The Effect of Vitamin D3 Supplementation on Hepcidin, Iron, and IL-6 Responses after a 100 km Ultra-Marathon

 , , , ,
, , , ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Run
2.2. Biochemical Analyses
2.3. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Latunde-Dada, G.O. Iron metabolism in athletes—Achieving a gold standard. Eur. J. Haematol. 2013, 90, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Clénin, G.; Cordes, M.; Huber, A.; Schumacher, Y.O.; Noack, P.; Scales, J.; Kriemler, S. Iron deficiency in sports—Definition, influence on performance and therapy. Swiss Med. Wkly. 2015, 145, w14196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bresgen, N.; Eckl, P.M. Oxidative stress and the homeodynamics of iron metabolism. Biomolecules 2015, 5, 808–847. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Valore, E.V.; Waring, A.J.; Ganz, T. Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J. Biol. Chem. 2001, 276, 7806–7810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out Ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camaschella, C.; Pagani, A. Advances in understanding iron metabolism and its crosstalk with erythropoiesis. Br. J. Haematol. 2018, 182, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Pak, M.; Lopez, M.A.; Gabayan, V.; Ganz, T.; Rivera, S. Suppression of hepcidin during anemia requires erythropoietic activity. Blood 2006, 108, 3730–3735. [Google Scholar] [CrossRef]
- Kim, A.; Nemeth, E. New insights into iron regulation and erythropoiesis. Curr. Opin. Hematol. 2015, 22, 199–205. [Google Scholar] [CrossRef]
- Badenhorst, C.E.; Dawson, B.; Goodman, C.; Sim, M.; Cox, G.R.; Gore, C.J.; Tjalsma, H.; Swinkels, D.W.; Peeling, P. Influence of post-exercise hypoxic exposure on hepcidin response in athletes. Eur. J. Appl. Physiol. 2014, 114, 951–959. [Google Scholar] [CrossRef]
- Govus, A.D.; Abbiss, C.R.; Garvican-Lewis, L.A.; Swinkels, D.W.; Laarakkers, C.M.; Gore, C.J.; Peeling, P. Acute hypoxic exercise does not alter post-exercise iron metabolism in moderately trained endurance athletes. Eur. J. Appl. Physiol. 2014, 114, 2183–2191. [Google Scholar] [CrossRef]
- Govus, A.D.; Peeling, P.; Abbiss, C.R.; Lawler, N.G.; Swinkels, D.W.; Laarakkers, C.M.; Thompson, K.G.; Peiffer, J.J.; Gore, C.J.; Garvican-Lewis, L.A. Live high, train low—Influence on resting and post-exercise hepcidin levels. Scand. J. Med. Sci. Sports 2017, 27, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Pigeon, C.; Ilyin, G.; Courselaud, B.; Leroyer, P.; Turlin, B.; Brissot, P.; Loréal, O. A new mouse liver-specific gene, encoding a protein homologous to human antimicrobial peptide hepcidin, is overexpressed during iron overload. J. Biol. Chem. 2001, 276, 7811–7819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, R.; Jung, C.-L.; Gabayan, V.; Deng, J.C.; Ganz, T.; Nemeth, E.; Bulut, Y. Hepcidin induction by pathogens and pathogen-derived molecules is strongly dependent on interleukin-6. Infect. Immun. 2014, 82, 745–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayeur, C.; Leyton, P.A.; Kolodziej, S.A.; Yu, B.; Bloch, K.D. BMP type II receptors have redundant roles in the regulation of hepatic hepcidin gene expression and iron metabolism. Blood 2014, 124, 2116–2123. [Google Scholar] [CrossRef] [Green Version]
- Kasprowicz, K.; Ziemann, E.; Ratkowski, W.; Laskowski, R.; Kaczor, J.J.; Dadci, R.; Antosiewicz, J. Running a 100-km ultra-marathon induces an inflammatory response but does not raise the level of the plasma iron-regulatory protein hepcidin. J. Sports Med. Phys. Fitness 2013, 53, 533–537. [Google Scholar]
- Chiu, Y.-H.; Lai, J.-I.; Wang, S.-H.; How, C.-K.; Li, L.-H.; Kao, W.-F.; Yang, C.-C.; Chen, R.-J. Early changes of the anemia phenomenon in male 100-km ultramarathoners. J. Chin. Med. Assoc. 2015, 78, 108–113. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, R.; Sánchez-Oliver, A.J.; Mata-Ordoñez, F.; Feria-Madueño, A.; Grimaldi-Puyana, M.; López-Samanes, Á.; Pérez-López, A. Effects of an Acute Exercise Bout on Serum Hepcidin Levels. Nutrients 2018, 10, 209. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.M.; Alvarez, J.A.; Kearns, M.D.; Hao, L.; Sloan, J.H.; Konrad, R.J.; Ziegler, T.R.; Zughaier, S.M.; Tangpricha, V. High-dose vitamin D3 reduces circulating hepcidin concentrations: A pilot, randomized, double-blind, placebo-controlled trial in healthy adults. Clin. Nutr. 2017, 36, 980–985. [Google Scholar] [CrossRef]
- Alon, D.B.; Chaimovitz, C.; Dvilansky, A.; Lugassy, G.; Douvdevani, A.; Shany, S.; Nathan, I. Novel role of 1,25(OH)(2)D(3) in induction of erythroid progenitor cell proliferation. Exp. Hematol. 2002, 30, 403–409. [Google Scholar] [CrossRef]
- Thomas, C.E.; Guillet, R.; Queenan, R.A.; Cooper, E.M.; Kent, T.R.; Pressman, E.K.; Vermeylen, F.M.; Roberson, M.S.; O’Brien, K.O. Vitamin D status is inversely associated with anemia and serum erythropoietin during pregnancy. Am. J. Clin. Nutr. 2015, 102, 1088–1095. [Google Scholar] [CrossRef] [Green Version]
- Nikooyeh, B.; Neyestani, T.R. Poor vitamin D status increases the risk of anemia in school children: National Food and Nutrition Surveillance. Nutrition 2018, 47, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Malczewska-Lenczowska, J.; Sitkowski, D.; Surała, O.; Orysiak, J.; Szczepańska, B.; Witek, K. The Association between Iron and Vitamin D Status in Female Elite Athletes. Nutrients 2018, 10, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacchetta, J.; Zaritsky, J.J.; Sea, J.L.; Chun, R.F.; Lisse, T.S.; Zavala, K.; Nayak, A.; Wesseling-Perry, K.; Westerman, M.; Hollis, B.W.; et al. Suppression of iron-regulatory hepcidin by vitamin D. J. Am. Soc. Nephrol. 2014, 25, 564–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knechtle, B.; Nikolaidis, P.T. Physiology and Pathophysiology in Ultra-Marathon Running. Front. Physiol. 2018, 9, 634. [Google Scholar] [CrossRef] [Green Version]
- Wołyniec, W.; Ratkowski, W.; Kasprowicz, K.; Jastrzębski, Z.; Małgorzewicz, S.; Witek, K.; Grzywacz, T.; Żmijewski, P.; Renke, M. Glomerular Filtration Rate Is Unchanged by Ultramarathon. J. Strength Cond. Res. 2018, 32, 3207–3215. [Google Scholar] [CrossRef]
- Bruce, R.A. Exercise testing of patients with coronary heart disease. Principles and normal standards for evaluation. Ann. Clin. Res. 1971, 3, 323–332. [Google Scholar]
- Hopkins, W.G. Individual responses made easy. J. Appl. Physiol. 2015, 118, 1444–1446. [Google Scholar] [CrossRef] [Green Version]
- Agergaard, J.; Trøstrup, J.; Uth, J.; Iversen, J.V.; Boesen, A.; Andersen, J.L.; Schjerling, P.; Langberg, H. Does vitamin-D intake during resistance training improve the skeletal muscle hypertrophic and strength response in young and elderly men?—A randomized controlled trial. Nutr. Metab. 2015, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Ogan, D.; Pritchett, K. Vitamin D and the athlete: Risks, recommendations, and benefits. Nutrients 2013, 5, 1856–1868. [Google Scholar] [CrossRef] [Green Version]
- He, C.-S.; Fraser, W.D.; Tang, J.; Brown, K.; Renwick, S.; Rudland-Thomas, J.; Teah, J.; Tanqueray, E.; Gleeson, M. The effect of 14 weeks of vitamin D3 supplementation on antimicrobial peptides and proteins in athletes. J. Sports Sci. 2016, 34, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Owens, D.J.; Fraser, W.D.; Close, G.L. Vitamin D and the athlete: Emerging insights. Eur. J. Sport Sci. 2015, 15, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Wyon, M.A.; Wolman, R.; Nevill, A.M.; Cloak, R.; Metsios, G.S.; Gould, D.; Ingham, A.; Koutedakis, Y. Acute Effects of Vitamin D3 Supplementation on Muscle Strength in Judoka Athletes: A Randomized Placebo-Controlled, Double-Blind Trial. Clin. J. Sport Med. 2016, 26, 279–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carswell, A.T.; Oliver, S.J.; Wentz, L.M.; Kashi, D.S.; Roberts, R.; Tang, J.C.Y.; Izard, R.M.; Jackson, S.; Allan, D.; Rhodes, L.E.; et al. Influence of Vitamin D Supplementation by Sunlight or Oral D3 on Exercise Performance. Med. Sci. Sports Exerc. 2018, 50, 2555–2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jastrzębska, M.; Kaczmarczyk, M.; Michalczyk, M.; Radzimiński, Ł.; Stępień, P.; Jastrzębska, J.; Wakuluk, D.; Suárez, A.D.; López Sánchez, G.F.; Cięszczyk, P.; et al. Can Supplementation of Vitamin D Improve Aerobic Capacity in Well Trained Youth Soccer Players? J. Hum. Kinet. 2018, 61, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Książek, A.; Dziubek, W.; Pietraszewska, J.; Słowińska-Lisowska, M. Relationship between 25(OH)D levels and athletic performance in elite Polish judoists. Biol. Sport 2018, 35, 191–196. [Google Scholar] [CrossRef]
- Skalska, M.; Nikolaidis, P.T.; Knechtle, B.; Rosemann, T.J.; Radzimiński, Ł.; Jastrzębska, J.; Kaczmarczyk, M.; Myśliwiec, A.; Dragos, P.; López-Sánchez, G.F.; et al. Vitamin D Supplementation and Physical Activity of Young Soccer Players during High-Intensity Training. Nutrients 2019, 11, 349. [Google Scholar] [CrossRef] [Green Version]
- Owens, D.J.; Tang, J.C.Y.; Bradley, W.J.; Sparks, A.S.; Fraser, W.D.; Morton, J.P.; Close, G.L. Efficacy of High-Dose Vitamin D Supplements for Elite Athletes. Med. Sci. Sports Exerc. 2017, 49, 349–356. [Google Scholar] [CrossRef]
- De Francesco Daher, E.; Mesquita Martiniano, L.V.; Lopes Lima, L.L.; Viana Leite Filho, N.C.; de Oliveira Souza, L.E.; Duarte Fernandes, P.H.; da Silva, S.L.; da Silva Junior, G.B. Acute kidney injury due to excessive and prolonged intramuscular injection of veterinary supplements containing vitamins A, D and E: A series of 16 cases. Nefrologia 2017, 37, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Burt, L.A.; Billington, E.O.; Rose, M.S.; Raymond, D.A.; Hanley, D.A.; Boyd, S.K. Effect of High-Dose Vitamin D Supplementation on Volumetric Bone Density and Bone Strength: A Randomized Clinical Trial. JAMA 2019, 322, 736–745. [Google Scholar] [CrossRef]
- Owens, D.J.; Allison, R.; Close, G.L. Vitamin D and the Athlete: Current Perspectives and New Challenges. Sports Med. 2018, 48, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Jastrzębska, M.; Kaczmarczyk, M.; Jastrzębski, Z. Effect of Vitamin D Supplementation on Training Adaptation in Well-Trained Soccer Players. J. Strength Cond. Res. 2016, 30, 2648–2655. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, M.E.; Tran, H.T.; Evans, M.V. A physiologically based pharmacokinetic model of vitamin D. J. Appl. Toxicol. 2017, 12, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Schoenmakers, I.; Gousias, P.; Jones, K.S.; Prentice, A. Prediction of winter vitamin D status and requirements in the UK population based on 25(OH) vitamin D half-life and dietary intake data. J. Steroid. Biochem. Mol. Biol. 2016, 164, 218–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braithwaite, V.S.; Crozier, S.R.; D’Angelo, S.; Prentice, A.; Cooper, C.; Harvey, N.C.; Jones, K.S. The Effect of Vitamin D Supplementation on Hepcidin, Iron Status, and Inflammation in Pregnant Women in the United Kingdom. Nutrients 2019, 11, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Cruz-Góngora, V.; Salinas-Rodríguez, A.; Villalpando, S.; Flores-Aldana, M. Serum Retinol but Not 25(OH)D Status Is Associated with Serum Hepcidin Levels in Older Mexican Adults. Nutrients 2019, 11, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M. Evaluation, treatment, and prevention of vitamin D deficiency: An Endocrine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roecker, L.; Meier-Buttermilch, R.; Brechtel, L.; Nemeth, E.; Ganz, T. Iron-regulatory protein hepcidin is increased in female athletes after a marathon. Eur. J. Appl. Physiol. 2005, 95, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Adegoke, S.A.; Smith, O.S.; Adekile, A.D.; Figueiredo, M.S. Relationship between serum 25-hydroxyvitamin D and inflammatory cytokines in paediatric sickle cell disease. Cytokine 2017, 96, 87–93. [Google Scholar] [CrossRef]
- Parsaie, N.; Ghavamzadeh, S.; Cheraghi, M. Effects of cholecalciferol supplementation on inflammatory markers and muscle damage indices of soccer players after a simulated soccer match. Nutrition 2019, 59, 37–43. [Google Scholar] [CrossRef]
- Pedersen, B.K. Anti-inflammatory effects of exercise: Role in diabetes and cardiovascular disease. Eur. J. Clin. Investig. 2017, 47, 600–611. [Google Scholar] [CrossRef] [Green Version]
- Dahlquist, D.T.; Stellingwerff, T.; Dieter, B.P.; McKenzie, D.C.; Koehle, M.S. Effects of macro- and micronutrients on exercise-induced hepcidin response in highly trained endurance athletes. Appl. Physiol. Nutr. Metab. 2017, 42, 1036–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badenhorst, C.E.; Dawson, B.; Cox, G.R.; Laarakkers, C.M.; Swinkels, D.W.; Peeling, P. Timing of post-exercise carbohydrate ingestion: Influence on IL-6 and hepcidin responses. Eur. J. Appl. Physiol. 2015, 115, 2215–2222. [Google Scholar] [CrossRef] [PubMed]
- McKay, A.K.A.; Peeling, P.; Pyne, D.B.; Welvaert, M.; Tee, N.; Leckey, J.J.; Sharma, A.P.; Ross, M.L.R.; Garvican-Lewis, L.A.; van Swelm, R.P.L.; et al. Acute carbohydrate ingestion does not influence the post-exercise iron-regulatory response in elite keto-adapted race walkers. J. Sci. Med. Sport 2019, 22, 635–640. [Google Scholar] [CrossRef] [PubMed]
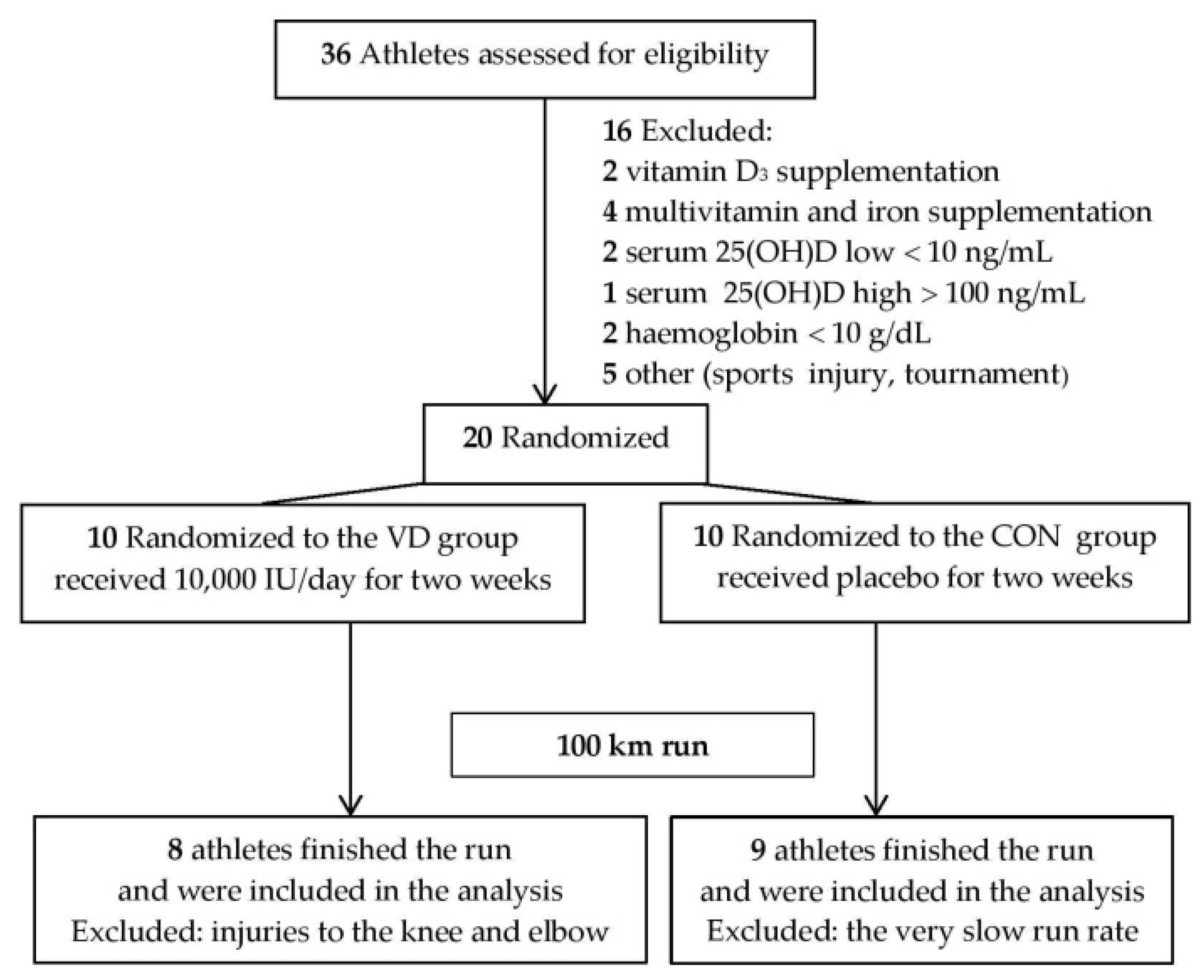

{kind=link}
| Variable | Vitamin D Group | Control Group | p |
|---|---|---|---|
| Number of subjects | 10 | 10 | |
| Age (years) | 39.0 ± 5.93 | 42.5 ± 8.37 | 0.143 |
| Vitamin D3 supplementation | 10,000 IU/14 days | 0 | |
| Finishers | 8 | 9 | |
| Age (years) | 39.56 ± 6.0 | 40.87 ± 2.36 | 0.281 |
| 25(OH)D—before supplementation (ng/mL) | 27.26 ± 7.09 | 27.13 ± 3.67 | 0.518 |
| BMI (kg/m2) | 24.31 ± 2.38 | 24.21 ± 2.51 | 0.533 |
| Height (cm) | 177.59 ± 6.12 | 178.59 ± 5.49 | 0.362 |
| Weight (kg) | 76.68 ± 7.93 | 76.80 ± 8.21 | 0.488 |
| VO2 max (mL × kg−1 × min−1) | 56.65 ± 4.88 | 53.30 ± 7.34 | 0.865 |
| Variable | Mean ± SD (pre) | pa | Mean ± SD (post) | pa | Mean Change ± SD | pa | Difference in Mean Change (Standardized) | Individual Response as SD | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CON (n = 9) | VD (n = 8) | CON (n = 9) | VD (n = 8) | CON (n = 9) | VD (n = 8) | ||||||
| Iron (µg/dL) | 112.78 ± 50.13 | 94.14 ± 23.98 | 0.828 | 35.78 ± 11.70 | 66.86 ± 36.24 | 0.028 | −77.00 ± 48.68 | −27.29 ± 39.57 | 0.041 | 49.7; 2.4; 97.0 (1.26; 0.06; 2.45) | −28.3 (−61.0; 46.0) |
| Hepcidin (ng/mL) | 0.54 ± 0.28 | 0.63 ± 0.22 | 0.232 | 0.72 ± 0.33 | 1.41 ± 1.02 | 0.056 | 0.18 ± 0.47 | 0.77 ± 0.92 | 0.196 | 0.59; −0.39; 1.58 (−0.88; −0.57; −2.34) | 0.79 (−0.67; 1.31) |
| IL6 (pg/mL) | 1.04 ± 0.35 | 0.87 ± 0.31 | 0.849 | 23.0 ± 1.40 | 21.83 ± 2.68 | 0.851 | 21.96 ± 1.56 | 20.96 ± 2.65 | 0.409 | −1.0; −3.6; 1.6 (−3.02; −10.91; 4.87) | 2.1 (−2.0; 3.6) |
| Ferritin (ng/mL) | 61.89 ± 34.61 | 35.43 ± 25.34 | 0.949 | 81.11 ± 43.86 | 47.00 ± 28.68 | 0.959 | 19.22 ± 18.48 | 11.57 ± 9.41 | 0.303 | −7.7; −23.1; 7.8 (−0.24; −0.73; 0.25) | −15.9 (−24.5; 9.8) |
| UIBC (µg/dL) | 199.75 ± 46.56 | 242.14 ± 60.85 | 0.069 | 274.75 ± 36.41 | 287.14 ± 71.95 | 0.334 | 75.00 ± 42.15 | 45.00 ± 40.27 | 0.183 | −30.0; −76.2; 16.2 (−0.55; −1.40; 0.30) | −12.4 (−52.6; 49.6) |
| TIBC (µg/dL) | 312.13 ± 45.98 | 336.29 ± 64.97 | 0.201 | 309.13 ± 34.14 | 354.00 ± 58.59 | 0.051 | −3.00 ± 29.73 | 17.71 ± 36.42 | 0.256 | 20.7, −17.3; 58.7 (0.39, −0.32; 1.09) | 21.0 (−36.3; 47.0) |
| sTfR (µg/mL) | 3.17 ± 0.75 | 3.19 ± 0.98 | 0.482 | 3.38 ± 1.03 | 3.07 ± 0.89 | 0.737 | 0.21 ± 0.38 | −0.11 ± 0.41 | 0.127 | −0.3; −0.8; 0.1 (−0.40; −0.94; 0.13) | 0.14 (−0.47; 0.50) |
| Variable | Mean ± SD (post) | pa | Mean ± SD (post12) | pa | Mean Change ± SD | pa | Difference in Mean Change (Standardized) | Individual Response as SD | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CON (n = 9) | VD (n = 8) | CON (n = 9) | VD (n = 8) | CON (n = 9) | VD (n = 8) | ||||||
| Iron (µg/dL) | 37.75 ± 10.79 | 66.86 ± 36.24 | 0.033 | 197.13 ± 69.07 | 184.29 ± 76.57 | 0.641 | 159.38 ± 68.62 | 117.43 ± 48.12 | 0.191 | −41.9; −107.9; 24.0 (−1.44; −3.70; 0.82) | −48.9 (−89.3; 56.5) |
| Hepcidin (ng/mL) | 0.77 ± 0.33 | 1.41 ± 1.02 | 0.068 | 0.71 ± 0.19 | 0.78 ± 0.37 | 0.320 | −0.06 ± 0.36 | −0.63±0.98 | 0.229 | −0.6; −1.6; 0.5 (−0.72; −2.05; 0.60) | 0.91 (−0.61;1.43) |
| IL6 (pg/mL) | 23.00 ± 1.40 | 21.83 ± 2.68 | 0.851 | 11.49 ± 6.83 | 7.09 ± 4.66 | 0.931 | −11.50 ± 7.82 | −14.73±5.37 | 0.388 | −3.2; −11.2; 4.7 (−1.51; −5.22; 2.21) | −5.7 (−10.4; 6.6) |
| Ferritin (ng/mL) | 72.75 ± 38.46 | 47.00 ± 28.68 | 0.924 | 105.63 ± 43.02 | 71.86 ± 33.38 | 0.958 | 32.88 ± 18.97 | 24.86 ± 9.99 | 0.320 | −8.0; −25.1; 9.1 (−0.20; −0.62; 0.22) | −16.1 (−25.6; 11.6) |
| UIBC (µg/dL) | 286.86 ± 32.43 | 287.14 ± 71.95 | 0.496 | 129.29 ± 43.87 | 133.86 ± 78.14 | 0.444 | −157.57 ± 45.21 | −153.29 ± 62.57 | 0.886 | 4.3; −60.6; 69.2 (0.08; −1.15; 1.32) | 43.3 (−55.9; 82.9) |
| TIBC (µg/dL) | 325.57 ± 23.85 | 354 ± 58.59 | 0.109 | 309.43 ± 26.01 | 318.14 ± 52.63 | 0.339 | −16.14 ± 26.81 | −35.86 ± 39.84 | 0.302 | −19.7; −60.1; 20.6 (−0.40; −1.22; 0.42) | 29.5 (−33.2; 53.3) |
| sTfR (µg/mL) | 3.49 ± 1.04 | 3.07 ± 0.89 | 0.803 | 2.93 ± 0.80 | 2.74 ± 0.49 | 0.714 | −0.56 ± 0.34 | −0.33 ± 0.52 | 0.334 | 0.2; −0.3; 0.7 (0.25; −0.29; 0.79) | 0.40 (−0.42; 0.70) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasprowicz, K.; Ratkowski, W.; Wołyniec, W.; Kaczmarczyk, M.; Witek, K.; Żmijewski, P.; Renke, M.; Jastrzębski, Z.; Rosemann, T.; Nikolaidis, P.T.; et al. The Effect of Vitamin D3 Supplementation on Hepcidin, Iron, and IL-6 Responses after a 100 km Ultra-Marathon. Int. J. Environ. Res. Public Health 2020, 17, 2962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17082962
Kasprowicz K, Ratkowski W, Wołyniec W, Kaczmarczyk M, Witek K, Żmijewski P, Renke M, Jastrzębski Z, Rosemann T, Nikolaidis PT, et al. The Effect of Vitamin D3 Supplementation on Hepcidin, Iron, and IL-6 Responses after a 100 km Ultra-Marathon. International Journal of Environmental Research and Public Health. 2020; 17(8):2962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17082962
Chicago/Turabian StyleKasprowicz, Katarzyna, Wojciech Ratkowski, Wojciech Wołyniec, Mariusz Kaczmarczyk, Konrad Witek, Piotr Żmijewski, Marcin Renke, Zbigniew Jastrzębski, Thomas Rosemann, Pantelis T. Nikolaidis, and et al. 2020. "The Effect of Vitamin D3 Supplementation on Hepcidin, Iron, and IL-6 Responses after a 100 km Ultra-Marathon" International Journal of Environmental Research and Public Health 17, no. 8: 2962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17082962