Seed Biopriming with Microbial Inoculant Triggers Local and Systemic Defense Responses against Rhizoctonia solani Causing Banded Leaf and Sheath Blight in Maize (Zea mays L.)

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of Pseudomonas aeruginosa MF-30
2.2. Green Fluorescent Protein (GFP)-Tagging and Root Colonization Assay
2.3. Nethouse Experiments
2.3.1. Preparation of Liquid Formulation
2.3.2. Planting Material and Growth Conditions
2.3.3. Experimental Set-Up
2.3.4. Effects of Seed Bio-Priming on the Accumulation of Defense-Related Biomolecules and Enzymes
2.3.5. RNA Extraction and qRT–PCR
2.3.6. Effects of Seed Bio-Priming on Disease Severity and Plant Growth
2.4. Statistical Analyses
3. Results
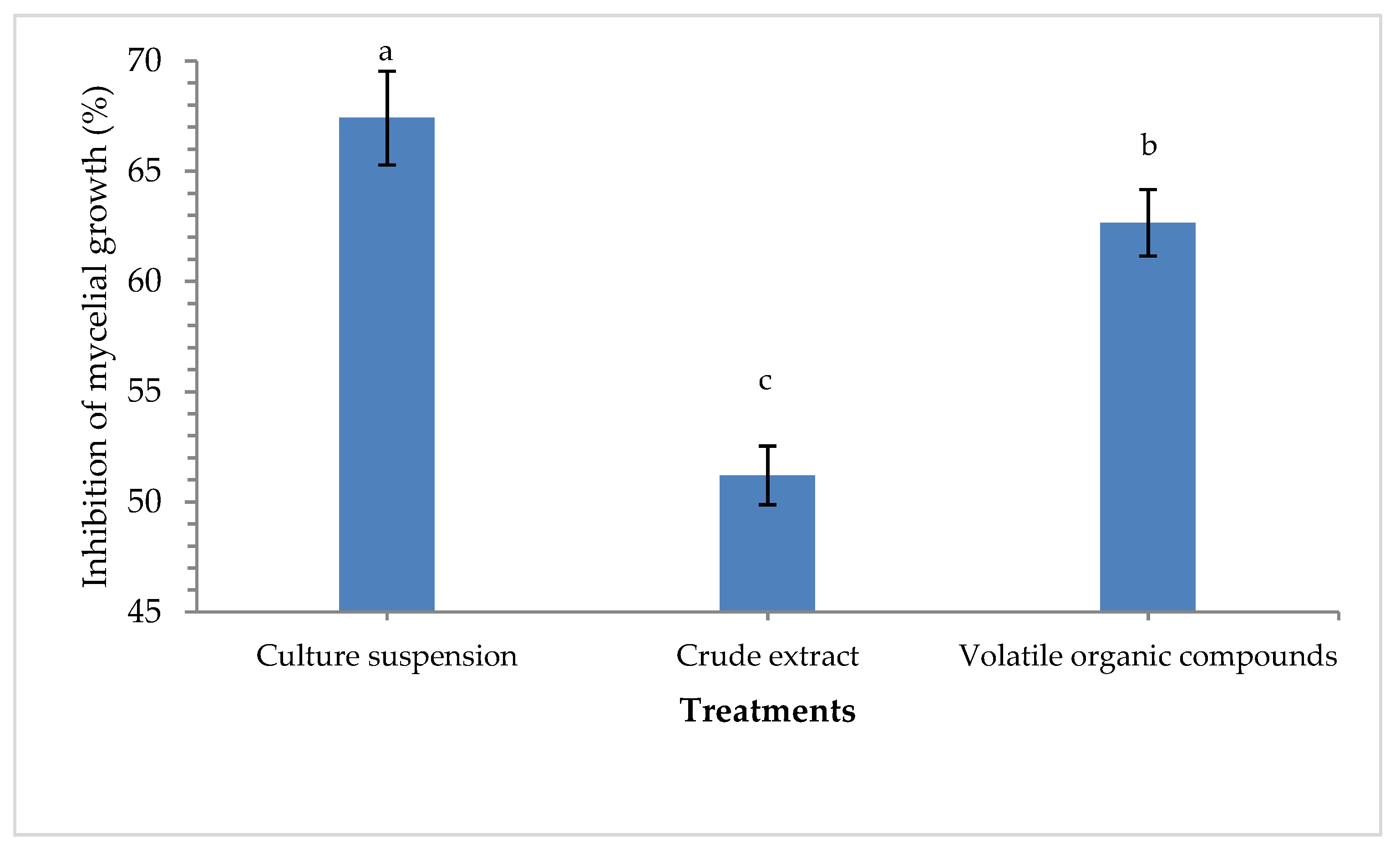
3.1. Characterization of Pseudomonas aeruginosa MF-30
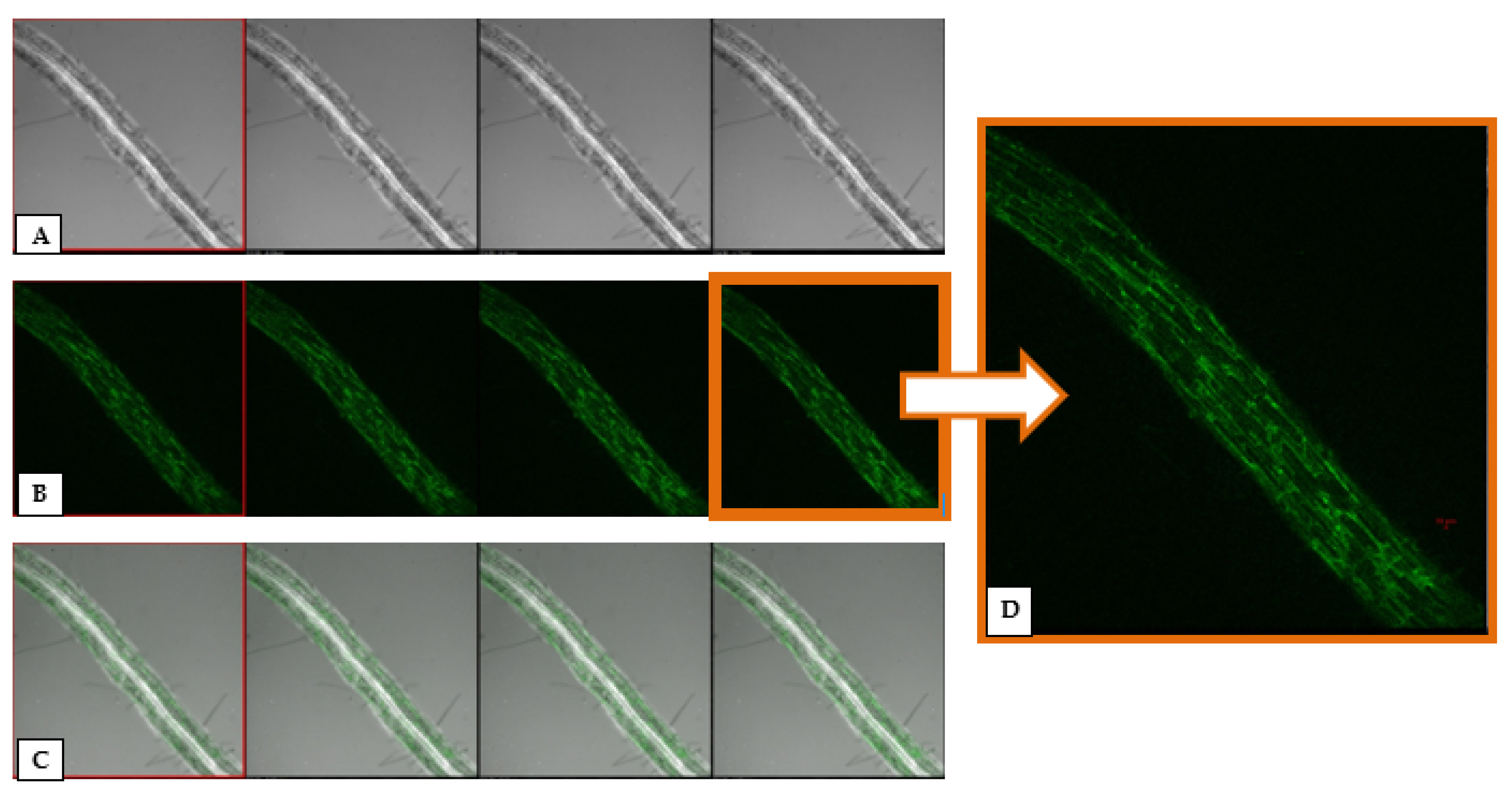
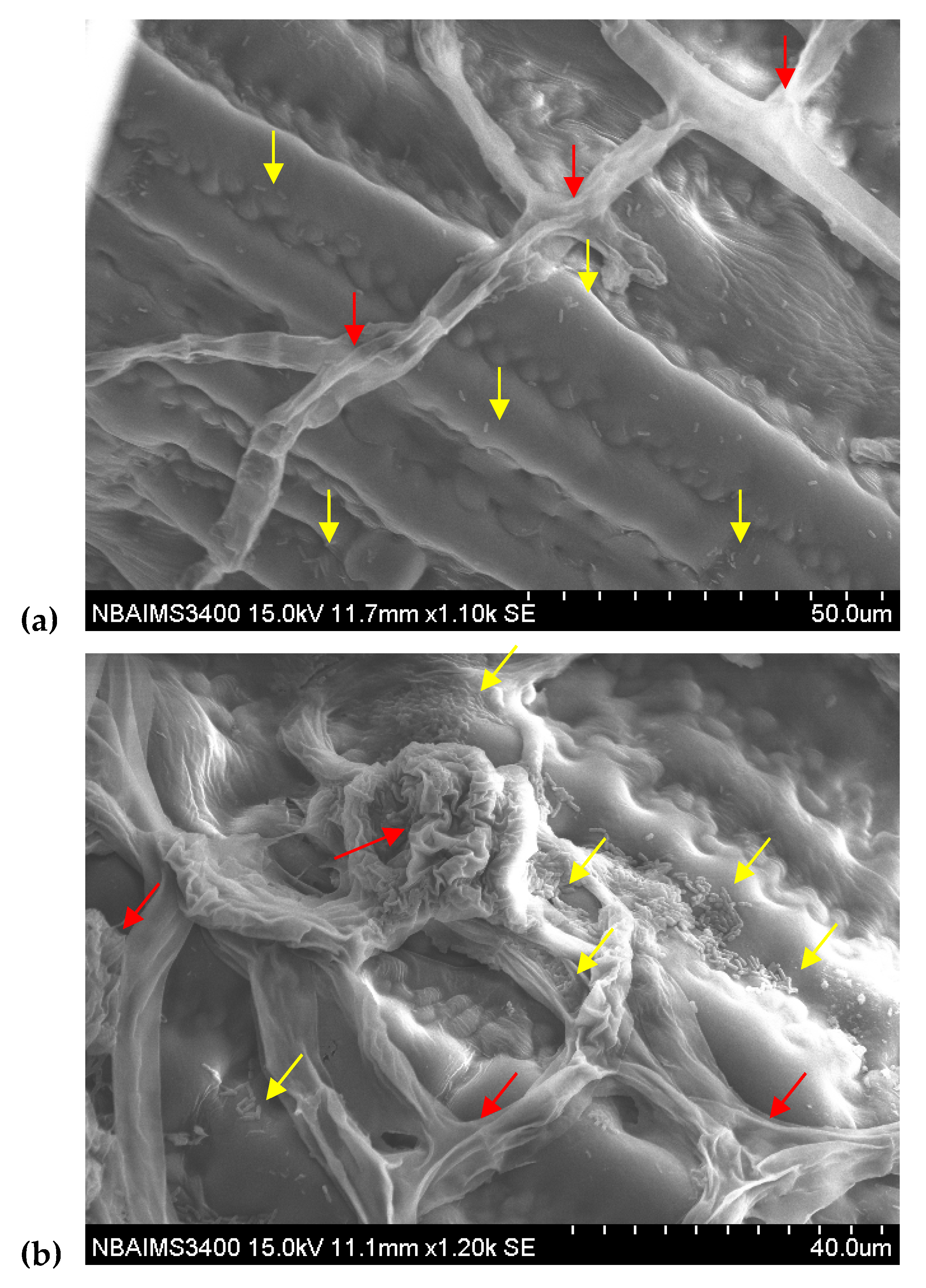
3.2. GFP-Tagging and Root Colonization Assay
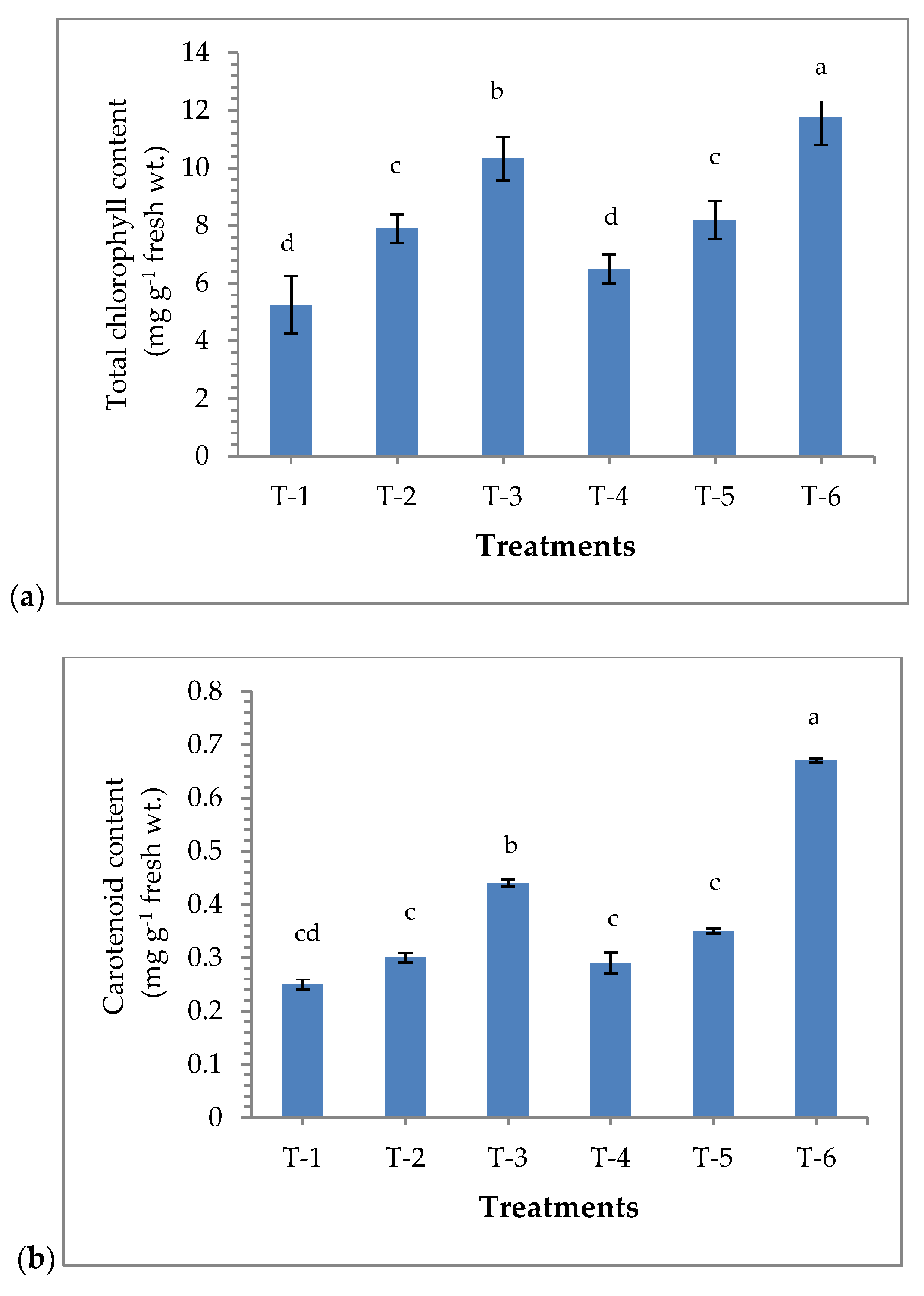
3.3. Effects of Seed Bio-Priming on the Accumulation of Chlorophyll and Carotenoids Content
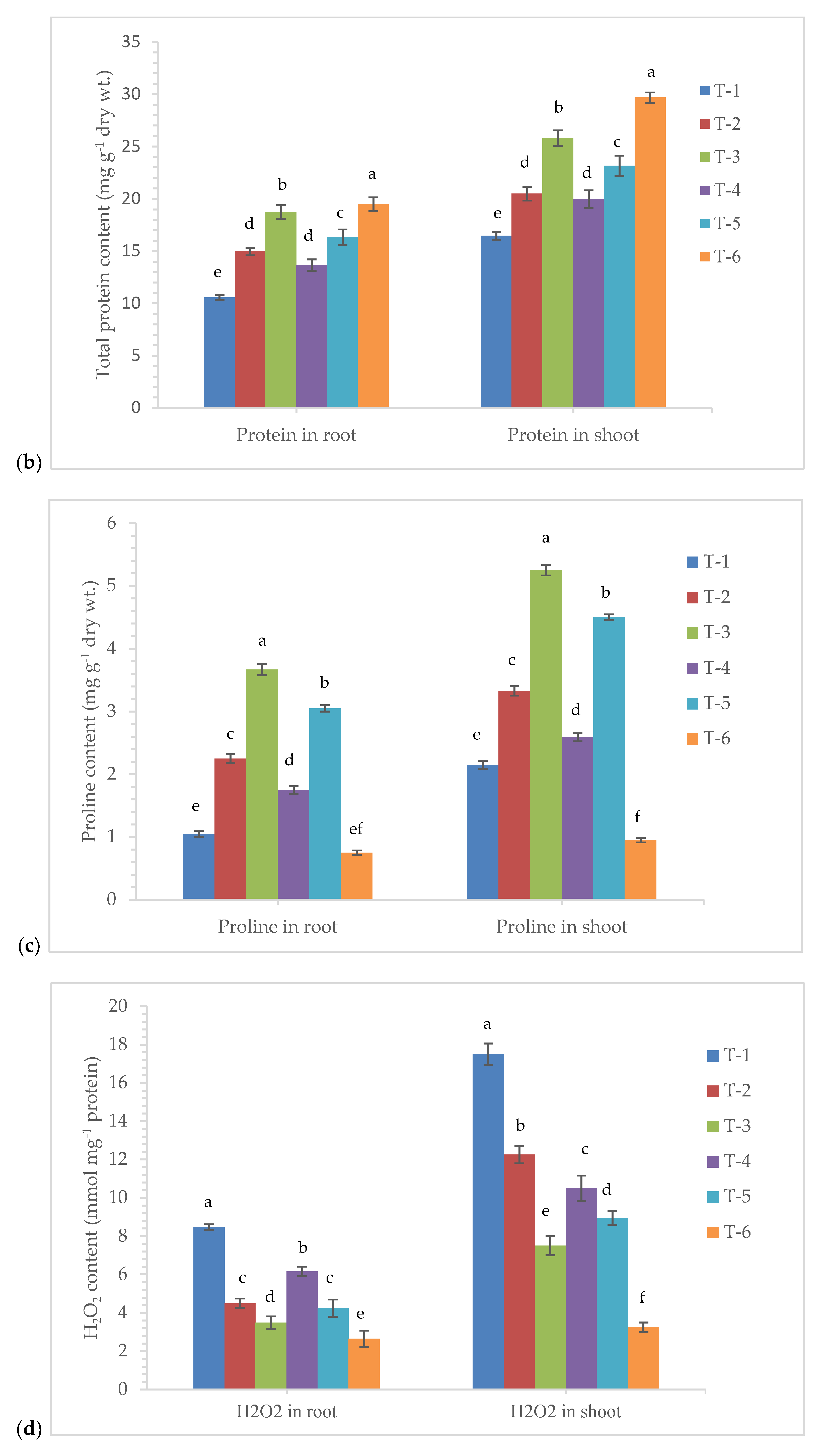
3.4. Effects of Seed Bio-Priming on Total Soluble Sugar, Protein, Proline, and H2O2 Content
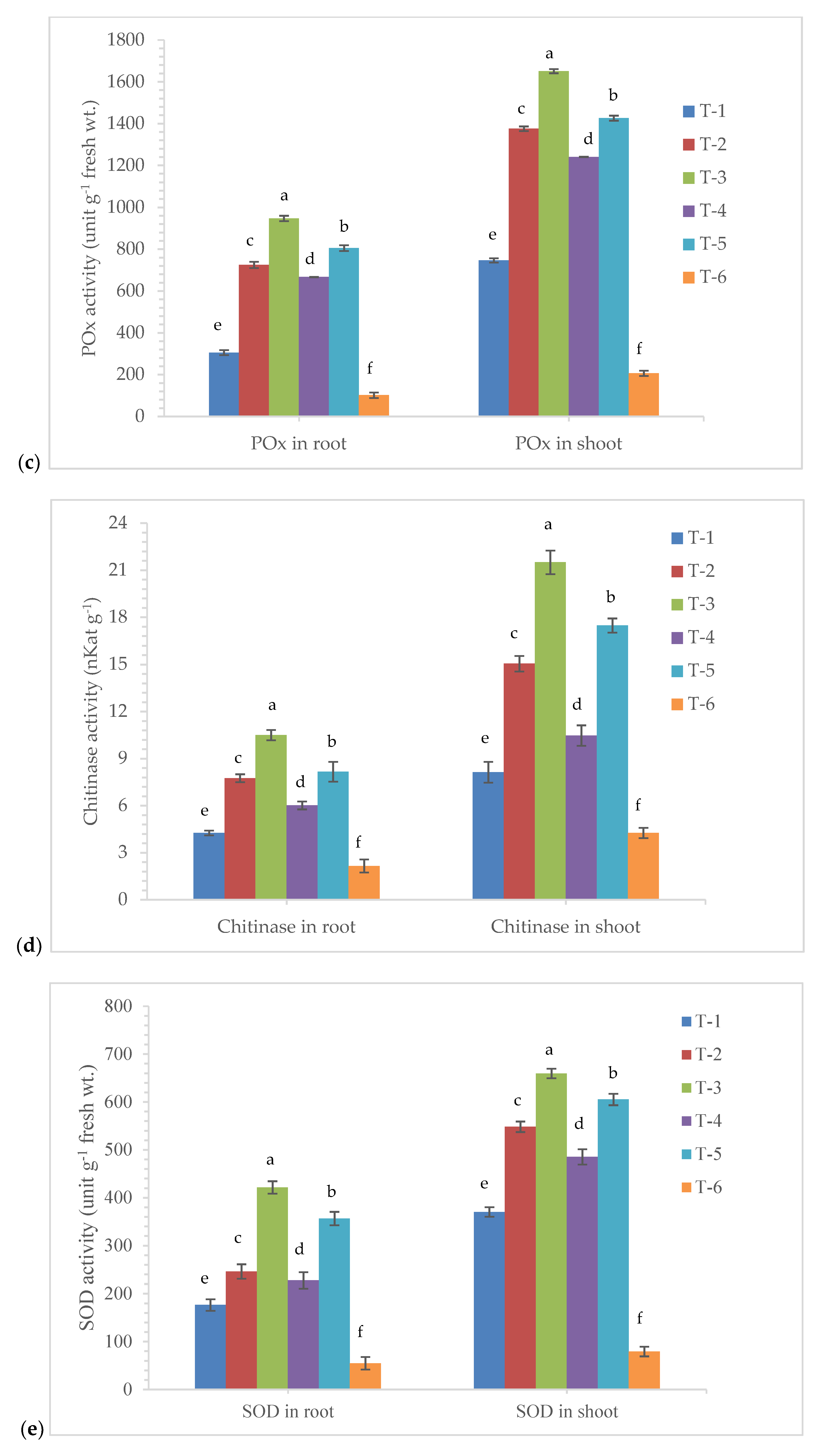
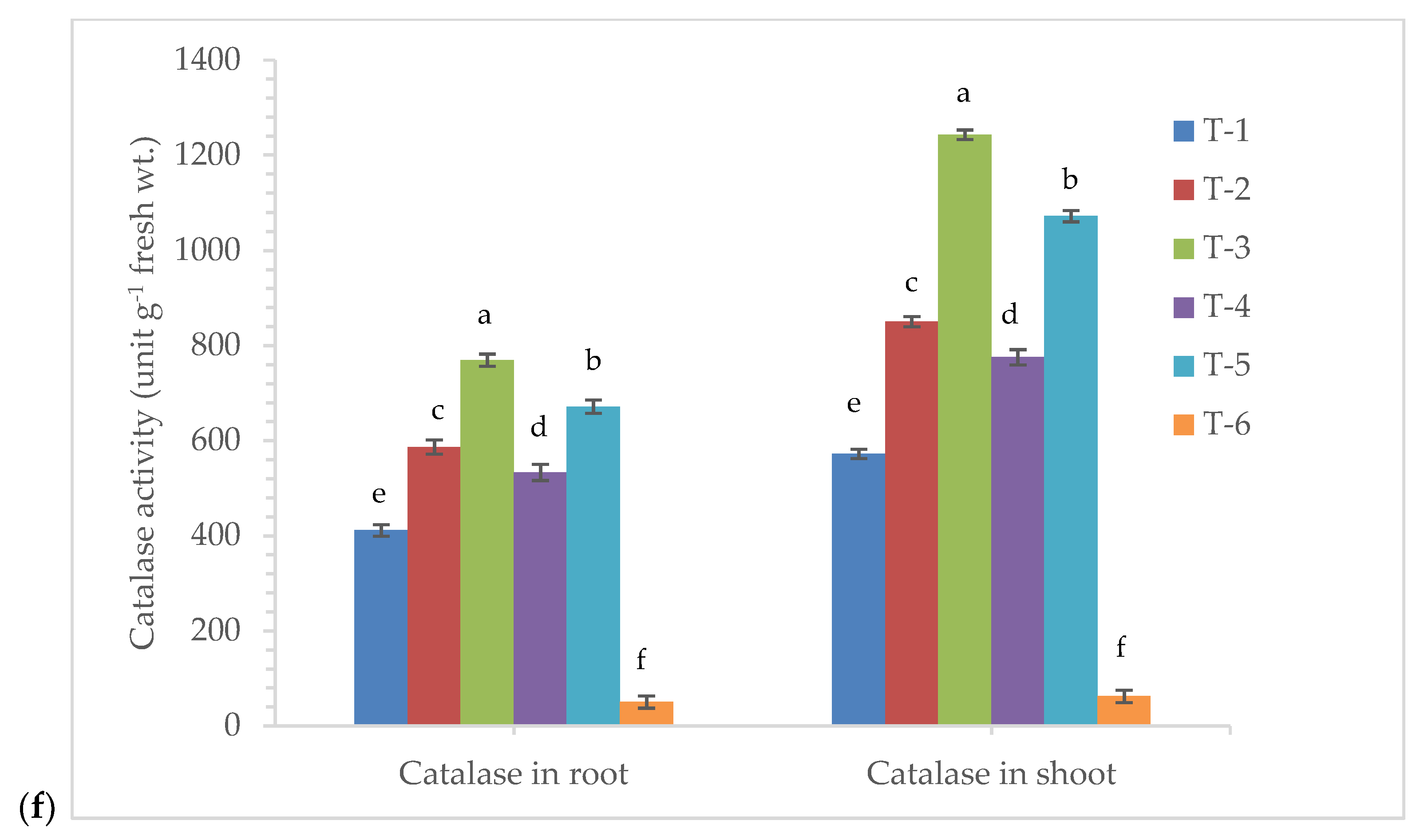
3.5. Effects of Seed Bio-Priming on Antioxidant Enzymes Activity
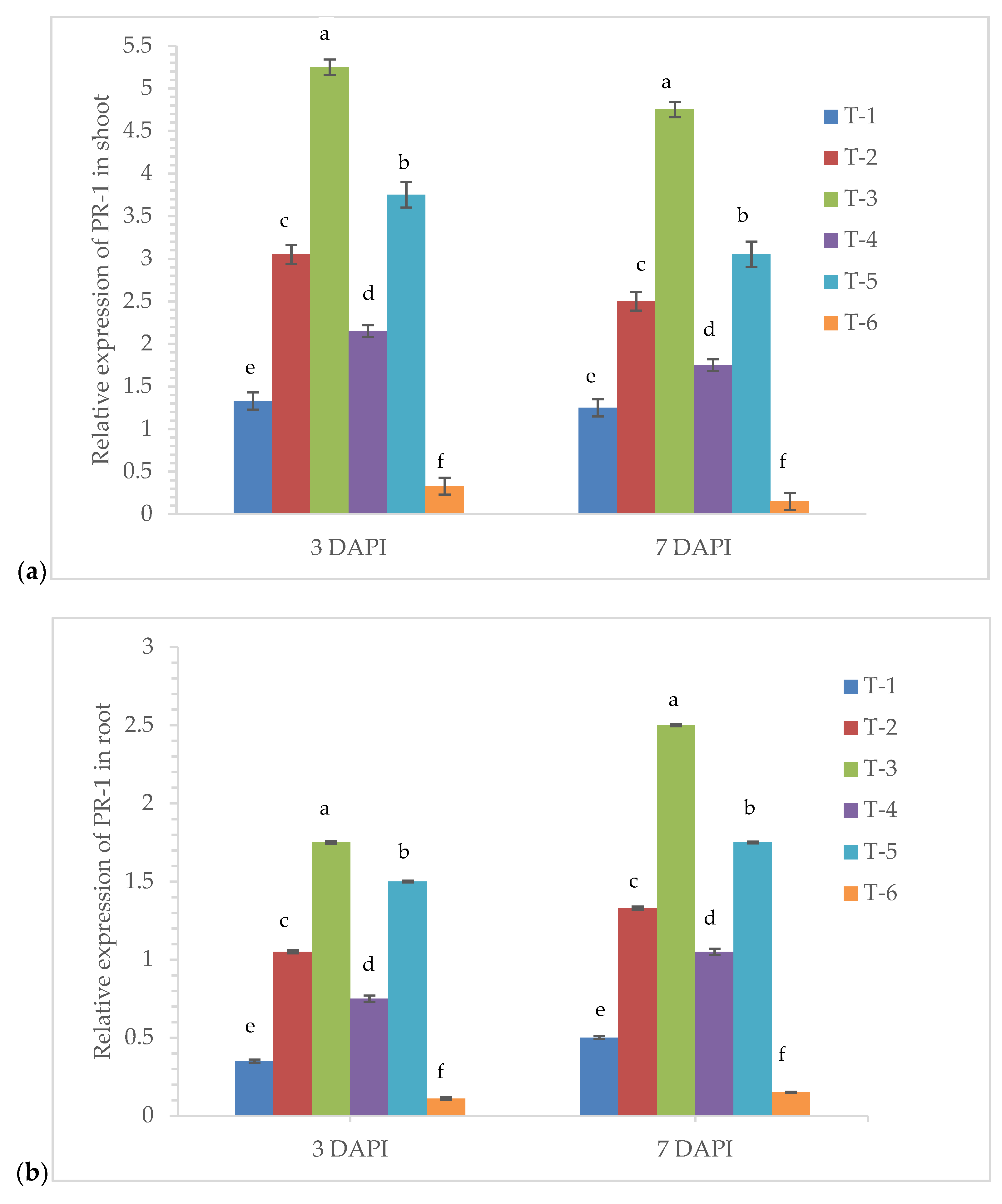
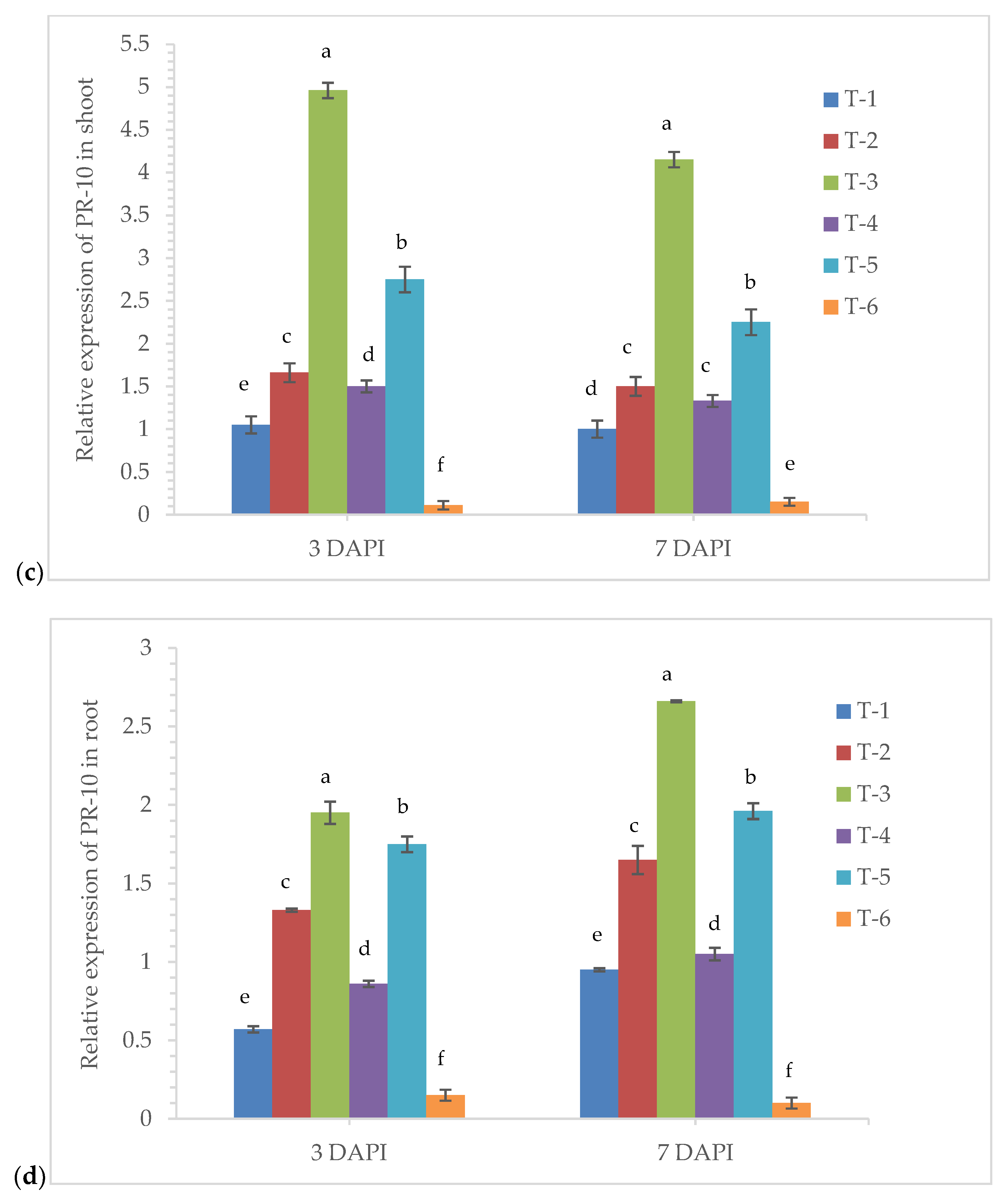
3.6. Gene Expression Analysis
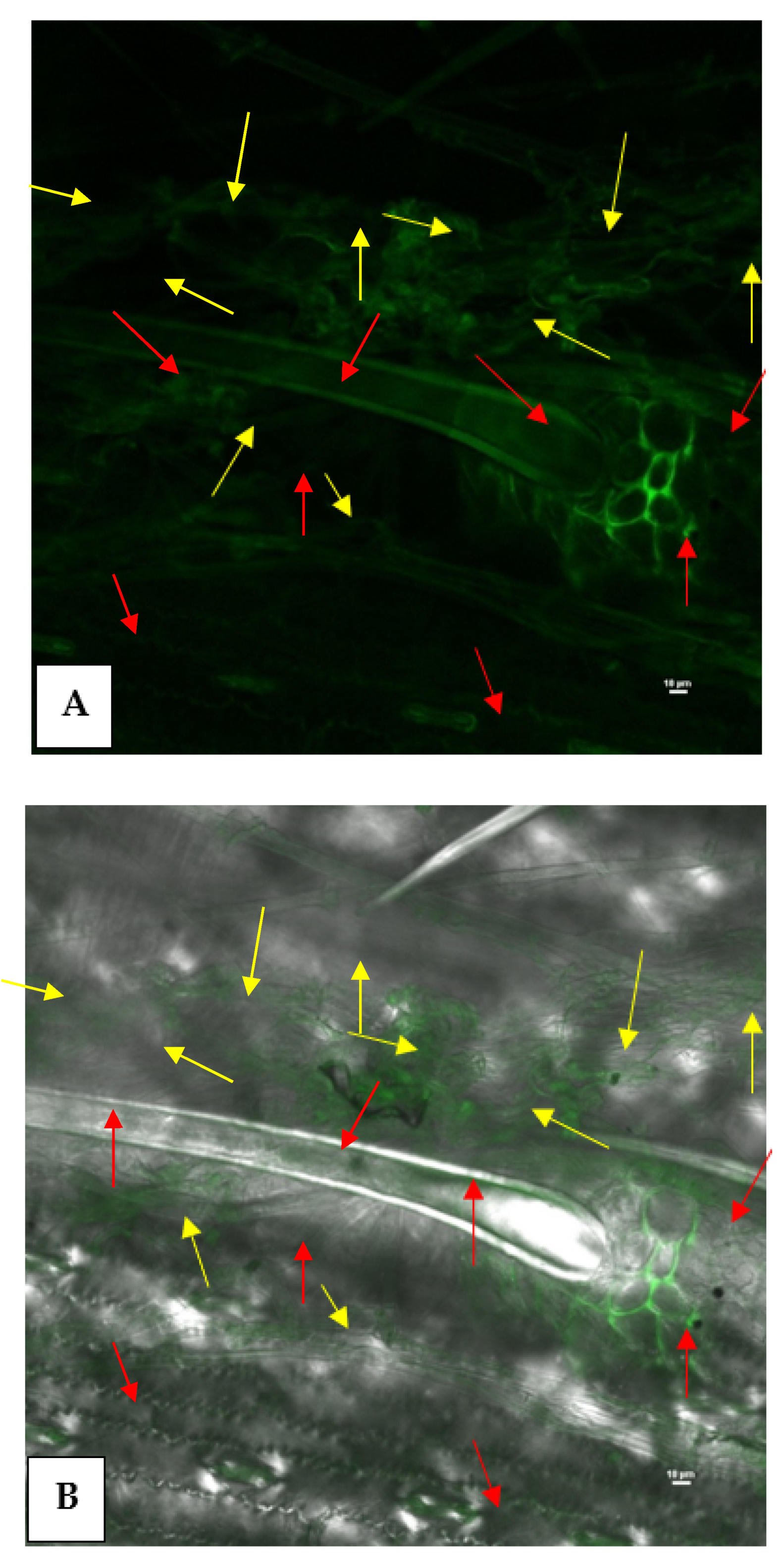
3.7. Epiphytic Colonization of GFP-Tagged P. aeruginosa MF-30 on Maize Leaf
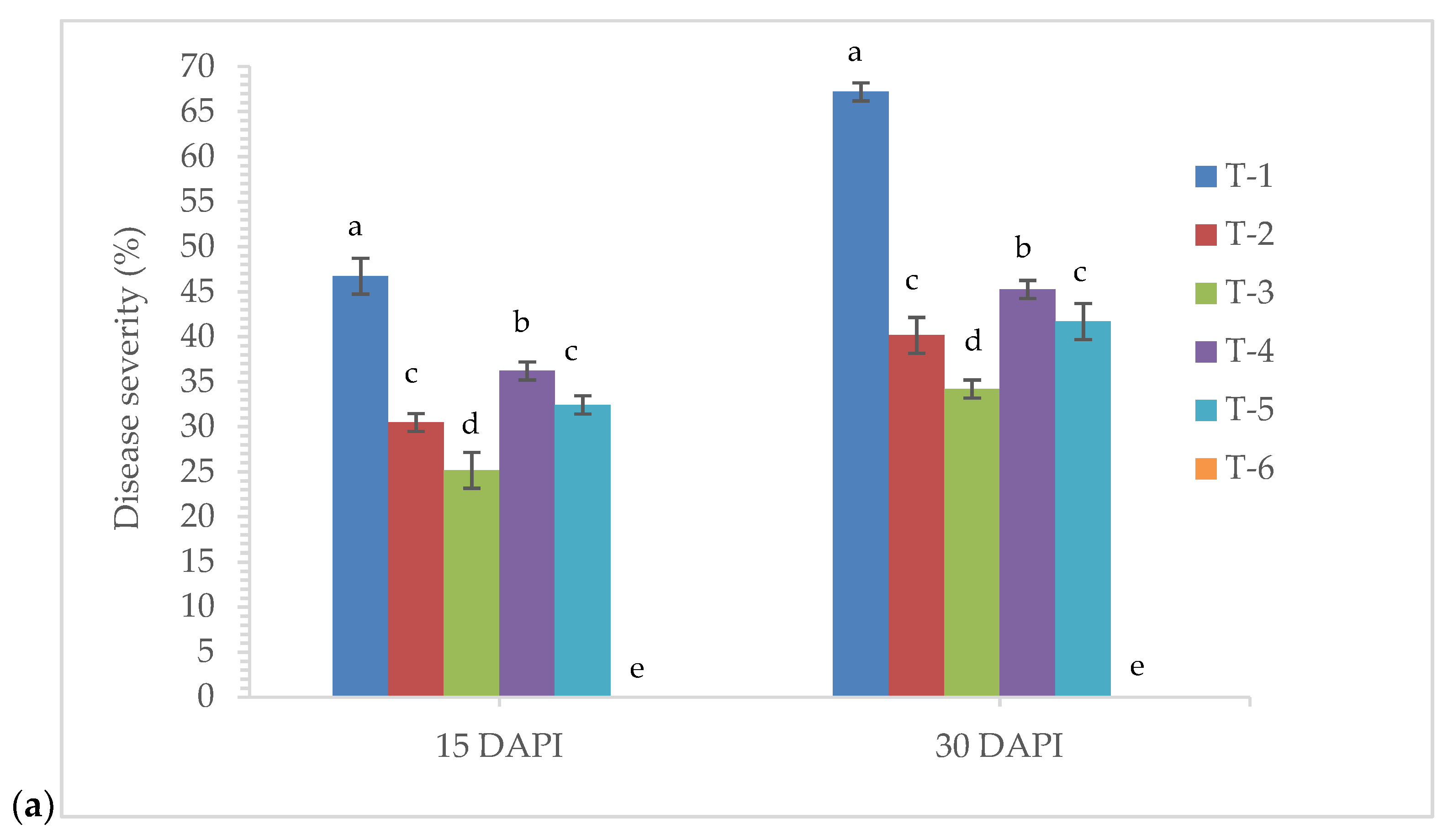
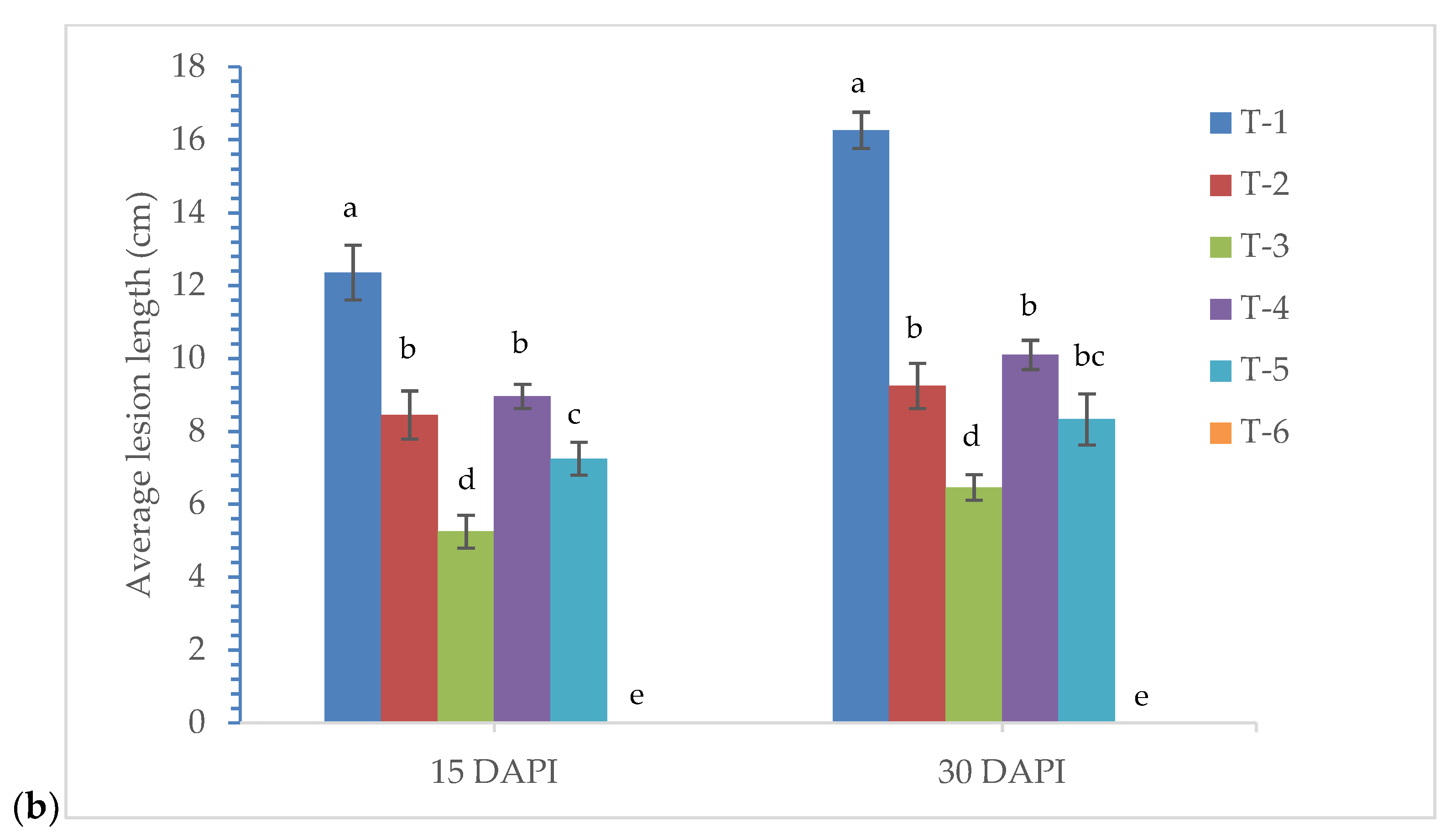
3.8. Effects of Seed Bio-Priming on Plant Growth and Disease Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anonymous. Annual Progress Report (AICRP on Maize): Kharif Maize; Indian Institute of Maize Research: Ludhiyana, India, 2016; p. 318.
- Anonymous. Annual Progress Report (AICRP on Maize): Rabi Maize; Indian Institute of Maize Research: Ludhiyana, India, 2017; p. 992.
- Payak, M.M.; Renfro, B.L. Diseases of maize new to India. Indian Phytopathol. Soc. Bull. 1966, 3, 14–18. [Google Scholar]
- Thakur, S.M.; Sharma, S.L.; Munjal, R.L. Correlation studies between incidence of banded sclerotial disease and ear yield in maize. Indian J. Mycol. Plant Pathol. 1973, 3, 180–181. [Google Scholar]
- Sumner, D.R.; Minton, N.A. Crop losses in corn induced by Rhizoctonia solani AG2-2 and nematodes. Phytopathology 1989, 79, 934–941. [Google Scholar] [CrossRef]
- Li, N.; Lin, B.; Wang, H.; Li, X.; Yang, F.; Ding, X.; Yan, J.; Chu, Z. Natural variation in ZmFBL41 confers banded leaf and sheath blight resistance in maize. Nat. Genet. 2019, 51, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, B.M.; Pixley, K.; Warburton, M.L.; Xie, C.-X. Molecular marker-assisted breeding options for maize improvement in Asia. Mol. Breed. 2010, 26, 339–356. [Google Scholar] [CrossRef]
- Chen, G.P.; Tan, H.; Zheng, D.B.; Yang, L.T.; Li, Y.R. QTL mapping of resistance to banded leaf and sheath blight in maize. Southwest China J. Agric. Sci. 2009, 22, 950–955. [Google Scholar]
- Lin, H.; Leng, P.; Pan, G.; Zhang, Z. Association analysis of candidate quantitative trait loci for resistance to banded leaf and sheath blight in maize. Int. J. Biosci. Biochem. Bioinform. 2013, 3, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Shahi, J.P. Banded leaf and sheath blight: An emerging disease of maize. Maydica 2012, 57, 215–219. [Google Scholar]
- Tang, H.T.; Rong, T.Z.; Yang, J.P. Research advance on sheath blight (Zea mays L.) in maize. J. Maize Sci. 2004, 12, 93–96. [Google Scholar]
- Sharma, R.C.; Srinivas, P.; Batsa, B.K. Banded leaf and sheath blight of maize its epidemiology and management. In Proceedings of a Maize Symposium; Rajbhandari, N.P., Ransom, J.K., Adhikari, K., Palmer, A.F.E., Eds.; NARC and CIMMYT: Kathmandu, Nepal, 2002; pp. 108–112. [Google Scholar]
- Singh, B.M.; Sharma, Y.R. Evaluation of maize germplasm to banded sclerotial disease and assessment of yield loss. Indian Phytopathol. 1976, 29, 129–132. [Google Scholar]
- Pinson, S.R.M.; Capdevielle, F.M.; Oard, J.H. Confirming QTLs and finding additional loci conditioning sheath blight resistance in rice using recombinant inbred lines. Crop Sci. 2005, 45, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.R.; Gour, H.N.; Rathore, R.S. Identification of host resistance against banded leaf and sheath blight of maize. J. Mycol. Plant Pathol. 2003, 33, 313–314. [Google Scholar]
- Sharma, R.C.; Rai, S.N.; Batsa, B.K. Identifying resistance to banded leaf and sheath blight of maize. Indian Phytopathol. 2005, 58, 121–122. [Google Scholar]
- Anshu, G.; Prasanna, B.M.; Sharma, R.C.; Rathore, R.S.; Saxena, S.C. Identification of resistance sources to banded leaf and sheath blight (Rhizoctonia solanif. sp. sasakii) in maize. Indian Phytopathol. 2007, 60, 162–166. [Google Scholar]
- Hooda, K.S.; Karjagi, C.G.; Sekhar, J.C.; Kumar, R.; Sai, D.S.; Kumar, S.; Shekhar, M.; Gowda, K.T.; Sharma, S.S.; Kaur, H.; et al. Identification of stable sources of disease resistance and their use in the development of single cross hybrids in maize. In Proceedings of the 11th Asian Maize Conference on “Addressing Climate Change Effects and Meeting Maize Demand for Asia”, Nanning, China, 7–11 November 2011; pp. 104–105. [Google Scholar]
- Madhavi, M.; Narayan, P.R.; Ranga, R.; Sudarshan, M.R. Evaluation of maize genotypes against banded leaf and sheath blight disease incited by Rhizoctonia solani f. sp. sasakii (Kuhn) Exner. J. Res. ANGRAU 2012, 40, 20–23. [Google Scholar]
- Vimla, B.; Mukherjee, B.K.; Ahuja, V.P. Combining ability analysis for resistance to banded leaf and sheath blight of maize. Indian J. Genet. 1988, 48, 75–79. [Google Scholar]
- Zhao, M.J.; Zhang, Z.M.; Zhang, S.H.; Li, W.; Jeffers, D.P.; Rong, T.Z.; Pan, G.T. Quantitative trait loci for resistance to banded leaf and sheath blight in maize. Crop Sci. 2006, 46, 1039–1045. [Google Scholar] [CrossRef]
- Yobo, K.S.; Laing, M.D.; Hunter, C.H.; Morris, M.J. Biological control of Rhizoctonia solani by two Trichoderma species isolated from South African composted soil. S. Afr. J. Plant Soil 2004, 88, 265–268. [Google Scholar] [CrossRef]
- Sivakumar, G.; Sharma, R.C.; Rai, S.N. Biocontrol of banded leaf and sheath blight of peat based Pseudomonas fluorescens formulation. Indian Phytopathol. 2000, 53, 190–192. [Google Scholar]
- Sharma, G.; Saxena, S.C. Evaluation of biocontrol agents against Rhizoctonia solani leaf and sheath blight of maize. Ann. Plant Prot. Soc. 2001, 9, 144–145. [Google Scholar]
- Meena, R.; Rathore, L.; Mathur, R.S. Efficacy of biocontrol agents against Rhizoctonia solani f. sp. sasakii causing banded leaf and sheath blight of maize. J. Mycol. Plant Pathol. 2003, 33, 310–312. [Google Scholar]
- Kuc, J. Induced immunity to plant disease. BioScience 1982, 32, 854–860. [Google Scholar]
- Bruisson, S.; Zufferey, M.; L’Haridon, F.; Trutmann, E.; Anand, A.; Dutartre, A.; De Vrieze, M.; Weisskopf, L. Endophytes and Epiphytes From the Grapevine Leaf Microbiome as Potential Biocontrol Agents Against Phytopathogens. Front. Microbiol. 2019, 10, 2726. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Audenaert, K.; Pattery, T.; Cornelis, P.; Höfte, M. Induction of systemic resistance to Botrytis cinerea in tomato by Pseudomonas aeruginosa 7NSK2: Role of salicylic acid, pyochelin, and pyocyanin. Mol. Plant Microbe Interact. 2002, 15, 1147–1156. [Google Scholar] [CrossRef] [Green Version]
- Kishore, G.K.; Pande, S.; Rao, J.N.; Podile, A.R. Pseudomonas aeruginosa inhibits the plant cell wall degrading enzymes of Sclerotium rolfsiiand reduces the severity of groundnut stem rot. Eur. J. Plant Pathol. 2005, 113, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.S.; Ayyadurai, N.; Pandiaraja, P.; Reddy, A.V.; Venkateswarlu, Y.; Prakash, O.; Sakthivel, N. Characterization of antifungal metabolite produced by a new strain Pseudomonas aeruginosa PUPa3 that exhibits broad-spectrum antifungal activity and biofertilizing traits. J. Appl. Microbiol. 2005, 98, 145–154. [Google Scholar] [CrossRef]
- Sahu, P.K.; Singh, S.; Gupta, A.; Singh, U.B.; Brahmaprakash, G.P.; Saxena, A.K. Antagonistic potential of bacterial endophytes and induction of systemic resistance against collar rot pathogen Sclerotium rolfsii in tomato. Biol. Control 2019, 137, 104014. [Google Scholar] [CrossRef]
- Krzyzanowska, D.; Obuchowski, M.; Bikowski, M.; Rychlowski, M.; Jafra, S. Colonization of potato rhizosphere by GFP-tagged Bacillus subtilis MB73/2, Pseudomonas sp. P482 and Ochrobactrum sp. A44 shown on large sections of roots using enrichment sample preparation and confocal laser scanning microscopy. Sensors 2012, 12, 17608–17619. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, W.; Kinkema, M.; Li, X.; Dong, X. Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. Proc. Natl. Acad. Sci. USA 1999, 96, 6523–6528. [Google Scholar] [CrossRef] [Green Version]
- Mou, Z.; Fan, W.; Dong, X. Inducers of Plant Systemic Acquired Resistance Regulate NPR1 Function through Redox Changes. Cell 2003, 113, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Durrent, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.B.; Malviya, D.; Singh, S.; Pradhan, J.K.; Singh, B.P.; Roy, M.; Imram, M.; Pathak, N.; Baisyal, B.M.; Rai, J.P.; et al. Bio-protective microbial agents from rhizosphere eco-systems trigger plant defense responses provide protection against sheath blight disease in rice (Oryza sativa L.). Microbiol. Res. 2016, 192, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.B.; Malviya, D.; Singh, S.; Imran, M.; Pathak, N.; Alam, M.; Rai, J.P.; Singh, R.K.; Sarma, B.K.; Sharma, P.K.; et al. Compatible salt-tolerant rhizosphere microbe-mediated induction of phenylpropanoid cascade and induced systemic responses against Bipolaris sorokiniana (Sacc.) Shoemaker causing spot blotch disease in wheat (Triticum aestivum L.). Appl. Soil Ecol. 2016, 108, 300–306. [Google Scholar] [CrossRef]
- Fan, W.; Dong, X. In Vivo Interaction between NPR1 and Transcription Factor TGA2 Leads to Salicylic Acid–Mediated Gene Activation in Arabidopsis. Plant Cell 2002, 14, 1377–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molla, K.A.; Karmakar, S.; Chanda, P.K.; Sarkar, S.N.; Datta, S.K.; Datta, K. Tissue-specific expression of Arabidopsis NPR1 gene in rice for sheath blight resistance without compromising phenotypic cost. Plant Sci. 2016, 250, 105–114. [Google Scholar] [CrossRef]
- Sarma, B.K.; Yadav, S.K.; Singh, S.; Singh, H.B. Microbial consortium mediated plant defense against phytopathogens: Readdressing for enhancing efficacy. Soil Biol. Biochem. 2015, 87, 25–33. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Singh, S.; Kumar, M.; Sahu, P.K.; Singh, H.V.; Kumar, S.; Roy, M.; Imran, M.; Rai, J.P.; et al. Trichoderma harzianum- and Methyl Jasmonate-Induced Resistance to Bipolaris sorokiniana Through Enhanced Phenylpropanoid Activities in Bread Wheat (Triticum aestivum L.). Front. Microbiol. 2019, 10, 1697. [Google Scholar] [CrossRef] [Green Version]
- Devi, K.A.; Pandey, G.; Rawat, A.K.S.; Sharma, G.D.; Pandey, P. The Endophytic Symbiont—Pseudomonas aeruginosa Stimulates the Antioxidant Activity and Growth of Achyranthes aspera L. Front. Microbiol. 2017, 8, 1897. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Xu, J.; Xie, W.; Yao, Z.; Yang, H.; Sun, C.; Li, X. Pseudomonas aeruginosaL10: A Hydrocarbon-Degrading, Biosurfactant-Producing, and Plant-Growth-Promoting Endophytic Bacterium Isolated From a Reed (Phragmites australis). Front. Microbiol. 2018, 9, 1087. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Munder, A.; Aravind, R.; Eapen, S.J.; Tümmler, B.; Raaijmakers, J.M. Friend or foe: Genetic and functional characterization of plant endophytic Pseudomonas aeruginosa. Environ. Microbiol. 2013, 15, 764–779. [Google Scholar] [CrossRef] [PubMed]
- Steindler, L.; Bertani, I.; de Sordi, L.; Schwager, S.; Eberl, L.; Venturi, V. LasI/R and RhlI/R quorum sensing in a strain of Pseudomonas aeruginosa beneficial to plants. Appl. Environ. Microbiol. 2009, 75, 5131–5140. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.S.; Bais, H.P.; Déziel, E.; Schweizer, H.P.; Rahme, L.G.; Fall, R.; Vivanco, J.M. Pseudomonas aeruginosa–plant root interactions. Pathogenicity, biofilm formation and root exudation. Plant Physiol. 2004, 134, 320–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, P.J.; Petrini, O.; Scott, H.M.L. The distribution of some fungal and bacterial endophytes in maize (Zea mays L.). New Phytol. 1992, 122, 299–305. [Google Scholar] [CrossRef]
- Pandey, P.; Kang, S.C.; Gupta, C.P.; Maheshwari, D.K. Rhizosphere Competent Pseudomonas aeruginosa GRC1 produces characteristic siderophore and enhances growth of Indian Mustard (Brassica campestris). Curr. Microbiol. 2005, 51, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Khan, S.; Iqbal, S.; Mirza, M.S.; Khan, Q.M. Inoculation method affects colonization and activity of Burkholderia phytofirmans PsJN during phytoremediation of diesel-contaminated soil. Int. Biodeterior. Biodegrad. 2013, 85, 331–336. [Google Scholar] [CrossRef]
- Mendez-Castro, F.A.; Alexander, M. Method for Establishing a Bacterial Inoculum on Corn Roots. Appl. Environ. Microbiol. 1983, 45, 248–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, S.R.; Kamble, U.R.; Sripathy, K.V.; Bhaskar, K.U.; Singh, D.P. Seed Bio-priming for Biotic and Abiotic Stress Management. In Microbial Inoculants in Sustainable Agricultural Productivity; Singh, D.P., Ed.; Springer: New Delhi, India, 2016; pp. 218–228. [Google Scholar] [CrossRef]
- Reddy, P.P. (Ed.) Bio-priming of seeds. In Recent Advances in Crop Protection; Springer: New Delhi, India, 2013; pp. 83–90. [Google Scholar] [CrossRef]
- Gonzalez-Garcıa, S.; Alvarez-Perez, J.M.; de Saenz Miera, L.E.; Cobos, R.; Ibañez, A.; Dıez-Galan, A.; Garzón-Jimeno, E.; Coque, J.J.R. Developing tools for evaluating inoculation methods of biocontrol Streptomyces sp. strains into grapevine plants. PLoS ONE 2019, 14, e0211225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutts, S.; Benincasa, P.; Wojtyla, L.; Kubala, S.; Pace, R.; Lechowska, K.; Quinet, M.; Garnczarska, M. Seed priming: New comprehensive approaches for an old empirical technique. In New Challenges in Seed Biology-Basic and Translational Research Driving Seed Technology; Tech. Open: Rijeka, Croatia, 2016; pp. 1–46. [Google Scholar]
- Mahmood, A.; Turgay, O.C.; Farooq, M.; Hayat, R. Seed biopriming with plant growth promoting rhizobacteria: A review. FEMS Microbiol. Ecol. 2016, 92, fiw112. [Google Scholar] [CrossRef]
- Singh, V.; Upadhyay, R.S.; Sarma, B.K.; Singh, H.B. Seed bio-priming with Trichoderma asperellum effectively modulate plant growth promotion in pea. Int. J. Agric. Environ. Biotechnol. 2016, 9, 361–365. [Google Scholar] [CrossRef]
- Yadav, R.S.; Singh, V.; Pal, S.; Meena, S.K.; Meena, V.S.; Sarma, B.K.; Singh, H.B.; Rakshit, A. Seed bio-priming of baby corn emerged as a viable strategy for reducing mineral fertilizer use and increasing productivity. Sci. Hortic. 2018, 241, 93–99. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Rajawat, M.V.S.; Singh, S.; Tyagi, S.P.; Saxena, A.K. A modified plate assay for rapid screening of potassium-solubilizing bacteria. Pedosphere 2016, 26, 768–773. [Google Scholar] [CrossRef]
- Sharma, P.; Kunawat, K.C.; Kaur, S.; Kaur, N. Assessment of zinc solubilization by endophytic bacteria in legume rhizosphere. Indian J. Appl. Res. 2014, 4, 439–441. [Google Scholar] [CrossRef]
- Cappuccino, J.C.; Sherman, N. (Eds.) Ammonia production. In Microbiology: A Laboratory Manual, 3rd ed.; Benjamin/Cummings Pub Co.: Redwood City, CA, USA, 1992; pp. 125–179. [Google Scholar]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Mishra, S.; Behera, N. Amylase activity of a starch degrading bacteria isolated from soil receiving kitchen wastes. Afr. J. Biotechnol. 2008, 7, 3326–3331. [Google Scholar]
- Meddeb-Mouelhi, F.; Moisan, J.K.; Beauregard, M. A comparison of plate assay methods for detecting extracellular cellulase and xylanase activity. Enzym. Microb. Technol. 2014, 66, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Kuddus, M.; Ahmad, I.Z. Isolation of novel chitinolytic bacteria and production optimization of extracellular chitinase. J. Genet. Eng. Biotechnol. 2013, 11, 39–46. [Google Scholar]
- Soares, M.M.; Silva, R.D.; Gomes, E. Screening of bacterial strains for pectinolytic activity: Characterization of the polygalacturonase produced by Bacillus sp. Revista de Microbiologia 1999, 30, 299–303. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, U.B.; Trivedi, M.; Sahu, P.K.; Paul, S.; Paul, D.; Saxena, A.K. Seed Biopriming with Salt-Tolerant Endophytic Pseudomonas geniculata Modulate Biochemical Responses Provides Ecological Fitness in Maize (Zea mays L.) Grown in Saline Sodic Soil. Int. J. Environ. Res. Public Health 2020, 17, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadasivam, S.; Manickam, A. Biochemical Methods; New Age International (P) Ltd.: New Delhi, India, 1996; p. 256. [Google Scholar]
- Thimmaiah, S.R. Standard Methods of Biochemical Analysis; Kalyani Publishers: New Delhi, India, 2012; pp. 421–426. [Google Scholar]
- Hooda, K.S.; Khokhar, M.K.; Parmar, H.; Gogoi, R.; Joshi, D.; Sharma, S.S.; Yadav, O.P. Banded Leaf and Sheath Blight of Maize: Historical Perspectives, Current Status and Future Directions. Proc. Natl. Acad. Sci. USA 2017, 87, 1041. [Google Scholar] [CrossRef]
- Kohlmeier, S.; Mancuso, M.; Tecon, R.; Harms, H.; van der Meer, J.R.; Wells, M. Bioreporters: Gfp versus lux revisited and single-cell response. Biosens. Bioelectron. 2007, 22, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Chalupowicz, L.; Zellermann, E.-M.; Fluegel, M.; Dror, O.; Eichenlaub, R.; Gartemann, K.-H.; Savidor, A.; Sessa, G.; Iraki, N.; Barash, I.; et al. Colonization and movement of GFP-labeled Clavibacter michiganensis subsp. Michiganensis during tomato infection. Phytopathology 2012, 102, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Qazi, S.N.A.; Rees, C.E.D.; Mellits, K.H.; Hill, P.J. Development of gfp vectors for expression in Listeria monocytogenes and other low G+C gram positive bacteria. Microb. Ecol. 2001, 41, 301–309. [Google Scholar] [CrossRef]
- Prieto, P.; Schilirò, E.; Maldonado-González, M.M.; Valderrama, R.; Barroso-Albarracín, J.B.; Mercado-Blanco, J. Root hairs play a key role in the endophytic colonization of olive roots by Pseudomonas spp. with biocontrol activity. Microb. Ecol. 2011, 62, 435–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, e669–e678. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, L.; Meng, H.; Wen, H.; Fan, Y.; Zhao, J. Maize ABP9 enhances tolerance to multiple stresses in transgenic Arabidopsis by modulating ABA signaling and cellular levels of reactive oxygen species. Plant Mol. Biol. 2001, 75, 365–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triantaphylides, C.; Krischke, M.; Hoeberichts, F.A.; Ksas, B.; Gresser, G.; Havaux, M.; van Breusegem, F.; Mueller, M.J. Singlet oxygen is the major reactive oxygen species involved in photo-oxidative damage to plants. Plant Physiol. 2008, 148, 960–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- De Sousa, D.P.F.; Braga, B.B.; Gondim, F.A.; Gomes-Filho, E.; Martins, K.; de Brito, P.O.B. Increased drought tolerance in maize plants induced by H2O2 is closely related to an enhanced enzymatic antioxidant system and higher soluble protein and organic solutes contents. Theor. Exp. Plant Physiol. 2016, 28, 297–306. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S.L.; Harman, G.E.; Monte, E. Translational Research on Trichoderma: From’ Omics to the Field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Zhang, L.; Xia, H.; Yu, C.; Dou, K.; Li, Y.; Chen, J. Omics for understanding synergistic action of validamycin A and Trichoderma asperellum GDFS1009 against maize sheath blight pathogen. Sci. Rep. 2017, 7, 40140. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Fei, B.; He, J.; Zhou, M.; Zhang, D.; Pan, L.; Li, S.; Liang, Y.; Wang, L.; Zhu, J.; et al. Transcriptome analysis reveals the host selection fitness mechanisms of the Rhizoctonia solani AG1IA pathogen. Sci. Rep. 2017, 7, 10120. [Google Scholar] [CrossRef] [PubMed]
- Zeilinger, S.; Omann, M. Trichoderma biocontrol: Signal transduction pathways involved in host sensing and mycoparasitism. Gene Regul. Syst. Biol. 2007, 8, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.P. Occurring rule and integrated protection technique of corn sheath disease. J. Maize Sci. 2000, 8, 71–72. [Google Scholar]
- Liu, W.; Liu, J.; Ning, Y.; Ding, B.; Wang, X.; Wang, Z.; Wang, G.L. Recent progress in understanding PAMP-and effector-triggered immunity against the rice blast fungus Magnaporthe oryzae. Mol. Plant 2013, 6, 605–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, A.; Lin, R.; Zhang, D.; Qin, P.; Xu, L.; Ai, P.; Ding, L.; Wang, Y.; Chen, Y.; Liu, Y. The evolution and pathogenic mechanisms of the rice sheath blight pathogen. Nat. Commun. 2013, 4, 1424. [Google Scholar] [CrossRef] [PubMed]
- Ruocco, M.; Lanzuise, S.; Vinale, F.; Marra, R.; Turrà, D.; Woo, S.L.; Lorito, M. Identification of a new biocontrol gene in Trichoderma atroviride: The role of an ABC transporter membrane pump in the interaction with different plant-pathogenic fungi. Mol. Plant Microbe 2009, 22, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Antico, C.J.; Colon, C.; Banks, T.; Ramonell, K.M. Insights into the role of jasmonic acid-mediated defenses against necrotrophic and biotrophic fungal pathogens. Front. Biol. 2012, 7, 48–56. [Google Scholar] [CrossRef]
- Backer, R.; Naidoo, S.; van den Berg, N. The Nonexpressor of Pathogenesis-Related Genes 1 (NPR1) and Related Family: Mechanistic Insights in Plant Disease Resistance. Front. Plant Sci. 2019, 10, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nic-Matos, G.; Narváez, M.; Peraza-Echeverría, S.; Sáenz, L.; Oropeza, C. Molecular cloning of two novel NPR1 homologue genes in coconut palm and analysis of their expression in response to the plant defense hormone salicylic acid. Genes Genom. 2017, 39, 1007–1019. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; de Wit, P.J.G.M. Fungal effector proteins. Annu. Rev. Phytopathol. 2009, 47, 233–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species-opportunistic: Avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, K.; Nishizawa, Y.; Tabei, Y.; Hibi, T.; Nakajima, M.; Akutsu, K. Detailed analysis of rice chitinase gene expression in transgenic cucumber plants showing different levels of disease resistance to gray mold (Botrytis cinerea). Plant Sci. 2002, 162, 655–662. [Google Scholar] [CrossRef]
- Muthukrishnan, S.; Liang, G.H.; Trick, H.N.; Gill, B.S. Pathogenesis related proteins and their genes in cereals. Plant Cell Tissue Organ Cult. 2001, 64, 93–114. [Google Scholar] [CrossRef]
- Karmakar, S.; Molla, K.A.; Das, K.; Sarkar, S.N.; Datta, S.K.; Datta, K. Dual gene expression cassette is superior than single gene cassette for enhancing sheath blight tolerance in transgenic rice. Sci. Rep. 2017, 7, 7900. [Google Scholar] [CrossRef] [Green Version]
- Molla, K.A.; Karmakar, S.; Chanda, P.K.; Ghosh, S.; Sarkar, S.N.; Datta, S.K.; Datta, K. Rice oxalate oxidase gene driven by green tissue-specific promoter increases tolerance to sheath blight pathogen (Rhizoctonia solani) in transgenic rice. Mol. Plant Pathol. 2013, 14, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, M.V.; Salguero, J.; Lloret, P.G. Auxin Modulated Initiation of Lateral Roots Is Linked to Pericycle Cell Length in Maize. Front. Plant Sci. 2019, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.S.; Webb, R.P.; Holaday, A.S.; Allen, R.D. Overexpression of superoxide dismutase protects plants from oxidative stress. Plant Physiol. 1993, 103, 1067–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Gene Name | Primers (5′–3′) |
|---|---|---|
| 1. | zmPR-1 | TCAGTCATGCCGTTCAGCTT |
| TTGTCCGCGTCCAGGAA | ||
| 2. | zmPR-10 | CAACCCGGAAGCCTACAACTAG |
| GAAATCCGTTCCCCATCGA | ||
| 3. | zmACTIN | GGGATTGCCGATCGTATGAG |
| GAGCCACCGATCCAGACACT |
| Treatments | Fresh wt. of Shoot (g) | Fresh wt. of Root (g) | Dry wt. of Shoot (g) | Dry wt. of Root (g) |
|---|---|---|---|---|
| T1 | 25.15 ± 1.05 f | 20.22 ± 0.45 e | 7.50 ± 0.15 e | 4.00 ± 0.05 f |
| T2 | 32.42 ± 0.96 d | 24.66 ± 0.66 c | 8.54 ± 0.11 d | 5.05 ± 0.06 d |
| T3 | 44.25 ± 1.00 b | 28.45 ± 1.05 b | 10.25 ± 0.15 b | 6.75 ± 0.10 b |
| T4 | 30.25 ± 1.10 e | 22.75 ± 1.25 d | 8.02 ± 0.09 d | 4.78 ± 0.11 e |
| T5 | 38.67 ± 0.75 c | 25.25 ± 0.85 c | 9.15 ± 0.07 c | 5.86 ± 0.05 c |
| T6 | 50.46 ± 1.33 a | 30.40 ± 1.12 a | 12.03 ± 0.10 a | 7.20 ± 0.08 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Singh, U.B.; Malviya, D.; Paul, S.; Sahu, P.K.; Trivedi, M.; Paul, D.; Saxena, A.K. Seed Biopriming with Microbial Inoculant Triggers Local and Systemic Defense Responses against Rhizoctonia solani Causing Banded Leaf and Sheath Blight in Maize (Zea mays L.). Int. J. Environ. Res. Public Health 2020, 17, 1396. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17041396
Singh S, Singh UB, Malviya D, Paul S, Sahu PK, Trivedi M, Paul D, Saxena AK. Seed Biopriming with Microbial Inoculant Triggers Local and Systemic Defense Responses against Rhizoctonia solani Causing Banded Leaf and Sheath Blight in Maize (Zea mays L.). International Journal of Environmental Research and Public Health. 2020; 17(4):1396. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17041396
Chicago/Turabian StyleSingh, Shailendra, Udai B. Singh, Deepti Malviya, Surinder Paul, Pramod Kumar Sahu, Mala Trivedi, Diby Paul, and Anil Kumar Saxena. 2020. "Seed Biopriming with Microbial Inoculant Triggers Local and Systemic Defense Responses against Rhizoctonia solani Causing Banded Leaf and Sheath Blight in Maize (Zea mays L.)" International Journal of Environmental Research and Public Health 17, no. 4: 1396. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17041396