Effect of Molybdenum on Plant Physiology and Cadmium Uptake and Translocation in Rape (Brassica napus L.) under Different Levels of Cadmium Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Plant Sampling and Analysis
2.3.1. Measurement of Chlorophyll Fluorescence Parameters
2.3.2. Measurement of Physiological Parameters and Biomass
2.3.3. Quantification of Inorganic Cations and Heavy Metals
2.4. Data Analysis
3. Results
3.1. Effects of Mo on Plant Physiological Parameters and Biomass in Cd-Stressed Rape
3.1.1. Fresh and Dry Weights of Shoots and Roots
3.1.2. Physiological Parameters
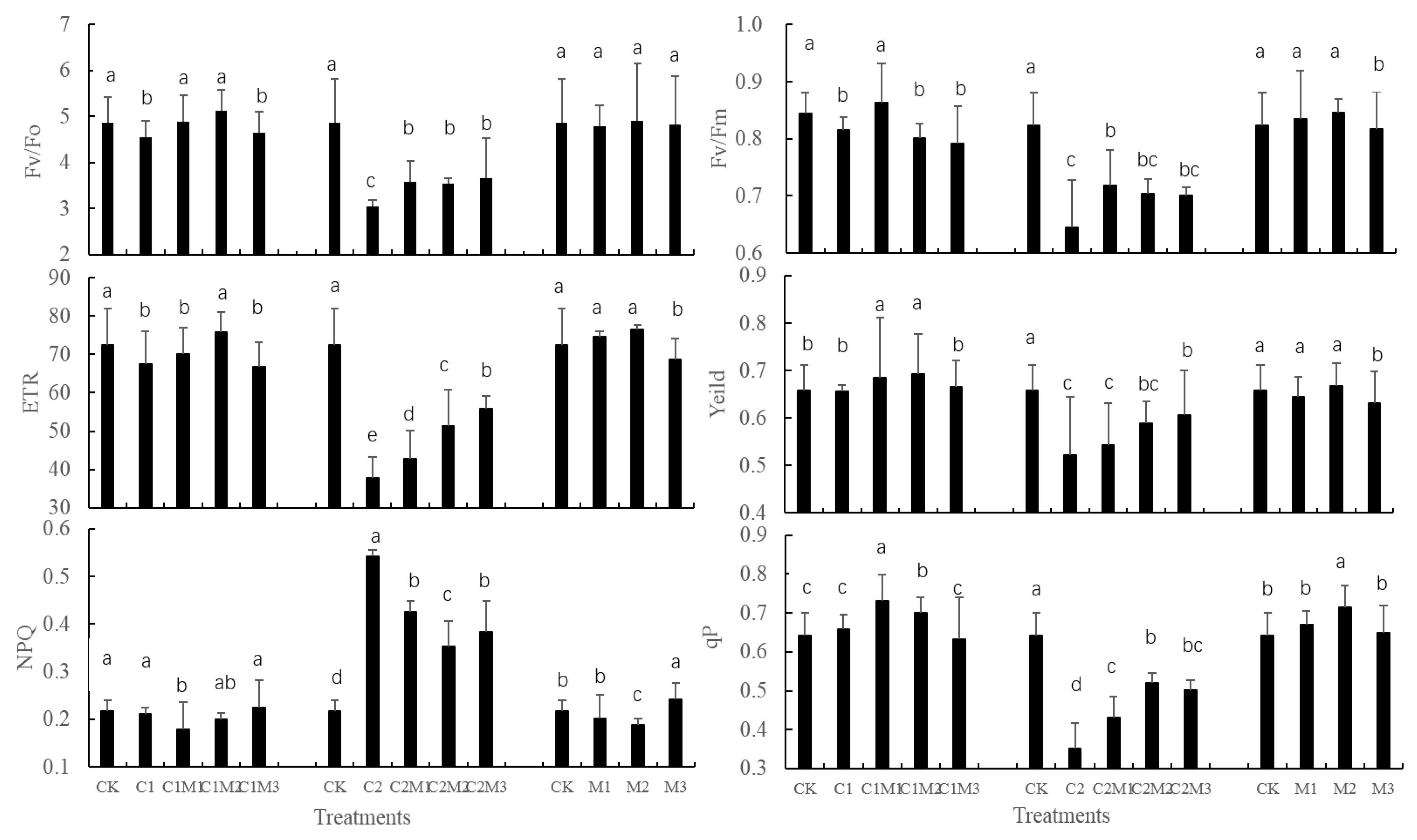
3.2. Effect of Mo on Chlorophyll Fluorescence Parameters in Cd-stressed Rape
3.3. Effect of Mo on the Uptake of Inorganic Cations in Cd-Stressed Rape
3.4. Effect of Mo on Cd Uptake, Enrichment, and Translocation in Cd-Stressed Rape
4. Discussion
4.1. Exogenous Mo Benefits Plant Physiology and Biomass in Cd-Stressed Rape
4.2. Exogenous Mo Improves Chlorophyll Fluorescence Parameters in Cd-Stressed Rape
4.3. Exogenous Mo Facilitates Cation Uptake in Cd-Stressed Rape
4.4. Exogenous Mo Modulates Cd Uptake, Enrichment, and Translocation in Rape
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Guo, Y.; Yang, J.; Guo, J.M.; Yang, J.X.; Zheng, G.D.; Li, Z.T.; Chen, T.B.; Yu, B.; Ni, L.; Wen, Q.; et al. Mechanism of nitrogen, phosphrous and potassium combined application promote absorption, transportation and accumulation of Pb in sunflower (Helianthns annuus L.). J. Plant Nutri. Ferti. 2019, 25, 1998–2008. [Google Scholar] [CrossRef]
- Wan, Y.N.; Yu, Y.; Wang, Q.; Qiao, Y.H.; Li, H.F. Cadmium uptake dynamics and translocation in rice seedling: Influence of different forms of selenium. Ecotoxicol. Environ. Saf. 2016, 133, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Camara, Y.A.; Wan, Y.N.; Yu, Y.; Wang, Q.; Li, H.F. Effect of selenium on uptake and translocation of arsenic in rice seedlings (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2018, 148, 869–875. [Google Scholar] [CrossRef]
- Handa, N.; Kohli, S.K.; Thukral, A.K.; Bhardwaj, R.; Ahmad, P. Protective role of selenium against chromium stress involving metabolites and essential elements in Brassica juncea L. seedlings. 3Biotech 2018, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Z.; Hu, Y.; Liu, Y.X. Heavy metal accumulation in iron plaque and growth of rice plants upon exposure to single and combined contamination by copper, cadmium and lead. Acta Ecolo. Sin. 2009, 29, 320–326. [Google Scholar] [CrossRef]
- Rollinson, C.L. The chemistry of Chromium, Molybdenum and Tungsten; Pergamon Press: New York, NY, USA, 1975. [Google Scholar] [CrossRef]
- Zhou, S.H.; Zhang, C.Y.; Xiao, Q.Y.; Zhuang, Y.; Gu, X.L.; Yang, F.; Xing, C.H.; Hu, G.L.; Cao, H.B. Effects of different levels of molybdenum on rumen microbiota and trace elements changes in tissues from goats. Biol. Trace Elem. Res. 2016, 174, 85–92. [Google Scholar] [CrossRef]
- Xu, T.; Sun, C.B. Aerosol flotation of low-grade refractory molybdenum ores. Int. J. Miner. Metall. Mater. 2012, 12, 11–16. [Google Scholar] [CrossRef]
- Wu, G.M.; Hu, P.J.; Zhou, J.W.; Dong, B.; Wu, L.H.; Luo, Y.M.; Christie, P. Sulfur application combined with water management enhances phytoextraction rate and decreases rice cadmium uptake in a Sedum plumbizincicola - Oryza sativa rotation. Plant Soil 2019, 440, 539–549. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Responses of nitric oxide and hydrogen sulfide in regulating oxidative defence system in wheat plants grown under cadmium stress. Physiol. Plant. 2020, 168, 345–360. [Google Scholar] [CrossRef]
- Filek, M.; Gzyl-Malcher, B.; Zembala, M.; Bednarska, E.; Laggner, P.; Kriechbaum, M. Effect of selenium on characteristics of rape chloroplasts modified by cadmium. J. Plant Physiol. 2010, 167, 28–33. [Google Scholar] [CrossRef]
- Li, Y.P.; Wang, S.L.; Zhang, Q.; Zang, F.; Nan, Z.R.; Sun, H.L.; Huang, W.; Bao, L.L. Accumulation, interaction and fractionation of fluoride and cadmium in sierozem and oilseed rape (Brassica napus L.) in northwest China. Plant Physiol. Biochem. 2018, 127, 457. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Su, M.Y.; Chen, Y.H.; Lin, F.F.; Luo, D.; Gao, S.F. Transfer characteristics of cadmium and lead from soil to the edible parts of six vegetable species in southeastern China. Environ. Pollut. 2006, 144, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Chunilall, V.; Kindness, A.; Jonnalagadda, S.B. Heavy metal uptake by spinach leaves grown on contaminated soils with lead, mercury, cadmium, and nickel. J. Environ. Sci. Health B 2004, 39, 473–481. [Google Scholar] [CrossRef]
- Chen, Y.X.; He, Y.F.; Luo, Y.M.; Yu, Y.L.; Lin, Q.; Wong, M.H. Physiological mechanism of plant roots exposed to cadmium. Chemosphere 2003, 50, 789–793. [Google Scholar] [CrossRef]
- Liu, J.S.; Shi, H.; Li, Y.Y. Effects of Cd2+ stress on photosynthesis and chlorophyll fluorescence characteristics of cucumber seedlings. Resear. Soil Water Conser. 2011, 18, 187–190, 196. [Google Scholar]
- Carrier, P.; Baryla, A.; Havaux, M. Cadmium distribution and microlocalization in oilseed rape (Brassica napus) after long-term growth on cadmium-contaminated soil. Planta 2003, 216, 939–950. [Google Scholar] [CrossRef]
- Ali, B.; Deng, X.Q.; Hu, X.Q.; Gill, R.A.; Ali, S.; Wang, S.; Zhou, W.J. Deteriorative effects of cadmium stress on antioxidant system and cellular structure in germinating seeds of Brassica napus L. J. Agric. Sci. Technol. 2018, 17, 63–74. [Google Scholar]
- Ali, B.; Qian, P.; Jin, R.; Ali, S.; Khan, M.; Aziz, R.; Zhou, W. Physiological and ultra-structural changes inbrassica napus seedlings induced by cadmium stress. Biol. Plant. 2014, 58, 131–138. [Google Scholar] [CrossRef]
- Filek, M.; Kocielniak, J.; Abanowska, M.; Bednarska, E.; Ewa, B. Selenium-induced protection of photosynthesis activity in rape (brassica napus) seedlings subjected to cadmium stress. fluorescence and EPR measurements. Photosynth. Res. 2010, 105, 27–37. [Google Scholar] [CrossRef]
- Wu, Z.C.; Zhao, X.H.; Sun, X.C.; Tan, Q.L.; Tang, Y.F.; Nie, Z.J.; Qu, C.J.; Chen, Z.X.; Hu, C.X. Antioxidant enzyme systems and the ascorbate-glutathione cycle as contributing factors to cadmium accumulation and tolerance in two oilseed rape cultivars (Brassica napus L.) under moderate cadmium stress. Chemosphere 2015, 138, 526–536. [Google Scholar] [CrossRef]
- Yan, H.; Filardo, F.; Hu, X.T.; Zhao, X.M.; Fu, D.H. Cadmium stress alters the redox reaction and hormone balance in oilseed rape (Brassica napus L.) leaves. Environ. Sci. Pollut. Res. 2016, 23, 3758–3769. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.G.; Xiao, X.; Yan, G.X.; Feng, T.T.; Liu, J.; Wu, X.M. Development of evaluation method for cadmium tolerance in Brassica napus seedlings. Chin. J. Oil Crop Sci. 2017, 39, 47–54. [Google Scholar] [CrossRef]
- Filek, M.; Keskinen, R.; Hartikainen, H.; Szarejko, I.; Janiak, A.; Miszalski, Z.; Golda, A. The protective role of selenium in rape seedlings subjected to cadmium stress. J. Plant Physiol. 2008, 165, 833–844. [Google Scholar] [CrossRef]
- Li, B.; Yang, J.X.; Chen, S.B.; Li, J.M.; Ma, Y.B. Field evidence of cadmium phytoavailability decreased effectively by rape straw and/or red mud with zinc sulphate in a Cd-contaminated calcareous soil. PLoS ONE 2014, 9, e109967. [Google Scholar] [CrossRef]
- Balík, J.; Pavliková, D.; Tlustoš, P.; Sýkora, K.; Černý, J. The fluctuation of molybdenum content in oilseed rape plants after the application of nitrogen and sulphur fertilizers. Plant Soil Environ. 2006, 52, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.Y.; Sun, X.C.; Hu, C.X.; Tan, Q.L.; Zhao, X.H.; Xu, S.J. Effects of tungsten on uptake, transport and subcellular distribution of molybdenum in oilseed rape at two different molybdenum levels. Plant Sci. 2017, 256, 87–93. [Google Scholar] [CrossRef]
- Ide, Y.; Kusano, M.; Oikawa, A.; Fukushima, A.; Tomatsu, H.; Saito, K.; Hirai, M.Y.; Fujiwara, T. Effects of molybdenum deficiency and defects in molybdate transporter MOT1 on transcript accumulation and nitrogen/sulphur metabolism in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 1483–1497. [Google Scholar] [CrossRef]
- Yaryura, P.; Cordon, G.; Leon, M.; Kerber, N.; Lagorio, M.G. Effect of phosphorus deficiency on reflectance and chlorophyll fluorescence of cotyledons of oilseed rape (Brassica napus L.). J. Agron. Crop Sci. 2009, 195, 186–196. [Google Scholar] [CrossRef]
- Hura, K.; Hura, T.; Dziurka, K.; Dziurka, M. Carbohydrate, phenolic and antioxidant level in relation to chlorophyll a content in oilseed winter rape (Brassica napus L.) inoculated with Leptosphaeria maculans. Eur. J. Plant Pathol. 2015, 143, 291–303. [Google Scholar] [CrossRef] [Green Version]
- Li, X.D.; Zhuang, K.Y.; Liu, Z.M.; Yang, D.Y.; Ma, N.N.; Meng, Q.W. Overexpression of a novel NAC-type tomato transcription factor, SINAM1, enhances the chilling stress tolerance of transgenic tobacco. J. Plant Physiol. 2016, 204, 54–65. [Google Scholar] [CrossRef]
- Teklić, T.; Spoljarević, M.; Stanisavljević, A.; Lisjak, M.; Vinković, T.; Parađiković, N.; Andrić, L.; Hancock, J.T. Assessment of the influence of different sample processing and cold storage duration on plant free proline content analyses. Phytochem. Anal. 2010, 21, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.L.; Zhang, F.; Ahmed, Z.I.; Rasheed, M.; Naeem, M.S.; Ye, Q.F.; Zhou, W.J. Differential morphological and physiological responses of two oilseed brassica species to a new herbicide zj0273 used in rapeseed fields. Pest. Biochem. Physiol. 2010, 98, 1–8. [Google Scholar] [CrossRef]
- Bela, K.; Horváth, E.; Galléa, Á.; Szabados, L.; Tari, I.; Csiszár, J. Plant glutathione peroxidases: Emerging role of the antioxidant enzymes in plant development and stress responses. J. Plant Physiol. 2015, 176, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.J.; Huang, Q.; Yan, B.; Wang, Y.; Qian, Z.Y.; Pan, J.X.; Kai, G.Y. Molecular cloning and expression analysis of a Cu/Zn SOD gene (BcCSD1) from Brassica campestris ssp. Chinensis. Food Chem. 2015, 186, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Samarah, N.H.; Mullen, R.E.; Anderson, I. Soluble sugar contents, germination, and vigor of soybean seeds in response to drought stress. J. New Seeds 2009, 10, 63–73. [Google Scholar] [CrossRef]
- Chen, H.; Wang, Z.B.; Chang, Q.Y.; Wang, W.; Fan, C.L.; Pang, G.F. Determination of trace elements in rape honey and its corresponding rape flower and dtem by ICP-MS. Spectrosc. Spectral Anal. 2014, 34, 548–552. [Google Scholar] [CrossRef]
- Kira, C.S.; Maio, F.D.; Maihara, V.A. Comparison of partial digestion procedures for determination of Ca, Cr, Cu, Fe, K, Mg, Mn, Na, P, and Zn in milk by inductively coupled plasma-optical emission spectrometry. J. AOAC. Int. 2004, 87, 151–156. [Google Scholar] [CrossRef]
- Fayiga, A.O.; Ma, L.Q.; Cao, X. Effects of heavy metals on growth and arsenic accumulation in the arsenic hyperaccumulator Pteris Vittata L. Environ. Pollut. 2004, 132, 289–296. [Google Scholar] [CrossRef]
- Ren, J.H.; Sun, H.J.; Wang, S.F.; Luo, J.; Ma, L.Q. Interactive effects of mercury and arsenic on their uptake, speciation and toxicity in rice seedling. Chemosphere 2014, 117, 737–744. [Google Scholar] [CrossRef]
- Han, Y.L.; Yuan, H.Y.; Huang, S.H.; Guo, Z.; Xia, B. Cadmium tolerance and accumulation by two species of iris. Ecotoxicology 2007, 16, 557–563. [Google Scholar] [CrossRef]
- Singh, S.; Eapen, S.; D’ Souza, S.F. Cadmium accumulation its influence on lipid peroxidation and antioxidative system in an aquatic plant, Bacopa monnieri L. Chemosphere 2006, 62, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Schat, H.; Sharma, S.S.; Vooijs, R. Heavy metal-induced accumulation of free proline in a metal-tolerant and a nontolerant ecotype of Silene vulgaris. Physiol. Plant. 1997, 101, 477–482. [Google Scholar] [CrossRef]
- Stroud, J.L.; Zhao, F.J.; Buchner, P.; Shinmachi, F.; McGrath, S.P.; Abecassis, J.; Hawkesford, M.J.; Shewry, P.R. Impacts of sulphur nutrition on selenium and molybdenum concentrations in wheat grain. J. Cereal Sci. 2010, 52, 111–113. [Google Scholar] [CrossRef]
- Yang, H.Y.; Shi, G.X.; Xu, Q.S.; Wang, H.X. Cadmium effects on mineral nutrition and stress in potamogeton crispus. Russ. J. Plant Physiol. 2011, 58, 253–260. [Google Scholar] [CrossRef]
- Bonifacio, A.; Carvalho, F.E.L.; Martins, M.O.; Neto, M.C.L.; Cunha, J.R.; Ribeiro, C.W.; Margis-Pinheiro, M.; Silveira, J.A.G. Silenced rice in both cytosolic ascorbate peroxidases displays pre-acclimation to cope with oxidative stress induced by 3-aminotriazole-inhibited catalase. J. Plant Physiol. 2016, 201, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.F.; Wang, Y.B.; Li, J.L. Impacts of Cu and Zn pollution on rape (Brassica chinensis L.) growth and its accumulation effect of heavy metals in paddy soil. Eco. Environ. 2011, 20, 1470–1477. [Google Scholar] [CrossRef]
- Wong, Y.S.; Luo, G.B.; Kwan, K.M.F. Peroxidation damage of oxygen free radicals induced by cadmium to plant. Acta Bot. Sin. 1997, 39, 36–40. [Google Scholar]
- Boo, H.O.; Hwang, S.J.; Bae, C.S.; Park, S.H.; Heo, B.G.; Gorinstein, S. Extraction and characterization of some natural plant pigments. Ind. Crop. Prod. 2012, 40, 129–135. [Google Scholar] [CrossRef]
- Ma, Y.L.; Wang, H.F.; Wang, P.; Yu, C.G.; Luo, S.Q.; Zhang, Y.F.; Xie, Y.F. Effects of cadmium stress on the antioxidant system and chlorophyll fluorescence characteristics of two Taxodium clones plant. Cell Reports 2018, 37, 1547–1555. [Google Scholar] [CrossRef]
- Gao, G.Q.; Jian, M.F.; Lu, L.; Ji, Y.; Wang, X.L.; Wang, Y.; Wang, X.R. Effects of Cu2+ or Cd2+ stress on photosynthetic pigment and photosynthetic fluorescence characteristics of Potamogeton malaianus. Chin. J. Appl. Environ. Biol. 2019, 25, 517–523. [Google Scholar] [CrossRef]
- Li, W.X.; Wang, Z.Y.; Zhang, F.S.; Han, X.R. Effect of molybdenum on growth of winter wheat seedlings induced by low temperature. J. Plant Nutri. Ferti. 2000, 6, 312–317. [Google Scholar] [CrossRef]
- Kevresan, S.; Petrovic, N.; Popovic, M. Nitrogen and protein metabolism in young peaplants as affected by different concentrations of nikel, cadmium, lead, and molybdenum. J. Plant Nutr. 2001, 24, 1633–1644. [Google Scholar] [CrossRef]
- Dong, J.; Wu, F.B.; Zhang, G.P. Influence of cadmium on antioxidant capacity and four microelement concentrations in tomato seedlings (Lycopersicon esculentum). Chemosphere 2006, 64, 1659–1666. [Google Scholar] [CrossRef]
- Iannone, M.F.; Groppa, M.D.; Benavides, M.P. Cadmium induces different biochemical responses in wild type and catalase-deficient tobacco plants. Environ. Exp. Bot. 2015, 109, 201–211. [Google Scholar] [CrossRef]
- Shah, K.; Kumar, R.G.; Verma, S.; Dubey, R.S. Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings. Plant Sci. 2001, 161, 1135–1144. [Google Scholar] [CrossRef]
- Meng, H.B.; Hua, S.B.; Shamsi, I.H.; Jilani, G.; Li, Y.L.; Jiang, L.X. Cadmium-induced stress on the seed germination and seedling growth of brassica napus L. and its alleviation through exogenous plant growth regulators. Plant Growth Regul. 2009, 58, 47–59. [Google Scholar] [CrossRef]
- Sun, J.; Bankston, J.R.; Payandeh, J.; Hinds, T.R.; Zagotta, W.N.; Zheng, N. Crystal structure of a plant dual-affinity nitrate transporter. Nature 2014, 507, 73–77. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.S.; Cortez, P.A.; Almeida, A.F.; Prasad, M.N.V.; Franca, M.G.C.; Cunda, M.; Jesus, R.M.; Mangabeira, P.A.O. Morphology, ultrastructure, and element uptake in Calophyllum brasiliense Cambess. (Calophyllaceae J. Agardh) seedlings under cadmium exposure. Environ. Sci. Pollut. Res. 2017, 24, 15576–15588. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Lukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.E.; Hu, C.X.; Sun, X.C.; Tan, Q.L.; Nie, Z.J.; Hu, X.M. Interactive effects of molybdenum and phosphorus fertilizers on photosynthetic characteristics of seedlings and grain yield of Brassica napus. Plant Soil 2010, 326, 345–353. [Google Scholar] [CrossRef]
- Kumchai, J.; Huang, J.Z.; Lee, C.Y.; Chen, C.Y.; Chin, S.W. Proline partially overcomes excess molybdenum toxicity in cabbage seedlings grown in vitro. Genet. Mol. Res. 2013, 12, 5589–5601. [Google Scholar] [CrossRef]
- Salvatori, E.; Fusaro, L.; Gottardini, E.; Pollastrini, M.; Goltsev, V.; Strasser, R.J.; Bussotti, F. Plant stress analysis: Application of prompt, delayed chlorophyll fluorescence and 820 nm modulated reflectance. insights from independent experiments. Plant Physiol. Biochem. 2014, 85, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Nesterenko, T.V.; Tikhomirov, A.A.; Shikhov, V.N. Ontogenetic approach to the assessment of plant resistance to prolonged stress using chlorophyll fluorescence induction method. Photosynthetica 2006, 44, 321–332. [Google Scholar] [CrossRef]
- Liu, H.E.; Hu, C.X.; Sun, X.C.; Nie, Z.J.; Tan, Q.L. Effect of molybdenum nutrition on growth of Brassica napus. Hubei Agricul. Sci. 2011, 50, 1305–1308. [Google Scholar] [CrossRef]
- Dezhban, A.; Shirvany, A.; Attarod, P.; Delshad, M.; Khoshnevis, M. Cadmium and lead effects on chlorophyll fluorescence, chlorophyll pigments and proline of robinia pseudoacacia. J. For. Res. 2015, 26, 323–329. [Google Scholar] [CrossRef]
- Lin, L.; Zhou, W.H.; Dai, H.X.; Cao, F.B.; Zhang, G.P.; Wu, F.B. Selenium reduces cadmium uptake and mitigates cadmium toxicity in rice. J. Hazard. Mater. 2012, 235–236, 343–351. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Li, Q.S.; Li, H.; Chen, Y. Absorption and accumulation of heavy metals in water spinach, and their relationships with nutrient elements. J. Eco. Rural Environ. 2013, 29, 225–229. [Google Scholar] [CrossRef]
- Zembala, M.; Filek, M.; Walas, S.; Mrowiec, H.; Kornaś, A.; Miszalski, Z.; Hartikainen, H. Effect of selenium on macro- and microelement distribution and physiological parameters of rape and wheat seedlings exposed to cadmium stress. Plant Soil 2010, 329, 457–468. [Google Scholar] [CrossRef]
- Edelstein, M.; Ben-Hur, M. Heavy metals and metalloids: Sources, risks and strategies to reduce their accumulation in horticultural crops. Sci. Hortic. 2018, 234, 431–444. [Google Scholar] [CrossRef]
- National Health commission of the People’s Republic of China; State Administrtion for Market Regulation. GB 2762-2017 National Food Safety Standard, Limits for Pollution of Food; China Standards Press: Beijing, China, 2017; pp. 4–5.
- Mourato, M.P.; Moreira, I.N.; Leitão, I.; Pinto, F.R.; Sales, J.R.; Martins, L.L. Effect of heavy metals in plants of the genus brassica. Int. J. Mol. Sci. 2015, 16, 17975–17998. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Khan, S.; Ding, X.D.; Khan, A.; Alam, M. The effects of biochar and rice husk on adsorption and desorption of cadmium on to soils with different water conditions (upland and saturated). Chemosphere 2017, 193, 1120–1126. [Google Scholar] [CrossRef]
- Yang, B.B.; Yang, C.; Shao, Z.Y.; Wang, H.; Zan, S.T.; Zhu, M.; Zhou, S.B.; Yang, R.Y. Selenium (se) does not reduce cadmium (cd) uptake and translocation in rice (Oryza sativa L.) in naturally occurred se-rich paddy fields with a high geological background of cd. Bull. Environ. Contam. Toxicol. 2019, 103, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.J.; Hu, C.X.; Sun, X.C.; Tan, Q.L.; Liu, H.E. Effects of molybdenum on ascorbate-glutathione cycle metabolism in Chinese cabbage (Brassica campestris L. ssp. pekinensis). Plant Soil 2007, 295, 13–21. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | Cd (mg) | Mo (mg) | Treatment | Cd (mg) | Mo (mg) |
|---|---|---|---|---|---|
| CK | 0 | 0 | C2M1 | 6 | 50 |
| C1 | 0.5 | 0 | C2M2 | 6 | 100 |
| C1M1 | 0.5 | 50 | C2M3 | 6 | 200 |
| C1M2 | 0.5 | 100 | M1 | 0 | 50 |
| C1M3 | 0.5 | 200 | M2 | 0 | 100 |
| C2 | 6 | 0 | M3 | 0 | 200 |
| Treatment | MDA | Proline | Soluble Proteins | Soluble Sugars | Chlorophyll | SOD | GPX |
|---|---|---|---|---|---|---|---|
| (mmol/kgFW) | (mg/kgFw) | (mg/kg·103) | (mg/kg·103) | (mg/kg·103) | (U/g·h) | (U/g·h) | |
| CK | 14.6 ± 0.8bc | 95.2 ± 6.4c | 17.4 ± 1.2a | 66.2 ± 2.2a | 2.51 ± 0.12a | 260 ± 12b | 97.5 ± 8.9a |
| C1 | 19.7 ± 1.6a | 142 ± 5a | 15.8 ± 1.1b | 59.4 ± 2.1b | 2.14 ± 0.31c | 251 ± 15c | 71.2 ± 3.6c |
| C1M1 | 15.4 ± 2.2b | 125 ± 8ab | 14.2 ± 1.7bc | 58.1 ± 2.1b | 2.25 ± 0.13b | 256 ± 15b | 80.8 ± 7.3b |
| C1M2 | 13.7 ± 0.7c | 115 ± 4b | 17.5 ± 0.8a | 60.2 ± 1.3b | 2.39 ± 0.15b | 277 ± 18a | 85.8 ± 6.7ab |
| C1M3 | 12.8 ± 1.8c | 121 ± 6ab | 18.7 ± 0.9a | 57.2 ± 2.1b | 2.58 ± 0.31a | 242 ± 9c | 68.7 ± 2.2c |
| CK | 14.6 ± 0.8c | 95.2 ± 6.4c | 17.4 ± 1.2a | 66.2 ± 2.2a | 2.51 ± 0.12a | 260 ± 12a | 97.5 ± 8.9a |
| C2 | 42.7 ± 1.5a | 211 ± 25a | 4.3 ± 0.5c | 40.1 ± 1.7c | 1.73 ± 0.10c | 204 ± 28b | 40.4 ± 7.7bc |
| C2M1 | 39.7 ± 1.9a | 221 ± 27a | 6.4 ± 0.1c | 47.2 ± 1.5bc | 1.78 ± 0.24c | 228 ± 22ab | 45.1 ± 2.9bc |
| C2M2 | 25.7 ± 2.1b | 187 ± 14b | 9.1 ± 1.0bc | 52.4 ± 1.9b | 1.97 ± 0.15b | 242 ± 20a | 54.7 ± 3.7b |
| C2M3 | 38.7 ± 1.6a | 212 ± 12a | 12.4 ± 1.3b | 53.2 ± 2.5b | 2.06 ± 0.27b | 210 ± 18b | 35.5 ± 5.7c |
| CK | 14.6 ± 0.8b | 95.2 ± 6.4b | 17.4 ± 1.2b | 66.2 ± 2.2ab | 2.51 ± 0.12ab | 260 ± 12a | 97.5 ± 8.9a |
| M1 | 10.4 ± 1.9c | 92.3 ± 5.8b | 20.4 ± 1.3a | 68.1 ± 2.2a | 2.63 ± 0.19a | 270 ± 9a | 98.8 ± 2.8a |
| M2 | 13.7 ± 2.1b | 94.6 ± 8.5b | 22.6 ± 0.8a | 72.5 ± 1.9a | 2.70 ± 0.11a | 278 ± 19a | 102 ± 13a |
| M3 | 21.7 ± 0.8a | 109 ± 10a | 13.7 ± 1.1c | 54.4 ± 3.2b | 2.33 ± 0.30b | 232 ± 21b | 88.0 ± 5.9b |
| Treatment | K+ | Na+ | Ca2+ | Mg2+ |
|---|---|---|---|---|
| (mg/kg) | (mg/kg) | (mg/kg) | (mg/kg) | |
| CK | 31.2 ± 2.2b | 4.78 ± 0.12a | 8.45 ± 0.25a | 4.31 ± 0.12a |
| C1 | 29.7 ± 1.7b | 4.36 ± 0.12a | 8.41 ± 0.18a | 4.21 ± 0.13a |
| C1M1 | 31.8 ± 2.5b | 4.55 ± 0.23a | 8.51 ± 0.26a | 4.31 ± 0.14a |
| C1M2 | 35.7 ± 1.2a | 4.75 ± 0.13a | 8.66 ± 0.23a | 4.44 ± 0.24a |
| C1M3 | 37.2 ± 1.6a | 4.78 ± 0.22a | 8.67 ± 0.07a | 4.53 ± 0.27a |
| CK | 31.2 ± 2.2a | 4.78 ± 0.12a | 8.45 ± 0.25a | 4.31 ± 0.12a |
| C2 | 14.9 ± 1.8d | 4.01 ± 0.17a | 8.01 ± 0.19a | 3.85 ± 0.28ab |
| C2M1 | 19.5 ± 0.9c | 4.25 ± 0.22a | 7.91 ± 0.22a | 3.54 ± 0.16b |
| C2M2 | 22.5 ± 1.3bc | 4.29 ± 0.37a | 8.01 ± 0.14a | 3.67 ± 0.25b |
| C2M3 | 26.7 ± 1.5b | 4.18 ± 0.11a | 7.96 ± 0.16a | 3.95 ± 0.35ab |
| CK | 31.2 ± 2.2b | 4.78 ± 0.12a | 8.45 ± 0.25a | 4.31 ± 0.12a |
| M1 | 32.2 ± 0.5b | 4.86 ± 0.23a | 8.67 ± 0.16a | 4.36 ± 0.14a |
| M2 | 37.7 ± 1.2a | 4.92 ± 0.13a | 8.76 ± 0.13a | 4.49 ± 0.15a |
| M3 | 39.2 ± 1.6a | 4.71 ± 0.22a | 8.72 ± 0.27a | 4.57 ± 0.18a |
| Treatment | Cd (mg/kg) | Bioconcentration | Translocation Factor | Tolerance Index (%) | |
|---|---|---|---|---|---|
| Shoot | Root | Factor | |||
| CK | 0.032 ± 0.009b | 0.074 ± 0.046c | 0.442 | 0.432 | 100 |
| C1 | 0.104 ± 0.033a | 0.222 ± 0.014a | 0.441 | 0.468 | 96.4 |
| C1M1 | 0.094 ± 0.021a | 0.201 ± 0.014a | 0.412 | 0.517 | 103 |
| C1M2 | 0.044 ± 0.012b | 0.101 ± 0.024b | 0.196 | 0.436 | 113 |
| C1M3 | 0.024 ± 0.008b | 0.066 ± 0.015c | 0.122 | 0.364 | 124 |
| CK | 0.032 ± 0.009d | 0.074 ± 0.046d | 0.442 | 0.432 | 100 |
| C2 | 0.657 ± 0.052a | 1.09 ± 0.07a | 0.28 | 0.603 | 82.4 |
| C2M1 | 0.386 ± 0.025b | 0.754 ± 0.042b | 0.183 | 0.512 | 89.3 |
| C2M2 | 0.297 ± 0.024b | 0.601 ± 0.015b | 0.144 | 0.494 | 92.9 |
| C2M3 | 0.191 ± 0.012c | 0.354 ± 0.034c | 0.091 | 0.54 | 85.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Z.; Wei, X.; Wan, D.; He, W.; Wang, X.; Xiong, Y. Effect of Molybdenum on Plant Physiology and Cadmium Uptake and Translocation in Rape (Brassica napus L.) under Different Levels of Cadmium Stress. Int. J. Environ. Res. Public Health 2020, 17, 2355. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17072355
Han Z, Wei X, Wan D, He W, Wang X, Xiong Y. Effect of Molybdenum on Plant Physiology and Cadmium Uptake and Translocation in Rape (Brassica napus L.) under Different Levels of Cadmium Stress. International Journal of Environmental Research and Public Health. 2020; 17(7):2355. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17072355
Chicago/Turabian StyleHan, Zhangxiong, Xuan Wei, Dejun Wan, Wenxiang He, Xijie Wang, and Ying Xiong. 2020. "Effect of Molybdenum on Plant Physiology and Cadmium Uptake and Translocation in Rape (Brassica napus L.) under Different Levels of Cadmium Stress" International Journal of Environmental Research and Public Health 17, no. 7: 2355. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17072355