Changes in Microbial Community Structures under Reclaimed Water Replenishment Conditions


Abstract
:1. Introduction
2. Materials and Methods
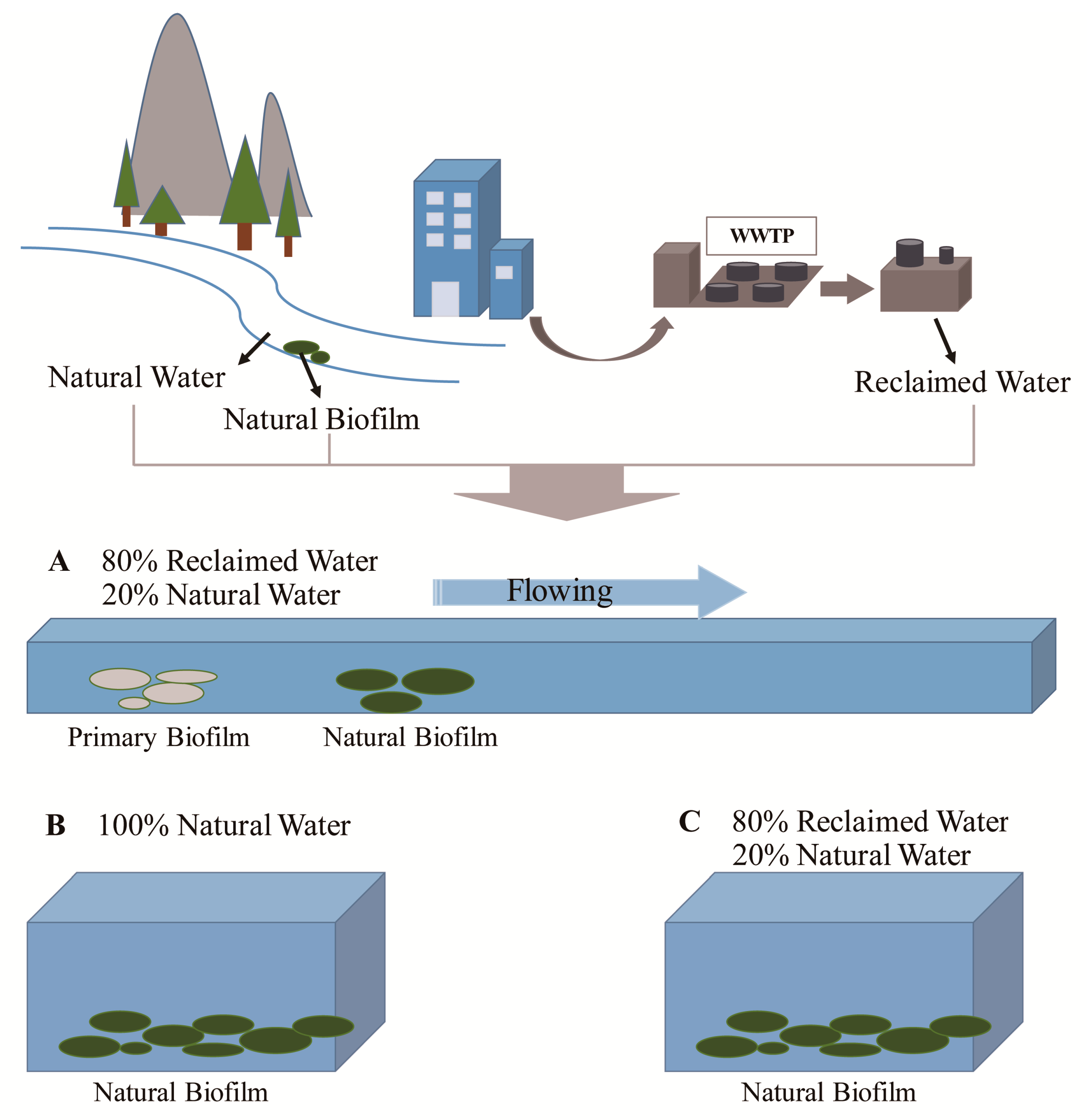
2.1. Water Resources for the Experiment
2.2. Experimental Design
2.3. Sample Collection and Preprocessing
2.4. Water Quality and Analytical Methods
2.5. Genomic DNA Extraction and High-Throughput Sequencing
2.6. Statistical Analysis
3. Results and Discussion
3.1. Water Quality
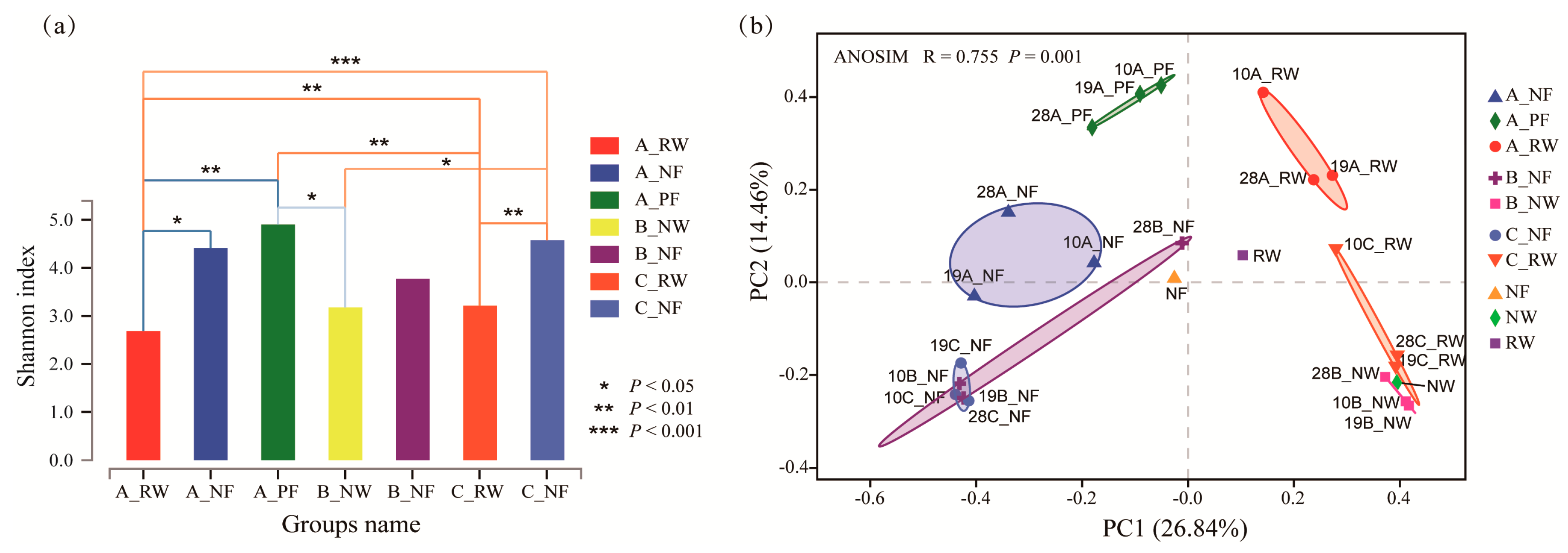
3.2. Sequencing Assessment and Taxonomic Diversity
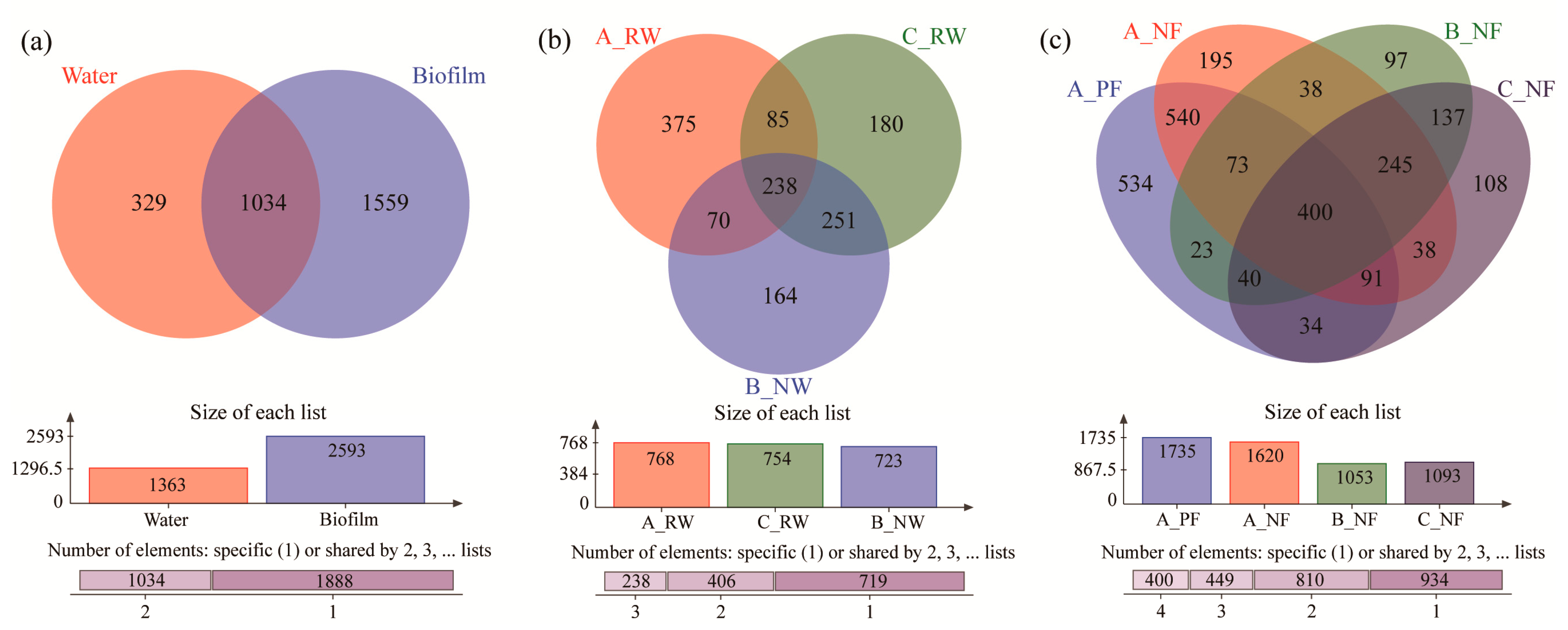
3.3. Common and Unique Microbial Taxa of Different Groups
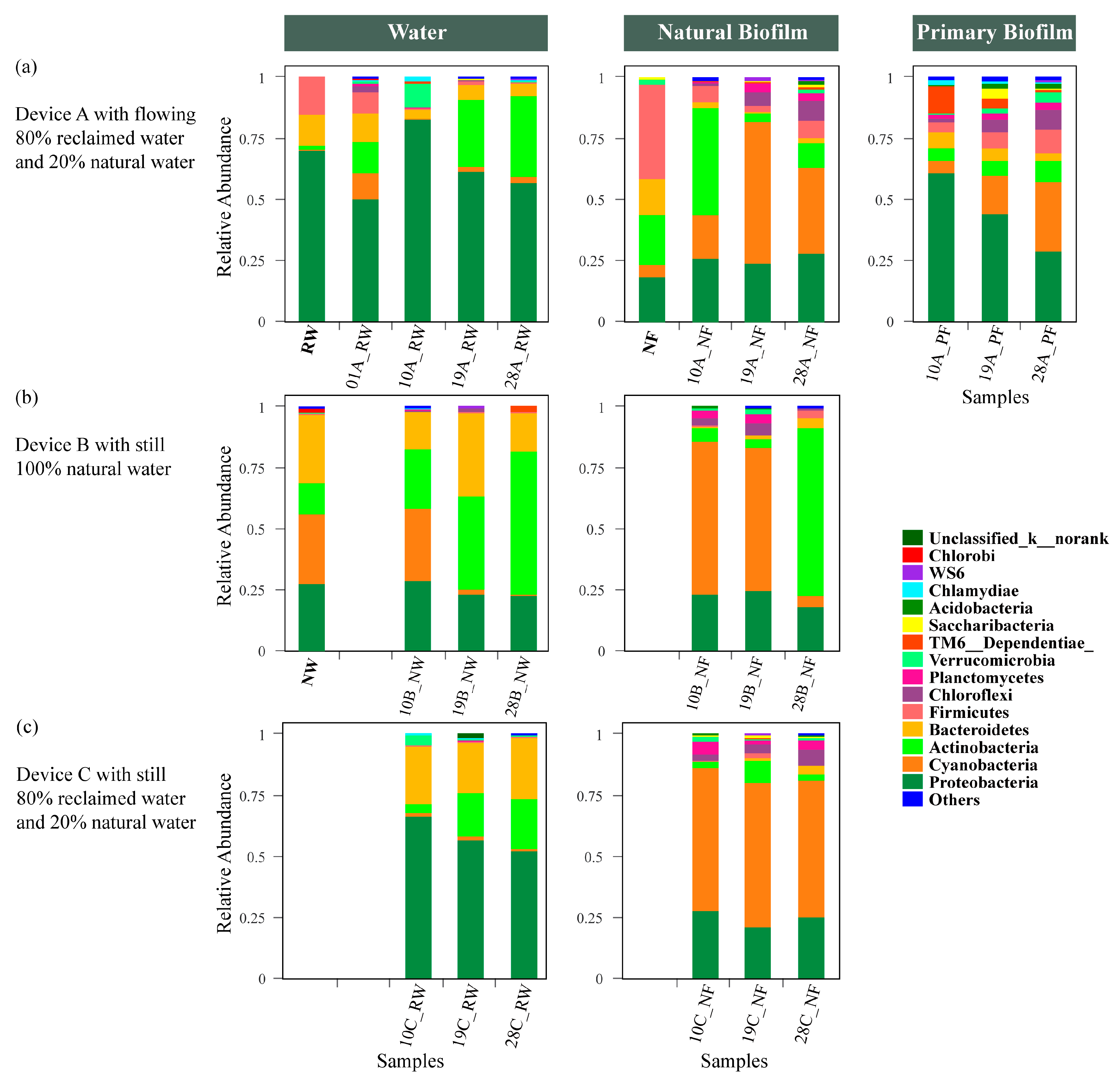
3.4. Bacterial Community Succession in Different Ecological Niches
3.4.1. Water
3.4.2. Natural Biofilm and Primary Biofilm
3.5. Correlation Between Microbial Community and Environmental Factors
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fei, P.P.; Yang, J.P. Analysis of Water Environment Pollution and Pollution Load at Village Scale in Hang-Jia-Hu Water-net Plain. Environ. Sci. Technol. 2011, 34, 104–109. [Google Scholar]
- Tang, W.; Mao, Z.; Zhang, H.; Shan, B.; Zhao, Y.; Ding, Y. Water resources: The prerequisite for ecological restoration of rivers in the Hai River Basin, northern China. Environ. Sci. Pollut. Res. 2015, 22, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, L.; Rene, E.R.; Hu, Q.; Ma, W.; Shen, Z. Biological nitrogen removal using soil columns for the reuse of reclaimed water: Performance and microbial community analysis. J. Environ. Manag. 2018, 217, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Jiao, W.; Chen, X.; Chen, W. An overview of reclaimed water reuse in China. J. Environ. Sci. 2011, 23, 1585–1593. [Google Scholar] [CrossRef]
- Mi, Y.; He, C.; Bian, H.; Cai, Y.; Sheng, L.; Liang, M. Ecological engineering restoration of a non-point source polluted river in Northern China. Ecol. Eng. 2015, 76, 142–150. [Google Scholar] [CrossRef]
- Deng, Y.; Bonilla, M.; Ren, H.; Zhang, Y. Health risk assessment of reclaimed wastewater: A case study of a conventional water reclamation plant in Nanjing, China. Environ. Int. 2018, 112, 235–242. [Google Scholar] [CrossRef]
- Chen, W.; Lu, S.; Jiao, W.; Wang, M.; Chang, A.C. Reclaimed water: A safe irrigation water source? Environ. Dev. 2013, 8, 74–83. [Google Scholar] [CrossRef]
- Pan, W.; Huang, Q.; Huang, G. Nitrogen and Organics Removal during Riverbank Filtration along a Reclaimed Water Restored River in Beijing, China. Water 2018, 10, 491. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Li, Y.; Zhang, Y.; Li, S. Eutrophication of enclosed landscape water supplemented by reclaimed water. J. Tianjin Univ. 2010, 43, 727–732. [Google Scholar]
- Ao, D.; Chen, R.; Wang, X.C.; Liu, Y.; Dzakpasu, M.; Zhang, L.; Huang, Y.; Xue, T.; Wang, N. On the risks from sediment and overlying water by replenishing urban landscape ponds with reclaimed wastewater. Environ. Pollut. 2018, 236, 488–497. [Google Scholar] [CrossRef]
- Longyang, Q.Q. Assessing the effects of climate change on water quality of plateau deep-water lake—A study case of Hongfeng Lake. Sci. Total Environ. 2019, 647, 1518–1530. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, Y.; Xu, T.; Wu, N.; Liang, M.; Hynds, P. Biofilm microbial community structure in an urban lake utilizing reclaimed water. Environ. Earth Sci. 2016, 75, 314. [Google Scholar] [CrossRef]
- Lu, S.; Sun, Y.; Zhao, X.; Wang, L.; Ding, A.; Zhao, X. Sequencing Insights into Microbial Communities in the Water and Sediments of Fenghe River, China. Toxicol. Arch. Environ. Contam. 2016, 71, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Hong, S.; Kim, S.; Zhang, X.; Yang, J.; Giesy, J.P.; Wang, T.; Lu, Y.; Yu, H.; Khim, J.S. Ecogenomic responses of benthic communities under multiple stressors along the marine and adjacent riverine areas of northern Bohai Sea, China. Chemosphere 2016, 172, 166. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.G.; Daley, A.D.; Pechauer, S.M.; Kalscheur, K.N.; Sullivan, M.J.; Kufta, S.L.; Miguel, R.; Gray, K.A.; Kelly, J.J. Development of associations between microalgae and denitrifying bacteria in streams of contrasting anthropogenic influence. FEMS Microbiol. Ecol. 2011, 77, 477–492. [Google Scholar] [CrossRef] [Green Version]
- Smucker, N.J.; Vis, M.L. Acid mine drainage affects the development and function of epilithic biofilms in streams. J. N. Am. Benthol. Soc. 2011, 30, 728–738. [Google Scholar] [CrossRef]
- Wang, T.; Li, Y.; Liang, M.; Yang, P.; Bai, Z. Biofilms on the surface of gravels and aquatic plants in rivers and lakes with reusing reclaimed water. Environ. Earth Sci. 2014, 72, 743–755. [Google Scholar]
- Hobbs, W.O.; Collyard, S.A.; Larson, C.; Carey, A.J.; O’Neill, S.M. Toxic burdens of freshwater biofilms and use as a source tracking tool in rivers and streams. Environ. Sci. Technol. 2019, 53, 11102–11111. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Li, S.; Peng, C.; Cheng, T.; Wang, C.; Guo, L.; Li, D. Nitrogen-cycling microbial community functional potential and enzyme activities in cultured biofilms with response to inorganic nitrogen availability. J. Environ. Sci. 2018, 76, 92–102. [Google Scholar] [CrossRef]
- Chao, Y.; Mao, Y.; Wang, Z.; Zhang, T. Diversity and functions of bacterial community in drinking water biofilms revealed by high-throughput sequencing. Sci. Rep. 2015, 5, 10044. [Google Scholar] [CrossRef] [PubMed]
- Glissman, K.; Chin, K.J.; Casper, P.; Conrad, R. Methanogenic Pathway and Archaeal Community Structure in the Sediment of Eutrophic Lake Dagow: Effect of Temperature. Microb. Ecol. 2004, 48, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Lu, S.; Zhao, X.; Ding, A.; Wang, L. Long-Term Oil Pollution and In Situ Microbial Response of Groundwater in Northwest China. Toxicol. Arch. Environ. Contam. 2017, 72, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Xu, S.; Zheng, D.; Li, J.; Tian, H.; Wang, Y. Effects of haze pollution on microbial community changes and correlation with chemical components in atmospheric particulate matter. Sci. Total Environ. 2018, 637, 507. [Google Scholar] [CrossRef]
- Osman, J.R.; Regeard, C.; Badel, C.; Fernandes, G.; DuBow, M.S. Variation of bacterial biodiversity from saline soils and estuary sediments present near the Mediterranean Sea coast of Camargue (France). Antonie Van Leeuwenhoek 2019, 112, 351–365. [Google Scholar] [CrossRef]
- Ung, P.; Peng, C.; Yuk, S.; Tan, R.; Ann, V.; Miyanaga, K.; Tanji, Y. Dynamics of bacterial community in Tonle Sap Lake, a large tropical flood-pulse system in Southeast Asia. Sci. Total Environ. 2019, 664, 414–423. [Google Scholar] [CrossRef]
- Nan, X.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar]
- Chang, J.; Mei, J.; Jia, W.; Chen, J.; Li, X.; Ji, B.; Wu, H. Treatment of heavily polluted river water by tidal-operated biofilters with organic/inorganic media: Evaluation of performance and bacterial community. Bioresour. Technol. 2019, 279, 34–42. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. Multidiscip. J. Microb. Ecol. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Xing, P. The Vertical Distribution of Sediment Archaeal Community in the “Black Bloom” Disturbing Zhushan Bay of Lake Taihu. Archaea-Int. Microbiol. J. 2016, 2016, 8232135. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Li, J.; Zhao, Y.; Xie, K.; Tang, X.; Wang, S.; Li, Z.; Liao, Y.; Xu, J.; Di, H.; et al. Ammonia oxidizers and nitrite-oxidizing bacteria respond differently to long-term manure application in four paddy soils of south of China. Sci. Total Environ. 2018, 633, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Huang, Y.; Zhang, M.; Yu, Y.; Lu, Y.; Kong, F. Bacterial community dynamics and functional variation during the long-term decomposition of cyanobacterial blooms in vitro. Sci. Total Environ. 2017, 598, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Lai, C.Y.; Shi, L.D.; Wang, K.D.; Dai, Y.J.; Liu, Y.W.; Ma, F.; Rittmann, B.E.; Zheng, P.; Zhao, H.P. Nitrate effects on chromate reduction in a methane-based biofilm. Water Res. 2017, 115, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Dirk, G.; Westcott, S.L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA-based studies. PLoS ONE 2013, 6, e27310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Q.; Yang, H.; Wang, X.; Wang, H. Effects of hexavalent chromium on intestinal histology and microbiota in Bufo gargarizans tadpoles. Chemosphere 2019, 216, 313–323. [Google Scholar] [CrossRef]
- Xue, Y.; Chen, H.; Yang, J.R.; Liu, M.; Huang, B.; Yang, J. Distinct patterns and processes of abundant and rare eukaryotic plankton communities following a reservoir cyanobacterial bloom. ISME J. 2018, 12, 2263–2277. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Kuramae, E.E.; de Hollander, M.; Klinkhamer, P.G.L.; van Veen, J.A. Functional traits dominate the diversity-related selection of bacterial communities in the rhizosphere. ISME J. 2017, 11, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.T.; Zheng, Y.M.; Hu, H.W.; Li, J.; Zhang, L.M.; Chen, B.D.; Chen, W.P.; He, J.Z. Coupling of soil prokaryotic diversity and plant diversity across latitudinal forest ecosystems. Sci. Rep. 2016, 6, 19561. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, Y.; Huang, X.; Ni, P.; Wu, Y.; Deng, Y.; Zhan, A. Adaptive shifts of bacterioplankton communities in response to nitrogen enrichment in a highly polluted river. Environ. Pollut. 2019, 245, 290–299. [Google Scholar] [CrossRef]
- Camargo, J.A.; Alonso, A. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: A global assessment. Environ. Int. 2006, 32, 831–849. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Leveque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Ji, P.; Rhoads, W.J.; Edwards, M.A.; Pruden, A. Impact of water heater temperature setting and water use frequency on the building plumbing microbiome. ISME J. 2017, 11, 1318–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottingham, K.L.; Brown, B.L.; Lennon, J.T. Biodiversity may regulate the temporal variability of ecological systems. Ecol. Lett. 2001, 4, 72–85. [Google Scholar] [CrossRef]
- Matias, M.G.; Combe, M.; Barbera, C.; Mouquet, N. Ecological strategies shape the insurance potential of biodiversity. Front. Microbiol. 2013, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Hua-Fang, S.; Yan, H.; Jin-Ya, W.; Yun-Xia, J.; Nora Fung-Yee, T.; Hong-Wei, Z. Comparison of the levels of bacterial diversity in freshwater, intertidal wetland, and marine sediments by using millions of illumina tags. Appl. Environ. Microbiol. 2012, 78, 8264. [Google Scholar]
- Jiang, X.T.; Peng, X.; Deng, G.H.; Sheng, H.F.; Wang, Y.; Zhou, H.W.; Tam, F.Y. Illumina Sequencing of 16S rRNA Tag Revealed Spatial Variations of Bacterial Communities in a Mangrove Wetland. Microb. Ecol. 2013, 66, 96–104. [Google Scholar] [CrossRef]
- Liu, Q.; Li, M.; Liu, R.; Zhang, Q.; Wu, D.; Zhu, D.; Shen, X.; Feng, C.; Zhang, F.; Liu, X. Removal of trimethoprim and sulfamethoxazole in artificial composite soil treatment systems and diversity of microbial communities. Front. Environ. Sci. Eng. 2019, 13, 28. [Google Scholar] [CrossRef]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A Guide to the Natural History of Freshwater Lake Bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; Liu, C.; Cheng, D.; Ying, X.U.; Liang, Y. Dynamic changes of bacterial community in two eutrophic areas of Donghu Lake in spring and summer. J. Huazhong Agric. Univ. 2017, 36, 52–59. [Google Scholar]
- Sun, W.; Xia, C.; Xu, M.; Guo, J.; Sun, G. Seasonality Affects the Diversity and Composition of Bacterioplankton Communities in Dongjiang River, a Drinking Water Source of Hong Kong. Front. Microbiol. 2017, 8, 1644. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Huang, S.; Sun, G.; Xu, Z.; Xu, M. Phylogenetic diversity, composition and distribution of bacterioplankton community in the Dongjiang River, China. FEMS Microbiol. Ecol. 2012, 80, 30–44. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Stevens, S.L.R.; Chan, L.K.; Bertilsson, S.; Rio, T.G.D.; Tringe, S.G.; Malmstrom, R.R.; Mcmahon, K.D. Ecophysiology of Freshwater Verrucomicrobia Inferred from Metagenome-Assembled Genomes. Msphere 2017, 2, e00277-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Steinberg, C.E.W.; Hartmann, H.M. Planktonic bloom-forming Cyanobacteria and the eutrophication of lakes and rivers. Freshw. Biol. 2010, 20, 279–287. [Google Scholar] [CrossRef]
- Niu, A.; Song, L.Y.; Xiong, Y.H.; Lu, C.J.; Junaid, M.; Pei, D.S. Impact of water quality on the microbial diversity in the surface water along the Three Gorge Reservoir (TGR), China. Ecotoxicol. Environ. Saf. 2019, 181, 412–418. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kouchiwa, T.; Hodoki, Y.; Hotta, K.; Uchida, H.; Harada, K.I. Distribution and identification of actinomycetes lysing cyanobacteria in a eutrophic lake. J. Appl. Phycol. 1998, 10, 391–397. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Suzuki, K. Distribution and algal-lysing activity of fruiting myxobacteria in Lake Suwa. J. Phycol. 2010, 26, 457–462. [Google Scholar] [CrossRef]
- Haukka, K.; Kolmonen, E.; Hyder, R.; Hietala, J.; Vakkilainen, K.; Kairesalo, T.; Haario, H.; Sivonen, K. Effect of Nutrient Loading on Bacterioplankton Community Composition in Lake Mesocosms. Microb. Ecol. 2006, 51, 137–146. [Google Scholar] [CrossRef]
- Falk, W.; Rudolf, A.; Jakob, P. Actinobacterial 16S rRNA genes from freshwater habitats cluster in four distinct lineages. Environ. Microbiol. 2010, 6, 242–253. [Google Scholar]
- Wei, Z.; Huaimin, C.; Ruochen, W.; Ganyu, F.; Zongpu, X.; Siyuan, H.U. Analysis on the reasons for the large bloom area of Lake Taihu in 2017. J. Lake Sci. 2019. [Google Scholar] [CrossRef]
- Mao, Y.F.; Liu, Y.; Li, H.; He, Q.; Ai, H.N.; Gu, W.K.; Yang, G.F. Distinct responses of planktonic and sedimentary bacterial communities to anthropogenic activities: Case study of a tributary of the Three Gorges Reservoir, China. Sci. Total Environ. 2019, 682, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Tian, C. Analysis on Microbial Diversity in the Sediments and Its Relationship with Environmental Factors in a Reservoir. Acta Sci. Nat. Univ. Pekin. 2018, 54, 625–632. [Google Scholar]
- Artigas, J.; Soley, S.; Pérez-Baliero, M.C.; Romaní, A.M.; Ruiz-González, C.; Sabater, S. Phosphorus use by planktonic communities in a large regulated Mediterranean river. Sci. Total Environ. 2012, 426, 180–187. [Google Scholar] [CrossRef]
- Huang, S.; Voutchkov, N.; Jiang, S. Balancing carbon, nitrogen and phosphorus concentration in seawater as a strategy to prevent accelerated membrane biofouling. Water Res. 2019, 165, 114978. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | B | C | ||
|---|---|---|---|---|
| Temperature (°C) | max | 29.1 | 28.0 | 27.9 |
| min | 28.1 | 27.2 | 27.2 | |
| pH | max | 9.4 | 9.9 | 9.8 |
| min | 9.1 | 9.6 | 9.6 | |
| Chemical oxygen demand (COD; mg/L) | max | 31 | 49 | 25 |
| min | 20 | 20 | 15 | |
| Dissolved oxygen (DO; mg/L) | max | 9.40 | 11.29 | 12.63 |
| min | 6.78 | 8.92 | 9.42 | |
| Nitrate (mg/L) | max | 190.60 | 2.06 | 51.98 |
| min | 43.56 | 0.51 | 42.66 | |
| Phosphate (mg/L) | max | 0.51 | 0 | 0.54 |
| min | 0 | 0 | 0.27 | |
| Ammonia nitrogen (mg/L) | max | 0.24 | 0.18 | 0.24 |
| min | 0.13 | 0 | 0.02 | |
| Conductivity (S/m) | max | 0.166 | 0.118 | 0.129 |
| min | 0.091 | 0.091 | 0.091 | |
| Na+ (mg/L) | max | 168.19 | 173.99 | 183.51 |
| min | 98.67 | 116.90 | 128.98 | |
| K+ (mg/L) | max | 19.53 | 10.96 | 21.62 |
| min | 12.83 | 7.19 | 15.36 | |
| Mg2+ (mg/L) | max | 49.35 | 41.77 | 33.97 |
| min | 28.30 | 34.70 | 31.22 | |
| Ca2+ (mg/L) | max | 118.11 | 27.90 | 46.22 |
| min | 65.35 | 24.34 | 32.55 | |
| Cl− (mg/L) | max | 282.80 | 189.78 | 240.87 |
| min | 131.82 | 123.07 | 163.90 | |
| SO42− (mg/L) | max | 292.04 | 231.11 | 201.39 |
| min | 119.46 | 160.95 | 144.67 |
| Sample | Operational Taxonomic Units (OTUs) | Shannon | Simpson | Chao |
|---|---|---|---|---|
| Natural water (NW) | 373 | 3.45 | 0.07 | 691.82 |
| Reclaimed water (RW) | 279 | 2.52 | 0.14 | 453.15 |
| 01A_RW | 731 | 4.24 | 0.04 | 926.71 |
| 10A_RW | 324 | 2.68 | 0.17 | 511.64 |
| 19A_RW | 473 | 2.62 | 0.16 | 705.60 |
| 28A_RW | 419 | 2.76 | 0.14 | 687.83 |
| 10B_NW | 492 | 3.78 | 0.05 | 819.54 |
| 19B_NW | 372 | 3.19 | 0.08 | 649.22 |
| 28B_NW | 337 | 2.56 | 0.25 | 603.22 |
| 10C_RW | 442 | 3.16 | 0.08 | 679.78 |
| 19C_RW | 422 | 3.31 | 0.07 | 714.97 |
| 28C_RW | 411 | 3.17 | 0.08 | 660.35 |
| Natural biofilm (NF) | 405 | 3.74 | 0.05 | 508.26 |
| 10A_NF | 530 | 3.48 | 0.16 | 591.33 |
| 19A_NF | 835 | 4.53 | 0.04 | 1100.04 |
| 28A_NF | 1236 | 5.23 | 0.02 | 1739.22 |
| 10A_PF | 804 | 4.30 | 0.04 | 1205.37 |
| 19A_PF | 1126 | 5.16 | 0.02 | 1483.47 |
| 28A_PF | 1272 | 5.25 | 0.02 | 1595.97 |
| 10B_NF | 673 | 4.60 | 0.02 | 898.18 |
| 19B_NF | 737 | 4.57 | 0.04 | 952.36 |
| 28B_NF | 204 | 2.14 | 0.44 | 206.63 |
| 10C_NF | 545 | 4.26 | 0.03 | 718.04 |
| 19C_NF | 751 | 4.88 | 0.02 | 846.21 |
| 28C_NF | 684 | 4.59 | 0.04 | 815.68 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Sun, Y.; Wang, X.; Xu, S. Changes in Microbial Community Structures under Reclaimed Water Replenishment Conditions. Int. J. Environ. Res. Public Health 2020, 17, 1174. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17041174
Li J, Sun Y, Wang X, Xu S. Changes in Microbial Community Structures under Reclaimed Water Replenishment Conditions. International Journal of Environmental Research and Public Health. 2020; 17(4):1174. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17041174
Chicago/Turabian StyleLi, Jie, Yujiao Sun, Xiaoyu Wang, and Shangwei Xu. 2020. "Changes in Microbial Community Structures under Reclaimed Water Replenishment Conditions" International Journal of Environmental Research and Public Health 17, no. 4: 1174. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17041174