, 1–10. [Google Scholar] [CrossRef]
- Volpe, A.; Del Moro, G.; Rossetti, S.; Tandoi, V.; Lopez, A. Remediation of PCE-contaminated groundwater from an industrial site in southern Italy: A laboratory-scale study. Process Biochem. 2007, 42, 1498–1505. [Google Scholar] [CrossRef]
- Chen, Z.F.; Zhao, Y.S.; Li, Q. Characteristics and kinetics of hexavalent chromium reduction by gallic acid in aqueous solutions. Water Sci. Technol. 2015, 71, 1694–1700. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, J.A.; Vilgalys, R.; Jackson, R.B. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl. Environ. Microbiol. 2005, 71, 4117–4120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, R.C.; Malik, S.; Beer, M.; Megharaj, M.; Naidu, R. Molecular characterization of chromium (VI) reducing potential in Gram positive bacteria isolated from contaminated sites. Soil Biol. Biochem. 2010, 42, 1857–1863. [Google Scholar] [CrossRef]
- Li, B.B.; Cheng, Y.Y.; Fan, Y.Y.; Liu, D.F.; Fang, C.Y.; Wu, C.; Li, W.W.; Yang, Z.C.; Yu, H.Q. Estimates of abundance and diversity of Shewanella genus in natural and engineered aqueous environments with newly designed primers. Sci. Total Environ. 2018, 637–638, 926–933. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bodek, I.; Lyman, W.J.; Reehl, W.F.; Rosenblatt, D.H. Environmental Inorganic Chemistry, 1st ed.; Elsevier: Amsterdam, The Netherlands, 1988. [Google Scholar]
- Rai, D.; Eary, L.E.; Zachara, J.M. Environmental chemistry of chromium. Sci. Total Environ. 1989, 86, 15–23. [Google Scholar] [CrossRef]
- Gong, Y.; Werth, C.J.; He, Y.; Su, Y.; Zhang, Y.; Zhou, X. Intracellular versus extracellular accumulation of Hexavalent chromium reduction products by Geobacter sulfurreducens PCA. Environ. Pollut. 2018, 240, 485–492. [Google Scholar] [CrossRef]
- Ksheminska, H.; Fedorovych, D.; Honchar, T.; Ivash, M.; Gonchar, M. Yeast tolerance to chromium depends on extracellular chromate reduction and Cr(III) chelation. Food Technol. Biotechnol. 2008, 46, 419–426. [Google Scholar]
- Mary Mangaiyarkarasi, M.S.; Vincent, S.; Janarthanan, S.; Subba Rao, T.; Tata, B.V.R. Bioreduction of Cr(VI) by alkaliphilic Bacillus subtilis and interaction of the membrane groups. Saudi J. Biol. Sci. 2011, 18, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Long, D.; Tang, X.; Cai, K.; Chen, G.; Chen, L.; Duan, D.; Zhu, J.; Chen, Y. Cr(VI) reduction by a potent novel alkaliphilic halotolerant strain Pseudochrobactrum saccharolyticum LY10. J. Hazard. Mater. 2013, 256–257, 24–32. [Google Scholar] [CrossRef]
- Viamajala, S.; Peyton, B.M.; Sani, R.K.; Apel, W.A.; Petersen, J.N. Toxic Effects of Chromium (VI) on Anaerobic and Aerobic Growth of Shewanella oneidensis MR-1. Biotechnol. Prog. 2004, 20, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Middleton, S.S.; Latmani, R.B.; Mackey, M.R.; Ellisman, M.H.; Tebo, B.M.; Criddle, C.S. Cometabolism of Cr(VI) by Shewanella oneidensis MR-1 produces cell-associated reduced chromium and inhibits growth. Biotechnol. Bioeng. 2003, 83, 627–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, K.H.; Gu, J.D. Mechanism of hexavalent chromium detoxification by microorganisms and bioremediation application potential: A review. Int. Biodeterior. Biodegrad. 2007, 59, 8–15. [Google Scholar] [CrossRef]
- Qian, J.; Wei, L.; Liu, R.; Jiang, F.; Hao, X.; Chen, G.H. An exploratory study on the pathways of Cr (VI) reduction in sulfate-reducing Up-flow Anaerobic Sludge Bed (UASB) reactor. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Xu, J.; Chen, N.; Li, M.; Feng, C. Microbial reduction fate of chromium (Cr) in aqueous solution by mixed bacterial consortium. Ecotoxicol. Environ. Saf. 2019, 170, 763–770. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Condition | Composition | Properties |
|---|---|---|
| ABIO | S + GW + sodium azide (1 g/L) | Autoclaved matrices with sodium azide |
| BIO | S + GW | Not autoclaved matrices |
| LATT | S + GW + lactate (3 mM) | Not autoclaved matrices amended with lactate |
| YE | S + GW + yeast extract (200 mg/L) | Not autoclaved matrices amended with yeast extract |
| Method Used | Sequence Primers | Amplicon Length | Genome | Reference |
|---|---|---|---|---|
| Eub338 | ACTCCTACGGGAGGCAGCAG | 200 bp | 16S rRNA gene All of the bacteria | [25] |
| Eub518 | ATTACCGCGGCTGCTGG | |||
| ChR | CGTACCCTGATCAATCACTT | 268 bp | Chromate reductase gene | [26] |
| ChRF | TCACGCCGGAATATAACTAC | |||
| 640F | RACTAGAGTCTTGTAGAGG | 195 bp | V3–V4 Shewanella oneidensis | [27] |
| 815R | AAGDYACCAAAYTCCGAGTA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ancona, V.; Campanale, C.; Tumolo, M.; De Paola, D.; Ardito, C.; Volpe, A.; Uricchio, V.F. Enhancement of Chromium (VI) Reduction in Microcosms Amended with Lactate or Yeast Extract: A Laboratory-Scale Study. Int. J. Environ. Res. Public Health 2020, 17, 704. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17030704
Ancona V, Campanale C, Tumolo M, De Paola D, Ardito C, Volpe A, Uricchio VF. Enhancement of Chromium (VI) Reduction in Microcosms Amended with Lactate or Yeast Extract: A Laboratory-Scale Study. International Journal of Environmental Research and Public Health. 2020; 17(3):704. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17030704
Chicago/Turabian StyleAncona, Valeria, Claudia Campanale, Marina Tumolo, Domenico De Paola, Claudio Ardito, Angela Volpe, and Vito Felice Uricchio. 2020. "Enhancement of Chromium (VI) Reduction in Microcosms Amended with Lactate or Yeast Extract: A Laboratory-Scale Study" International Journal of Environmental Research and Public Health 17, no. 3: 704. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17030704