
Development of Genus-Specific PCR Primers for Molecular Monitoring of Invasive Nostocalean Cyanobacteria

Abstract
:1. Introduction
2. Materials and Methods
2.1. Target Cyanobacteria
2.2. Sampling and Phytoplankton Analysis
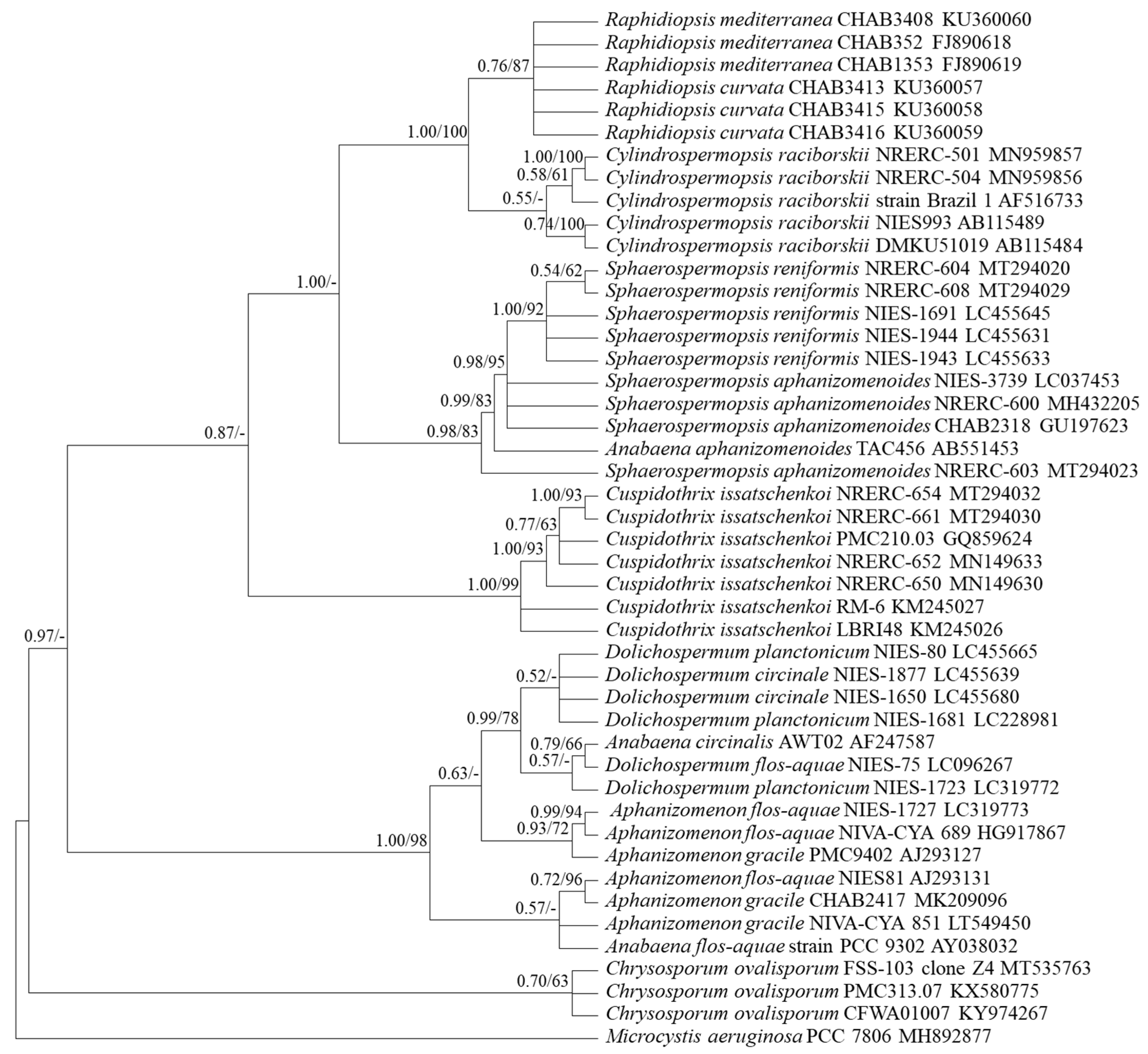
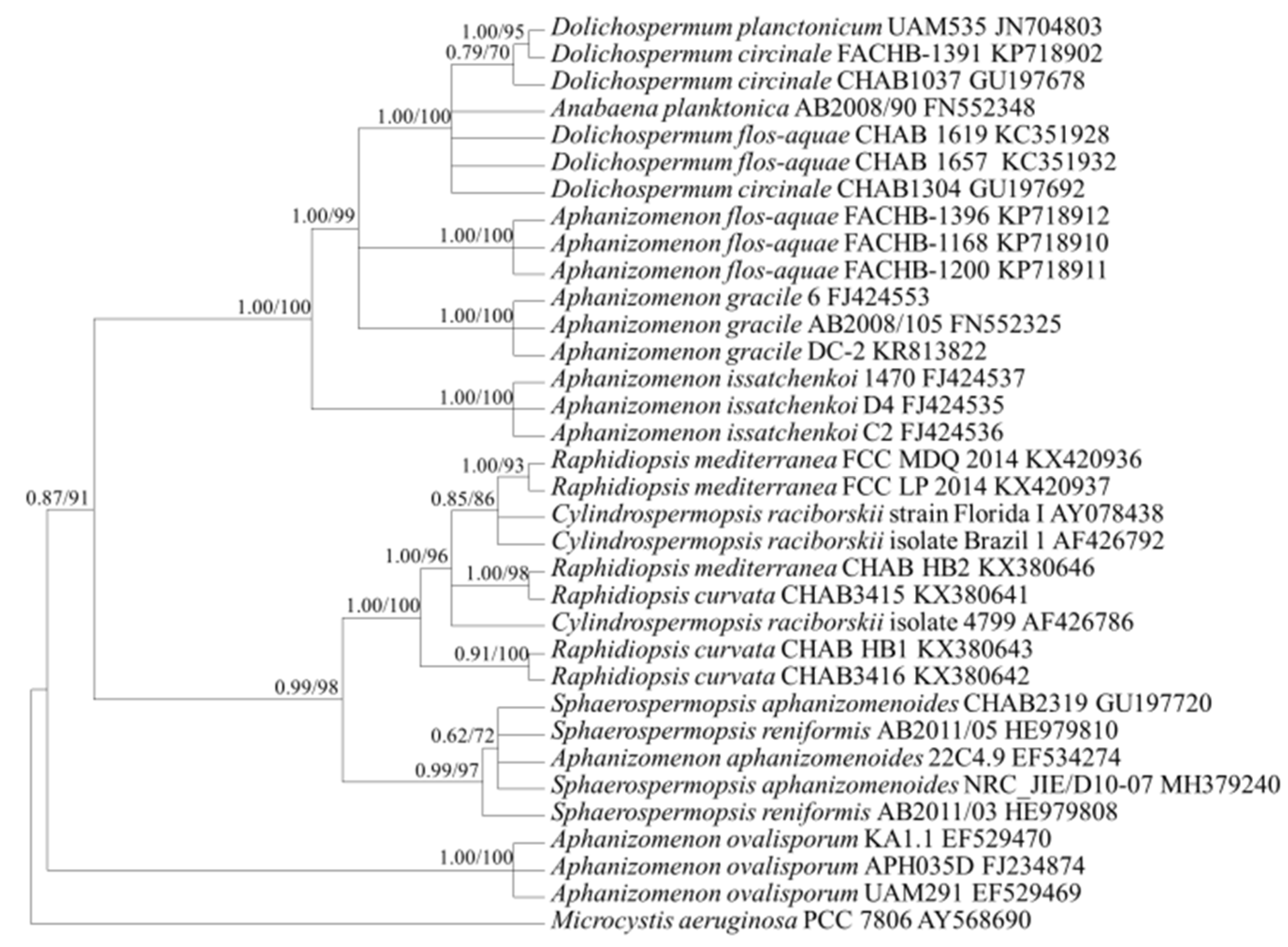
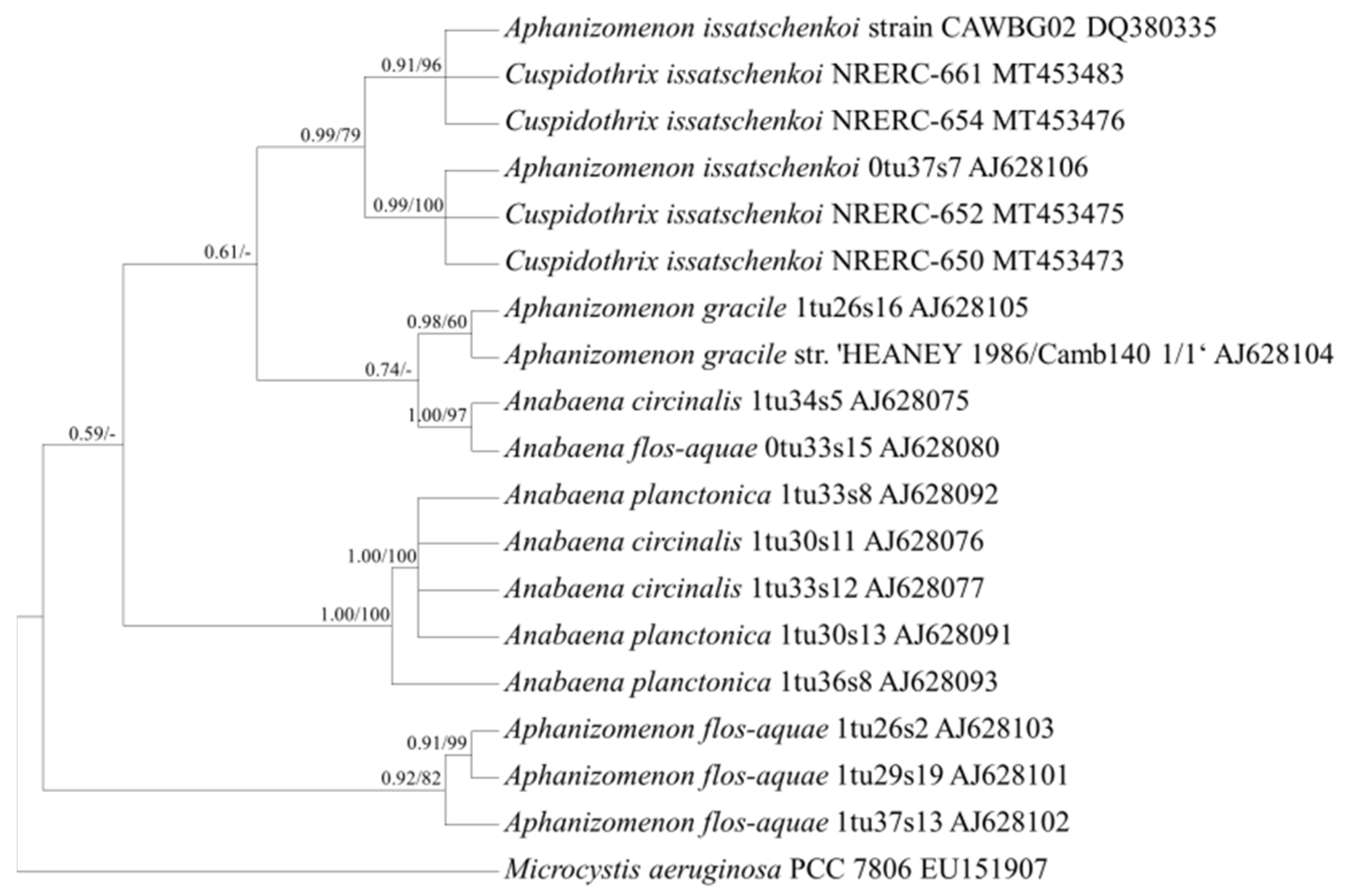
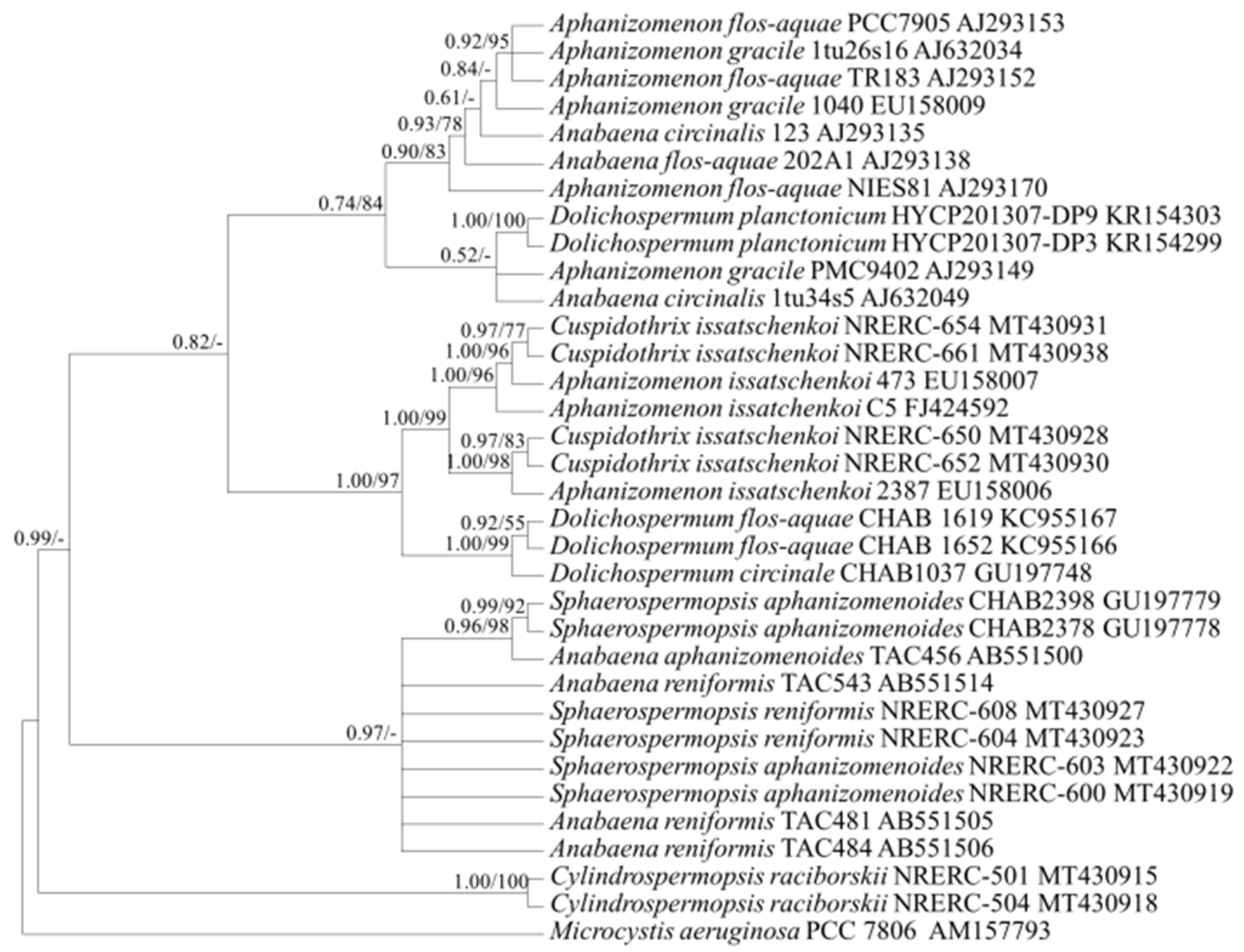
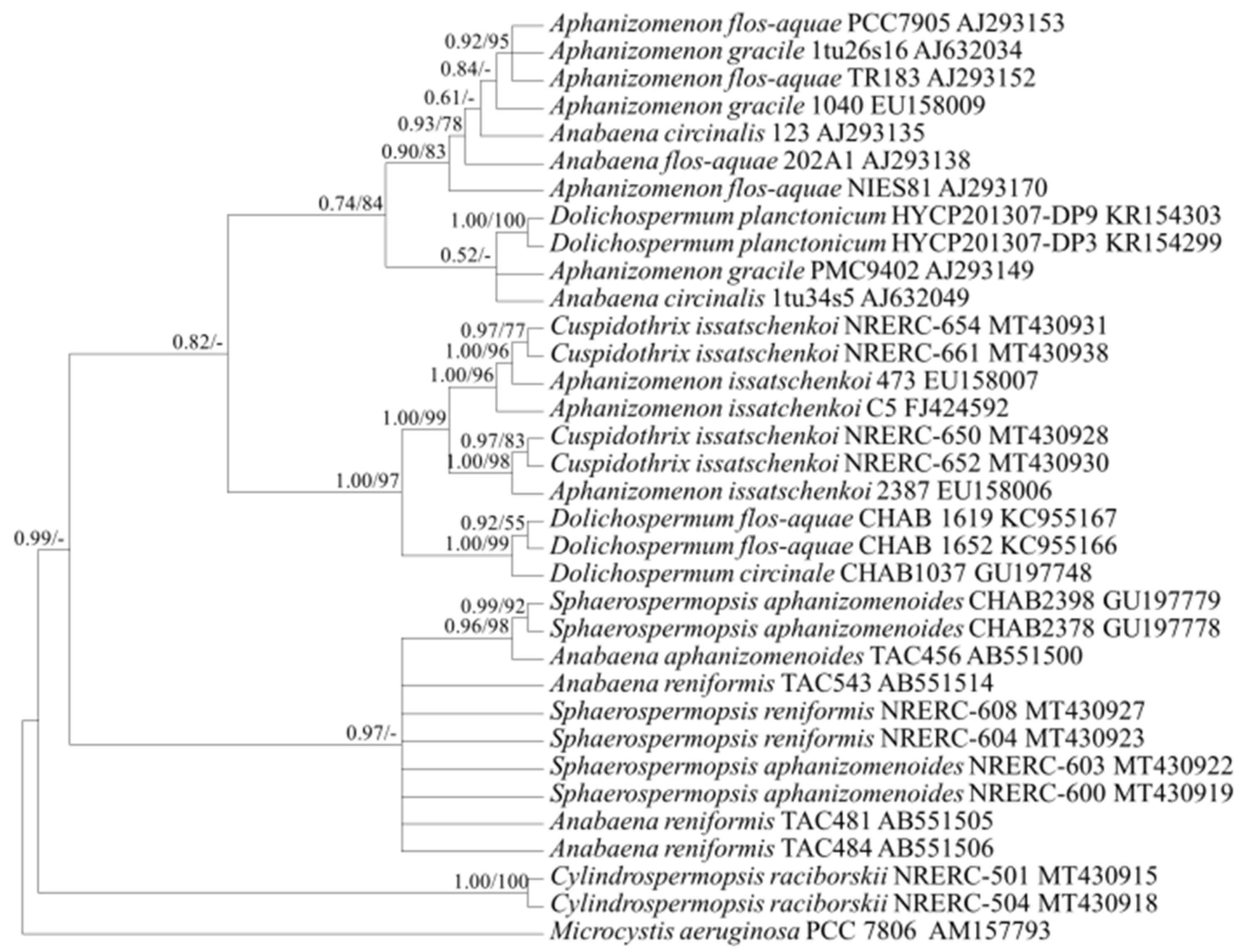
2.3. Target Gene Sequences and Phylogenetic Analysis
2.3.1. DNA Extraction
2.3.2. Gene Sequence Characteristics and Phylogenetic Analysis
2.4. Design of Genus-Specific Primers and Determination of PCR Amplification Conditions
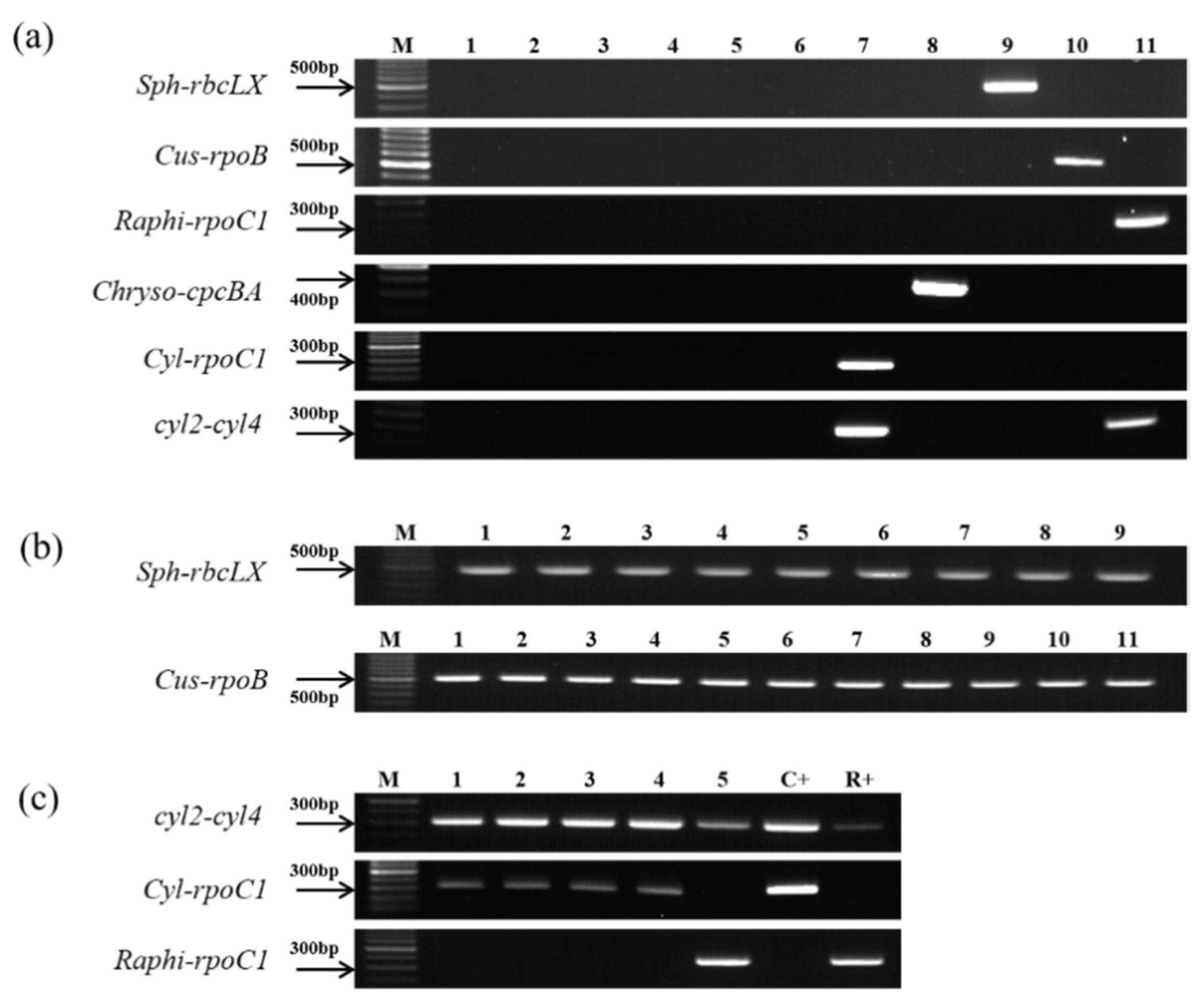
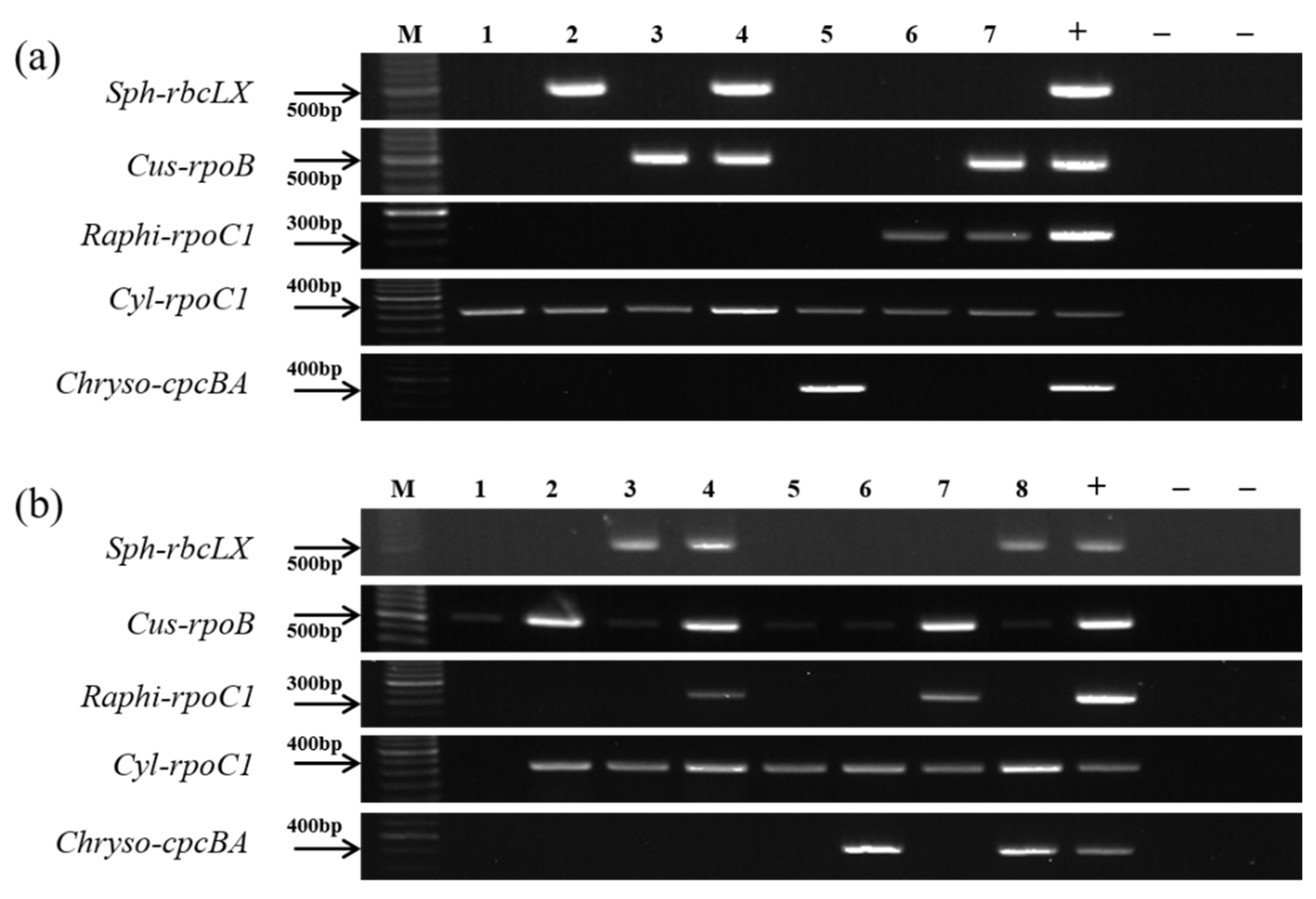
2.5. Verification of the Genus-Specific Primers
3. Results
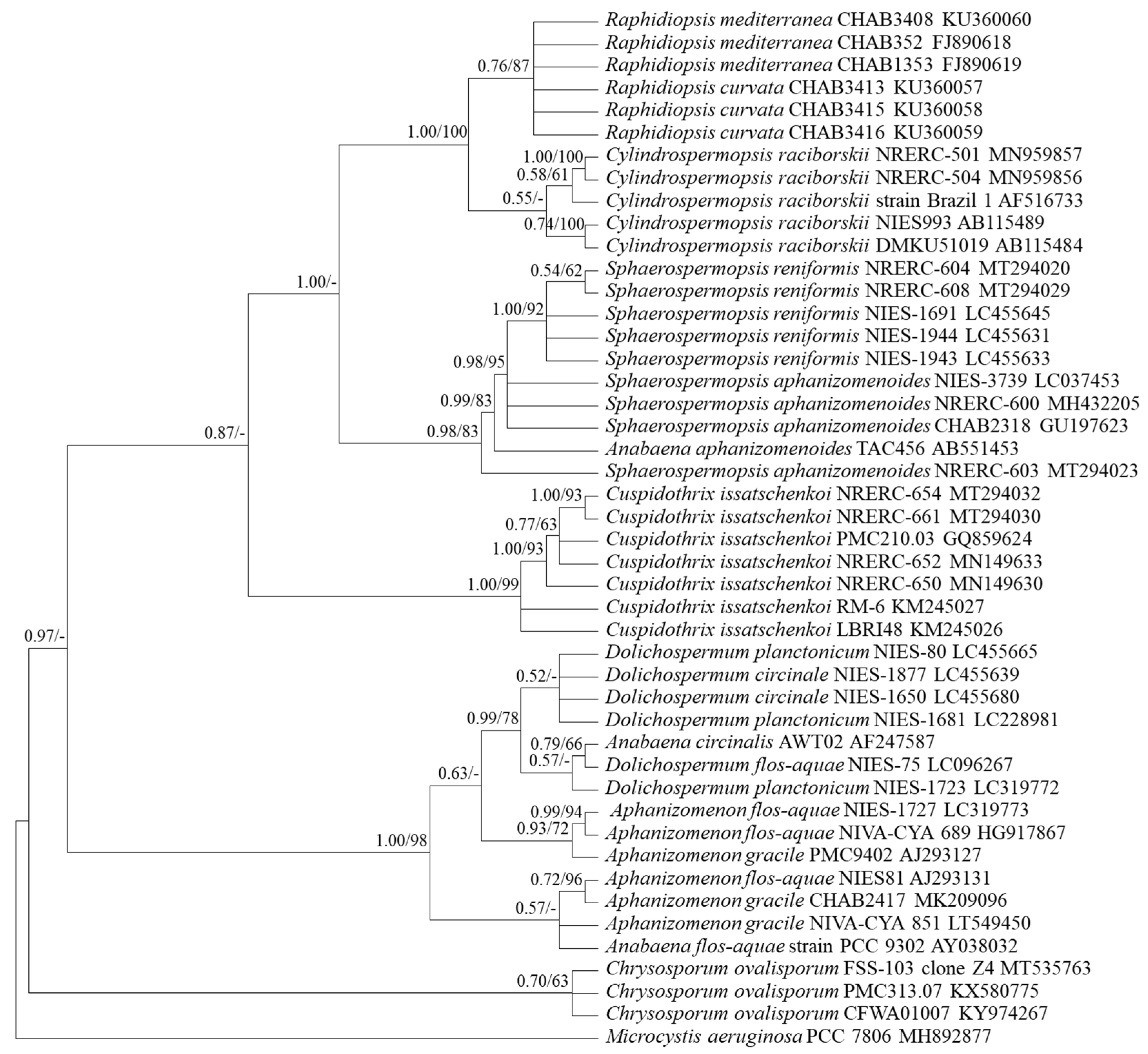
3.1. Selection of Genetic Markers through Phylogenetic Analyses
3.2. Design of Genus-Specific PCR Primers
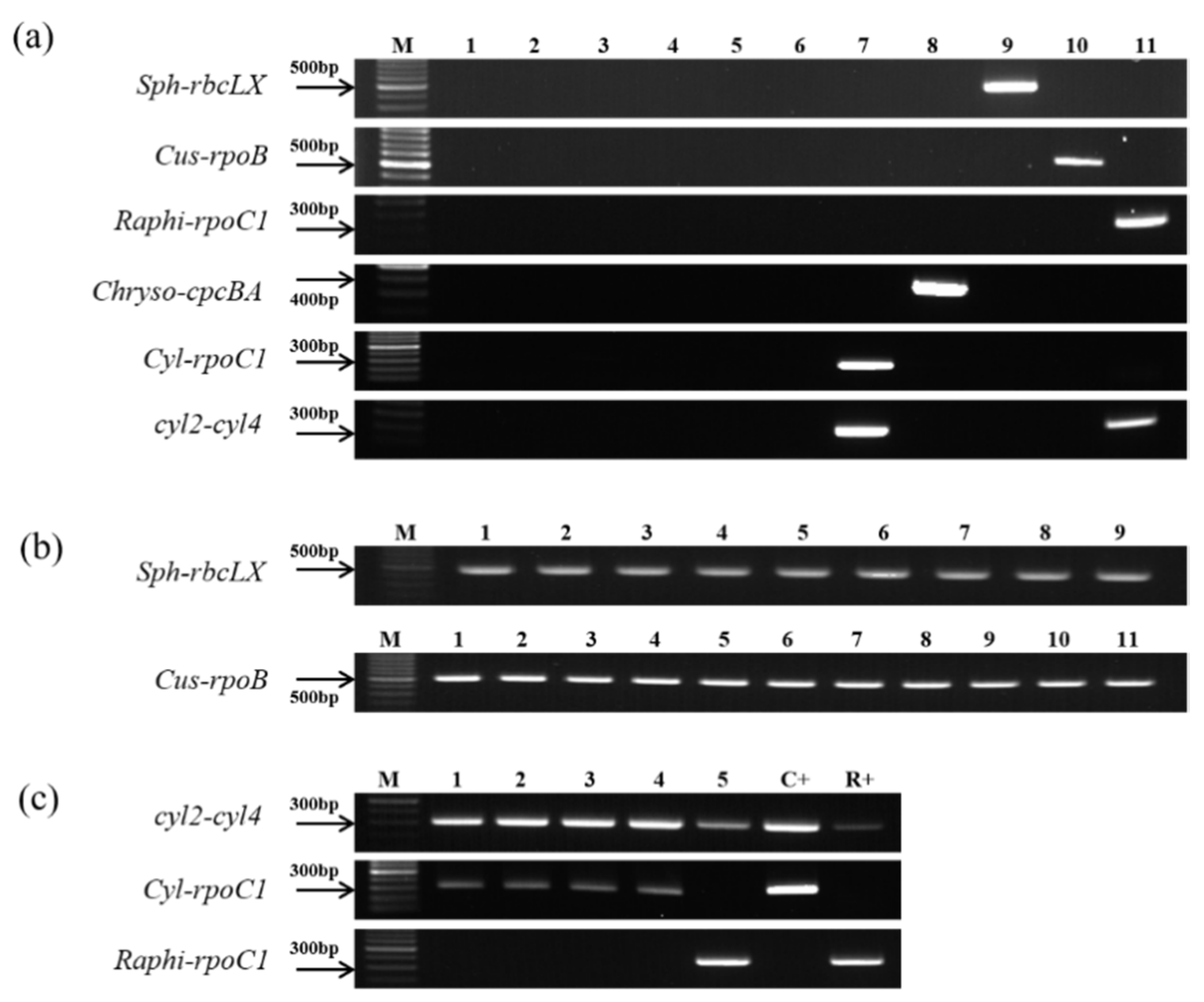
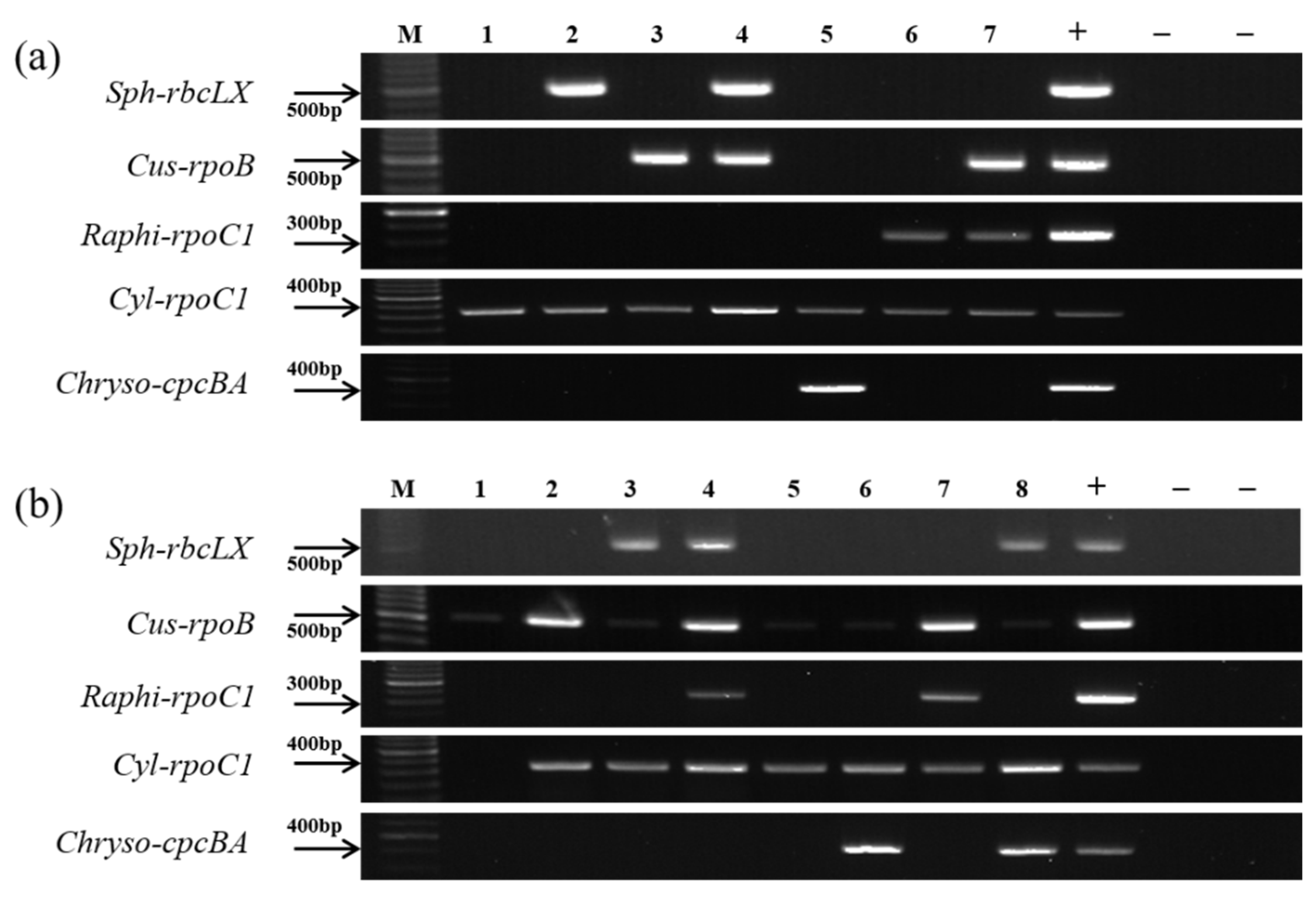
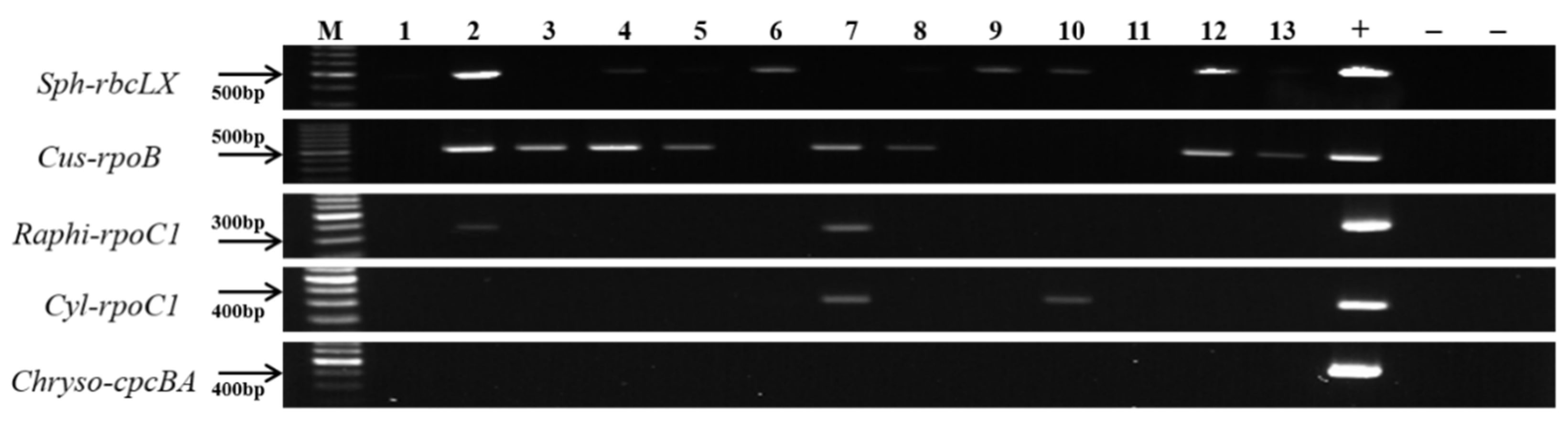
3.3. Verification of the Genus-Specific Primers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kaštovský, J.; Hauer, T.; Mareš, J.; Krautová, M.; Bešta, T.; Komárek, J.; Desortová, B.; Heteša, J.; Hindáková, A.; Houk, V. A review of the alien and expansive species of freshwater cyanobacteria and algae in the Czech Republic. Biol. Invasions 2010, 12, 3599–3625. [Google Scholar] [CrossRef]
- Sinha, R.; Pearson, L.A.; Davis, T.W.; Burford, M.A.; Orr, P.T.; Neilan, B.A. Increased incidence of Cylindrospermopsis raciborskii in temperate zones–is climate change responsible? Water Res. 2012, 46, 1408–1419. [Google Scholar] [CrossRef]
- Sukenik, A.; Hadas, O.; Kaplan, A.; Quesada, A. Invasion of Nostocales (cyanobacteria) to Subtropical and Temperate Freshwater Lakes—Physiological, Regional, and Global Driving Forces. Front. Microbiol. 2012, 3, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Cirés, S.; Ballot, A. A review of the phylogeny, ecology and toxin production of bloom-forming Aphanizomenon spp. and related species within the Nostocales (cyanobacteria). Harmful Algae 2016, 54, 21–43. [Google Scholar] [CrossRef]
- Stüken, A.; Rücker, J.; Endrulat, T.; Preussel, K.; Hemm, M.; Nixdorf, B.; Karsten, U.; Wiedner, C. Distribution of three alien cyanobacterial species (Nostocales) in northeast Germany: Cylindrospermopsis raciborskii, Anabaena bergii and Aphanizomenon aphanizomenoides. Phycologia 2006, 45, 696–703. [Google Scholar] [CrossRef]
- Budzyńska, A.; Rosińska, J.; Pełechata, A. Environmental factors driving the occurrence of the invasive cyanobacterium Sphaerospermopsis aphanizomenoides (Nostocales) in temperate lakes. Sci. Total Environ. 2019, 650, 1338–1347. [Google Scholar] [CrossRef]
- Kokociński, M.; Soininen, J. New insights into the distribution of alien cyanobacterium Chrysosporum bergii (Nostocales, Cyanobacteria). Phycological. Res. 2019, 67, 208–214. [Google Scholar] [CrossRef]
- Dahllöf, I.; Baillie, H.; Kjelleberg, S. rpoB-based microbial community analysis avoids limitations inherent in 16S rRNA gene intraspecies heterogeneity. Appl. Environ. Microbiol. 2000, 66, 3376–3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gugger, M.; Lyra, C.; Henriksen, P.; Coute, A.; Humbert, J.-F.; Sivonen, K. Phylogenetic comparison of the cyanobacterial genera Anabaena and Aphanizomenon. Int. J. Syst. Evol. Microbiol. 2002, 52, 1867–1880. [Google Scholar]
- Richert, K.; Brambilla, E.; Stackebrandt, E. The phylogenetic significance of peptidoglycan types: Molecular analysis of the genera Microbacterium and Aureobacterium based upon sequence comparison of gyrB, rpoB, recA and ppk and 16SrRNA genes. Syst. Appl. Microbiol. 2007, 30, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Tamas, I.; Svircev, Z.; Andersson, S.G. Determinative value of a portion of the nifH sequence for the genera Nostoc and Anabaena (Cyanobacteria). Curr. Microbiol. 2000, 41, 197–200. [Google Scholar] [CrossRef]
- Rajaniemi, P.; Hrouzek, P.; Kaštovska, K.; Willame, R.; Rantala, A.; Hoffmann, L.; Komárek, J.; Sivonen, K. Phylogenetic and morphological evaluation of the genera Anabaena, Aphanizomenon, Trichormus and Nostoc (Nostocales, Cyanobacteria). Int. J. Syst. Evol. Microbiol. 2005, 55, 11–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Shi, J.; Lin, S.; Li, R. Unraveling molecular diversity and phylogeny of Aphanizomenon (nostocales, cyanobacteria) strains isolated from china. J. Phycol. 2010, 46, 1048–1058. [Google Scholar] [CrossRef]
- Katrianna, H.; Fewer, D.P.; Fewer, L.M.; Lyra, C.; Eronen, E.; Sivonen, K. Genetic diversity in strains of the genus Anabaena isolated from planktonic and benthic habitats of the Gulf of Finland (Baltic Sea). FEMS Microbiol. Ecol. 2008, 64, 199–208. [Google Scholar]
- Tomitani, A.; Knoll, A.H.; Cavanaugh, C.M.; Ohno, T. The evolutionary diversification of cyanobacteria: Molecular–phylogenetic and paleontological perspectives. Proc. Natl. Acad. Sci. USA 2006, 103, 5442–5447. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J.; Mareš, J. An update to modern taxonomy (2011) of freshwater planktic heterocytous cyanobacteria. Hydrobiologia 2012, 698, 327–351. [Google Scholar] [CrossRef]
- Aguilera, A.; Gómez, E.B.; Kaštovský, J.; Echenique, R.O.; Salerno, G.L. The polyphasic analysis of two native Raphidiopsis isolates supports the unification of the genera Raphidiopsis and Cylindrospermopsis (Nostocales, Cyanobacteria). Phycologia 2018, 57, 130–146. [Google Scholar] [CrossRef]
- Kim, Y.; Park, H.-K.; Kim, I.-S. Invasion and toxin production by exotic nostocalean cyanobacteria (Cuspidothrix, Cylindrospermopsis, and Sphaerospermopsis) in the Nakdong River, Korea. Harmful Algae 2020, 100, 101954. [Google Scholar] [CrossRef] [PubMed]
- Bolch, C.J.S.; Blackburn, S.I. Isolation and purification of Australian isolates of the toxic cyanobacterium Microcystis aeruginosa Kütz. J. Appl. Phycol. 1996, 8, 5–13. [Google Scholar] [CrossRef]
- Park, H.-K.; Lee, H.-J.; Heo, J.; Yun, J.-H.; Kim, Y.-J.; Kim, H.-M.; Hong, D.-G.; Lee, I.-J. Deciphering the key factors determining spatio-temporal heterogeneity of cyanobacterial bloom dynamics in the Nakdong River with consecutive large weirs. Sci. Total Environ. 2021, 755, 143079. [Google Scholar] [CrossRef]
- Komárek, J. Freshwater Flora of Central Europe Vol. 19/3 Cyanoprokaryota, 3rd Part: Heterocytous Genera; Springer: Berlin, Germany, 2013. [Google Scholar]
- John, D.M.; Whitton, B.M.; Brook, A.J. The Freshwater Algal Flora of the British Isles; An Identification Guide to Freshwater and Terrestrial Algae, 2nd ed.; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 2, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, K.M.; Schembri, M.A.; Baker, P.D.; Saint, C.P. Molecular characterization of the toxic cyanobacterium Cylindrospermopsis raciborskii and design of a species-specific PCR. Appl. Environ. Microbiol. 2000, 66, 332–338. [Google Scholar] [CrossRef] [Green Version]
- Stiller, J.W.; McClanahan, A. Phyto-specific 16S rDNA PCR primers for recovering algal and plant sequences from mixed samples. Mol. Ecol. Notes 2005, 5, 1–3. [Google Scholar] [CrossRef]
- Bergsland, K.J.; Haselkorn, R. Evolutionary relationships among eubacteria, cyanobacteria, and chloroplasts: Evidence from the rpoC1 gene of Anabaena sp. strain PCC 7120. J. Bacteriol. 1991, 173, 3446–3455. [Google Scholar] [CrossRef] [Green Version]
- Thomazeau, S.; Houdan-Fourmont, A.; Couté, A.; Duval, C.; Couloux, A.; Rousseau, F.; Bernard, C. The contribution of sub-saharan african strains to the phylogeny of cyanobacteria: Focusing on the nostocaceae (nostocales, cyanobacteria). J. Phycol. 2010, 46, 564–579. [Google Scholar] [CrossRef]
- Fathalli, A.; Jenhani, A.B.R.; Moreira, C.; Welker, M.; Romdhane, M.; Antunes, A.; Vasconcelos, V. Molecular and phylogenetic characterization of potentially toxic cyanobacteria in Tunisian freshwaters. Syst. Appl. Microbiol. 2011, 34, 303–310. [Google Scholar] [CrossRef]
- Fathalli, A.; Jenhani, A.B.R.; Moreira, C.; Azevedo, J.; Welker, M.; Romdhane, M.; Antunes, A.; Vasconcelos, V. Genetic variability of the invasive cyanobacteria Cylindrospermopsis raciborskii from Bir M’cherga reservoir (Tunisia). Arch. Microbiol. 2011, 193, 595–604. [Google Scholar] [CrossRef]
- Robertson, B.R.; Tezuka, N.; Watanabe, M.M. Phylogenetic analyses of Synechococcus strains (cyanobacteria) using sequences of 16S rDNA and part of the phycocyanin operon reveal multiple evolutionary lines and reflect phycobilin content. Int. J. Syst. Evol. Microbiol. 2001, 51, 861–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyble, J.; Paerl, H.W.; Neilan, B.A. Genetic characterization of Cylindrospermopsis raciborskii (cyanobacteria) isolates from diverse geographic origins based on nifH and cpcBA-IGS nucleotide sequence analysis. Appl. Environ. Microbiol. 2002, 68, 2567–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballot, A.; Swe, T.; Mjelde, M.; Cerasino, L.; Hostyeva, V.; Miles, C.O. Cylindrospermopsin- and Deoxycylindrospermopsin-Producing Raphidiopsis raciborskii and Microcystin-Producing Microcystis spp. in Meiktila Lake, Myanmar. Toxins 2020, 12, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lima, D.V.N.; Pacheco, A.B.F.; Goulart, C.L. Physiological responses of Raphidiopsis raciborskii (Cyanobacteria) strains to water conductivity: Effect of sodium and magnesiumions. Hydrobiologia 2020, 847, 2449–2464. [Google Scholar] [CrossRef]
- Sidelev, S.; Koksharova, O.; Babanazarova, O.; Fastner, J.; Chernova, E.; Gusev, E. Phylogeographic, toxicological and ecological evidence for the global distribution of Raphidiopsis raciborskii and its northernmost presence in Lake Nero, Central Western Russia. Harmful Algae 2020, 98, 101889. [Google Scholar] [CrossRef]
- Vico, P.; Bonilla, S.; Cremella, B.; Aubriot, L.; Iriarte, A.; Piccini, C. Biogeography of the cyanobacterium Raphidiopsis (Cylindrospermopsis) raciborskii: Integrating genomics, phylogenetic and toxicity data. Mol. Phylogenet. Evol. 2020, 148, 106824. [Google Scholar] [CrossRef]
- Liu, J.; Huang, S.; Sun, M.; Liu, S.; Liu, Y.; Wang, W.; Zhang, X.; Wang, H.; Hua, W. An improved allele-specific PCR primer design method for SNP marker analysis and its application. Plant Methods 2012, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Cirés, S.; Wörmer, L.; Ballot, A.; Agha, R.; Wiedner, C.; Velázquez, D.; Casero, M.C.; Quesada, A. Phylogeography of cylindrospermopsin and paralytic shellfish toxin-producing Nostocales cyanobacteria from Mediterranean Europe (Spain). Appl. Environ. Microbiol. 2014, 80, 1359–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, M.; McGregor, G.; Eaglesham, G.; Wickramasinghe, W.; Shaw, G. First evidence for the production of cylindrospermopsin and deoxy-cylindrospermopsin by the freshwater benthic cyanobacterium, Lyngbya wollei (Farlow ex Gomont) Speziale and Dyck. Harmful Algae 2007, 6, 73–80. [Google Scholar] [CrossRef]
- Ballot, A.; Fastner, J.; Wiedner, C. Paralytic shellfish poisoning toxin-producing cyanobacterium Aphanizomenon gracile in northeast Germany. Appl. Environ. Microbiol. 2010, 76, 1173–1180. [Google Scholar] [CrossRef] [Green Version]
- Namikoshi, M.; Murakami, T.; Watanabe, M.F.; Oda, T.; Yamada, J.; Tsujimura, S.; Nagai, H.; Oishi, S. Simultaneous production of homoanatoxin-a, ana-toxin-a, and a new non-toxic 4-hydroxyhomoanatoxin-a by the cyanobacterium Raphidiopsis mediterranea Skuja. Toxicon 2003, 42, 533–538. [Google Scholar] [CrossRef]
- Ledreux, A.; Thomazeau, S.; Catherine, A.; Duval, C.; Yéprémian, C.; Marie, A.; Bernard, C. Evidence for saxitoxins production by the cyanobacterium Aphanizomenon gracile in a French recreational water body. Harmful Algae 2010, 10, 88–97. [Google Scholar] [CrossRef]
- Casero, M.C.; Ballot, A.; Agha, R.; Quesada, A.; Cirés, S. Characterization of saxitoxin production and release and phylogeny of sxt genes in paralytic shellfish poisoning toxin-producing Aphanizomenon gracile. Harmful Algae 2014, 37, 28–37. [Google Scholar] [CrossRef]
- Banker, R.; Teltsch, B.; Sukenik, A.; Carmeli, S. 7-Epicylindrospermopsin, a toxic minor metabolite of the cyanobacterium Aphanizomenon ovalisporum from Lake Kinneret, Israel. J. Nat. Prod. 2000, 63, 387–389. [Google Scholar] [CrossRef]
- Moreira, C.; Vasconcelos, V.; Antunes, A. Phylogeny and biogeography of cyanobacteria and their produced toxins. Mar. Drugs 2013, 11, 4350–4369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiripi, L.; Nagy, L.; Kalmár, T.; Kovács, A.; Vörös, L. Insect (Locusta migratoria migratorioides) test monitoring the toxicity of cyanobacteria. Neurotoxicology 1998, 19, 605–608. [Google Scholar] [PubMed]
- Sabour, B.; Loudiki, M.; Oudra, B.; Vasconcelos, V.; Oubraim, S.; Fawzi, B. Contributed article, Dynamics and toxicity of Anabaena aphanizomenoides (Cyanobacteria) water blooms in the shallow brackish Oued Mellah lake (Morocco). Aquat. Ecosyst. Health Manag. 2005, 8, 95–104. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Monocultures of strains. | ||||||||||||||||
| Sample No | Scientific Name | Strain No. | Positive Control | |||||||||||||
| 1 | Aphanizomenon flos-aquae | NRERC-008 | no | |||||||||||||
| 2 | Aphanizomenon flos-aquae | NRERC-009 | no | |||||||||||||
| 3 | Aphanizomenon gracile | NH-5 | no | |||||||||||||
| 4 | Dolichospermum planctonicum | NRERC-101 | no | |||||||||||||
| 5 | Dolichospermum circinale | NRERC-103 | no | |||||||||||||
| 6 | Dolichospermum flos-aquae | NRERC-108 | no | |||||||||||||
| 7 | Cylindrospermopsis raciborskii | CS-1101 | yes | |||||||||||||
| 8 | Chrysosporum ovalisporum | CS-1034 | yes | |||||||||||||
| 9 | Sphaerospermopsis aphanizomenoides | NRERC-600 | yes | |||||||||||||
| 10 | Cuspidothrix issatschenkoi | NIVA CYA-711 | yes | |||||||||||||
| 11 | Raphidiopsis mediterranea | CS-1037 | yes | |||||||||||||
| (b) Mixed samples with different combinations of monocultures of strains. | ||||||||||||||||
| Sample No. Strains | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |||||||||
| Aphanizomenon flos-aquae NRERC-008 | + | + | + | + | + | + | + | |||||||||
| Aphanizomenon gracile NH-5 | + | + | + | + | + | + | + | |||||||||
| Dolichospermum planctonicum NRERC-101 | + | + | + | + | + | + | + | |||||||||
| Cylindrospermopsis raciborskii CS-1101 | + | + | + | + | + | + | + | |||||||||
| Sphaerospermopsis aphanizomenoides NRERC-600 | - | + | - | + | - | - | - | |||||||||
| Cuspidothrix issatschenkoi NIVA CYA-711 | - | - | + | + | - | - | + | |||||||||
| Raphidiopsis mediterranea CS-1037 | - | - | - | - | - | + | + | |||||||||
| Chrysosporum ovalisporum CS-1034 | - | - | - | - | + | - | - | |||||||||
| (c) Spiked field samples with different combinations of monocultures of strains. | ||||||||||||||||
| Sample No. Strains | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||||||||
| Aphanizomenon gracile NH-5 | - | + | + | + | + | + | + | + | ||||||||
| Dolichospermum planctonicum NRERC-101 | - | + | + | + | + | + | + | + | ||||||||
| Cylindrospermopsis raciborskii CS-1101 | - | + | + | + | + | + | + | + | ||||||||
| Sphaerospermopsis aphanizomenoides NRERC-600 | - | - | + | + | - | - | - | + | ||||||||
| Cuspidothrix issatschenkoi NIVA CYA-711 | - | + | - | + | - | - | + | - | ||||||||
| Raphidiopsis mediterranea CS-1037 | - | - | - | + | - | - | + | - | ||||||||
| Chrysosporum ovalisporum CS-1034 | - | - | - | - | - | + | - | + | ||||||||
| (a) 16S rRNA (1306 bp) | (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) | (10) | (11) | (12) | |||||||||||||||||||||||||||||||||||
| (1) Aphanizomenon flos-aquae | |||||||||||||||||||||||||||||||||||||||||||||||
| (2) Aphanizomenon gracile | 98.7 | ||||||||||||||||||||||||||||||||||||||||||||||
| (3) Dolichospermum planctonicum | 98.1 | 98.2 | |||||||||||||||||||||||||||||||||||||||||||||
| (4) Dolichospermum flos-aquae | 97.8 | 98.4 | 97.3 | ||||||||||||||||||||||||||||||||||||||||||||
| (5) Dolichospermum circinale | 98.6 | 98.4 | 99.3 | 97.5 | |||||||||||||||||||||||||||||||||||||||||||
| (6) Cylindrospermopsis raciborskii | 93.3 | 93.6 | 93.5 | 93.1 | 93.7 | ||||||||||||||||||||||||||||||||||||||||||
| (7) Chrysosporum ovalisporum | 95.2 | 95.6 | 95.1 | 94.7 | 95.3 | 93.5 | |||||||||||||||||||||||||||||||||||||||||
| (8) Sphaerospermopsis reniformis | 93.1 | 93.3 | 93.1 | 92.0 | 92.9 | 96.1 | 93.8 | ||||||||||||||||||||||||||||||||||||||||
| (9) Sphaerospermopsis aphanizomenoides | 93.4 | 93.7 | 93.3 | 92.4 | 93.2 | 96.7 | 94.1 | 99.5 | |||||||||||||||||||||||||||||||||||||||
| (10) Cuspidothrix issatschenkoi | 95.6 | 96.7 | 95.2 | 96.1 | 95.0 | 93.4 | 94.6 | 93.7 | 94.1 | ||||||||||||||||||||||||||||||||||||||
| (11) Raphidiopsis mediterranea | 93.4 | 93.7 | 93.6 | 93.1 | 93.8 | 99.7 | 93.5 | 96.4 | 96.9 | 93.5 | |||||||||||||||||||||||||||||||||||||
| (12) Raphidiopsis curvata | 93.1 | 93.7 | 93.3 | 93.0 | 93.4 | 99.3 | 93.0 | 96.0 | 96.5 | 93.1 | 99.2 | ||||||||||||||||||||||||||||||||||||
| (b) rbcLX(761 bp) | (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | |||||||||||||||||||||||||||||||||||||||
| (1) Aphanizomenon flos-aquae | |||||||||||||||||||||||||||||||||||||||||||||||
| (2) Aphanizomenon gracile | 100.0 | ||||||||||||||||||||||||||||||||||||||||||||||
| (3) Dolichospermum planctonicum | 94.9 | 94.9 | |||||||||||||||||||||||||||||||||||||||||||||
| (4) Dolichospermum flos-aquae | 89.0 | 89.0 | 90.9 | ||||||||||||||||||||||||||||||||||||||||||||
| (5) Dolichospermum circinale | 88.7 | 88.7 | 90.8 | 99.3 | |||||||||||||||||||||||||||||||||||||||||||
| (6) Sphaerospermopsis aphanizomenoides | 85.4 | 85.4 | 85.0 | 82.1 | 81.6 | ||||||||||||||||||||||||||||||||||||||||||
| (7) Sphaerospermopsis reniformis | 85.6 | 85.6 | 85.4 | 81.6 | 81.1 | 98.6 | |||||||||||||||||||||||||||||||||||||||||
| (8) Cuspidothrix issatschenkoi | 88.1 | 88.1 | 86.2 | 91.4 | 91.5 | 81.0 | 81.1 | ||||||||||||||||||||||||||||||||||||||||
| (c) cpcBA(357 bp) | (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) | (10) | |||||||||||||||||||||||||||||||||||||
| (1) Aphanizomenon flos-aquae | |||||||||||||||||||||||||||||||||||||||||||||||
| (2) Aphanizomenon gracile | 90.0 | ||||||||||||||||||||||||||||||||||||||||||||||
| (3) Dolichospermum planctonicum | 81.2 | 77.9 | |||||||||||||||||||||||||||||||||||||||||||||
| (4) Dolichospermum flos-aquae | 90.0 | 100.0 | 77.9 | ||||||||||||||||||||||||||||||||||||||||||||
| (5) Cylindrospermopsis raciborskii | 67.8 | 64.3 | 66.9 | 64.3 | |||||||||||||||||||||||||||||||||||||||||||
| (6) Chrysosporum ovalisporum | 66.4 | 63.5 | 66.4 | 63.5 | 68.3 | ||||||||||||||||||||||||||||||||||||||||||
| (7) Sphaerospermopsis aphanizomenoides | 74.8 | 72.9 | 75.2 | 72.9 | 77.2 | 70.9 | |||||||||||||||||||||||||||||||||||||||||
| (8) Cuspidothrix issatschenkoi | 86.2 | 83.2 | 78.0 | 83.2 | 73.4 | 71.0 | 72.8 | ||||||||||||||||||||||||||||||||||||||||
| (9) Raphidiopsis mediterranea | 66.1 | 63.2 | 66.9 | 63.2 | 98.1 | 68.1 | 76.2 | 71.8 | |||||||||||||||||||||||||||||||||||||||
| (10) Raphidiopsis curvata | 67.2 | 64.5 | 67.1 | 64.5 | 95.8 | 66.4 | 78.6 | 73.0 | 95.3 | ||||||||||||||||||||||||||||||||||||||
| (d) rpoB(504 bp) | (1) | (2) | (3) | (4) | (5) | (6) | |||||||||||||||||||||||||||||||||||||||||
| (1) Aphanizomenon flos-aquae | |||||||||||||||||||||||||||||||||||||||||||||||
| (2) Aphanizomenon gracile | 99.4 | ||||||||||||||||||||||||||||||||||||||||||||||
| (3) Dolichospermum planctonicum | 80.5 | 79.9 | |||||||||||||||||||||||||||||||||||||||||||||
| (4) Dolichospermum circinale | 74.8 | 74.8 | 85.8 | ||||||||||||||||||||||||||||||||||||||||||||
| (5) Dolichospermum flos-aquae | 74.6 | 74.6 | 69.3 | 64.7 | |||||||||||||||||||||||||||||||||||||||||||
| (6) Cuspidothrix issatschenkoi | 84.5 | 84.2 | 77.5 | 69.6 | 75.1 | ||||||||||||||||||||||||||||||||||||||||||
| (e) rpoC1(452 bp) | (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) | (10) | (11) | ||||||||||||||||||||||||||||||||||||
| (1) Aphanizomenon flos-aquae | |||||||||||||||||||||||||||||||||||||||||||||||
| (2) Aphanizomenon gracile | 78.1 | ||||||||||||||||||||||||||||||||||||||||||||||
| (3) Dolichospermum planctonicum | 77.2 | 81.2 | |||||||||||||||||||||||||||||||||||||||||||||
| (4) Dolichospermum flos-aquae | 75.4 | 74.6 | 72.9 | ||||||||||||||||||||||||||||||||||||||||||||
| (5) Dolichospermum circinale | 75.8 | 79.9 | 89.6 | 73.2 | |||||||||||||||||||||||||||||||||||||||||||
| (6) Cylindrospermopsis raciborskii | 75.1 | 71.0 | 71.6 | 69.7 | 71.2 | ||||||||||||||||||||||||||||||||||||||||||
| (7) Chrysosporum ovalisporum | 70.7 | 66.7 | 70.3 | 77.9 | 69.0 | 67.8 | |||||||||||||||||||||||||||||||||||||||||
| (8) Sphaerospermopsis aphanizomenoides | 93.5 | 77.7 | 75.9 | 74.6 | 76.4 | 76.1 | 70.7 | ||||||||||||||||||||||||||||||||||||||||
| (9) Cuspidothrix issatschenkoi | 79.8 | 79.5 | 76.3 | 74.8 | 78.4 | 73.5 | 71.9 | 80.5 | |||||||||||||||||||||||||||||||||||||||
| (10) Raphidiopsis mediterranea | 73.4 | 70.4 | 71.7 | 69.5 | 69.6 | 96.4 | 66.0 | 74.5 | 72.8 | ||||||||||||||||||||||||||||||||||||||
| (11) Raphidiopsis curvata | 76.5 | 73.2 | 74.4 | 70.8 | 72.0 | 94.2 | 69.6 | 77.1 | 72.5 | 93.7 | |||||||||||||||||||||||||||||||||||||
| Target Genus | Marker | Primer | Sequence (5′→3′) | Annealing Temperature (°C) | Product Size (bp) |
|---|---|---|---|---|---|
| Sphaerospermopsis | rbcLX | Sph-rbcLX–F | AAAATCTATGGGGCTGGGTC | 59 | 461 |
| Sph-rbcLX–R | ACTATTTGGTTTTTGGCACTTA | ||||
| Cuspidothrix | rpoB | Cus-rpoB–F | TCGCCTATTCTCACCAATGG | 58 | 496 |
| Cus-rpoB–R | ATCAAAGGTCCACAAGTACC | ||||
| Raphidiopsis | rpoC1 | Raphi-rpoC1-F | TACCCTCAAGCCAGAAATGG | 55 | 353 |
| Rahpi-rpoC1-R | TGGTCTTCTGTTAATAACTGC | ||||
| Cylindrospermopsis | rpoC1 | Cyl-rpoC1-F | ATTTTGTGAGCGGATCTTTG | 55 | 325 |
| Cyl-rpoC1-R | GGTCTTCTGTTAACAGTTGT | ||||
| Chrysosporum | cpcBA | Chry-cpcBA-F | TTGAACGGTTTGCGCGAAACC | 58 | 449 |
| Chry-cpcBA-R | ACAGCTTCGGTTGCACCATCAATT |
| No. | Station | Sampling Date | Total Phytoplankton | Diatoms | Green Algae | Other Flagellates | Cyanobacteria | ||||
| 1 | SJ | 19.08.19 | 40,990 | 130 | 6740 | 2330 | 31,790 | ||||
| 2 | ND | 19.08.05 | 98,764 | 450 | 7020 | 420 | 90,874 | ||||
| 3 | GM | 19.07.08 | 9677 | 65 | 1000 | 3350 | 5262 | ||||
| 4 | GM | 19.08.05 | 21,074 | 88 | 2665 | 159 | 18,162 | ||||
| 5 | CG | 19.07.29 | 18,464 | 2850 | 820 | 3380 | 11,414 | ||||
| 6 | CG | 19.08.26 | 15,408 | 920 | 490 | 1600 | 12,398 | ||||
| 7 | CG | 19.09.30 | 19,933 | 5700 | 3140 | 9800 | 1293 | ||||
| 8 | DS | 19.08.19 | 31,653 | 1360 | 3350 | 12,220 | 14,723 | ||||
| 9 | DS | 19.09.09 | 48,858 | 13,920 | 4860 | 1700 | 28,378 | ||||
| 10 | DS | 19.09.26 | 2761 | 1132 | 732 | 314 | 583 | ||||
| 11 | HC | 19.07.08 | 326,528 | 215 | 2675 | 595 | 323,043 | ||||
| 12 | CH | 19.09.16 | 89,470 | 5470 | 1990 | 2280 | 79,730 | ||||
| 13 | CH | 19.09.30 | 28,935 | 12,600 | 1280 | 10,640 | 4415 | ||||
| No. | Microsystis spp. | Aphanizomenon spp. | Dolichospermum spp. | Cylindrosper-mopsis spp. | Cuspidothrix spp. | Sphaerospermopsis spp. | |||||
| 1 | 8160 | 0 | 0 | 0 | 0 | 0 | |||||
| 2 | 36,350 | 534 | 28,112 | 0 | 1937 | 308 | |||||
| 3 | 1025 | 100 | 2503 | 0 | 254 | 0 | |||||
| 4 | 11,810 | 169 | 3469 | 0 | 755 | 25 | |||||
| 5 | 1765 | 0 | 0 | 0 | 39 | 0 | |||||
| 6 | 7110 | 30 | 0 | 0 | 0 | 88 | |||||
| 7 | 755 | 41 | 0 | 77 | 120 | 0 | |||||
| 8 | 7335 | 5435 | 964 | 0 | 29 | 0 | |||||
| 9 | 22,380 | 0 | 10 | 0 | 0 | 48 | |||||
| 10 | 350 | 30 | 18 | 23 | 0 | 25 | |||||
| 11 | 258,800 | 4624 | 179 | 0 | 0 | 0 | |||||
| 12 | 65,500 | 595 | 1122 | 0 | 170 | 103 | |||||
| 13 | 1125 | 239 | 153 | 0 | 21 | 0 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, I.-S.; Park, H.-K.; Kim, Y.-J. Development of Genus-Specific PCR Primers for Molecular Monitoring of Invasive Nostocalean Cyanobacteria. Int. J. Environ. Res. Public Health 2021, 18, 5703. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18115703
Kim I-S, Park H-K, Kim Y-J. Development of Genus-Specific PCR Primers for Molecular Monitoring of Invasive Nostocalean Cyanobacteria. International Journal of Environmental Research and Public Health. 2021; 18(11):5703. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18115703
Chicago/Turabian StyleKim, In-Soo, Hae-Kyung Park, and Yong-Jin Kim. 2021. "Development of Genus-Specific PCR Primers for Molecular Monitoring of Invasive Nostocalean Cyanobacteria" International Journal of Environmental Research and Public Health 18, no. 11: 5703. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18115703