
The Olfactory System as Marker of Neurodegeneration in Aging, Neurological and Neuropsychiatric Disorders


{kind=link}
Abstract
:1. Introduction
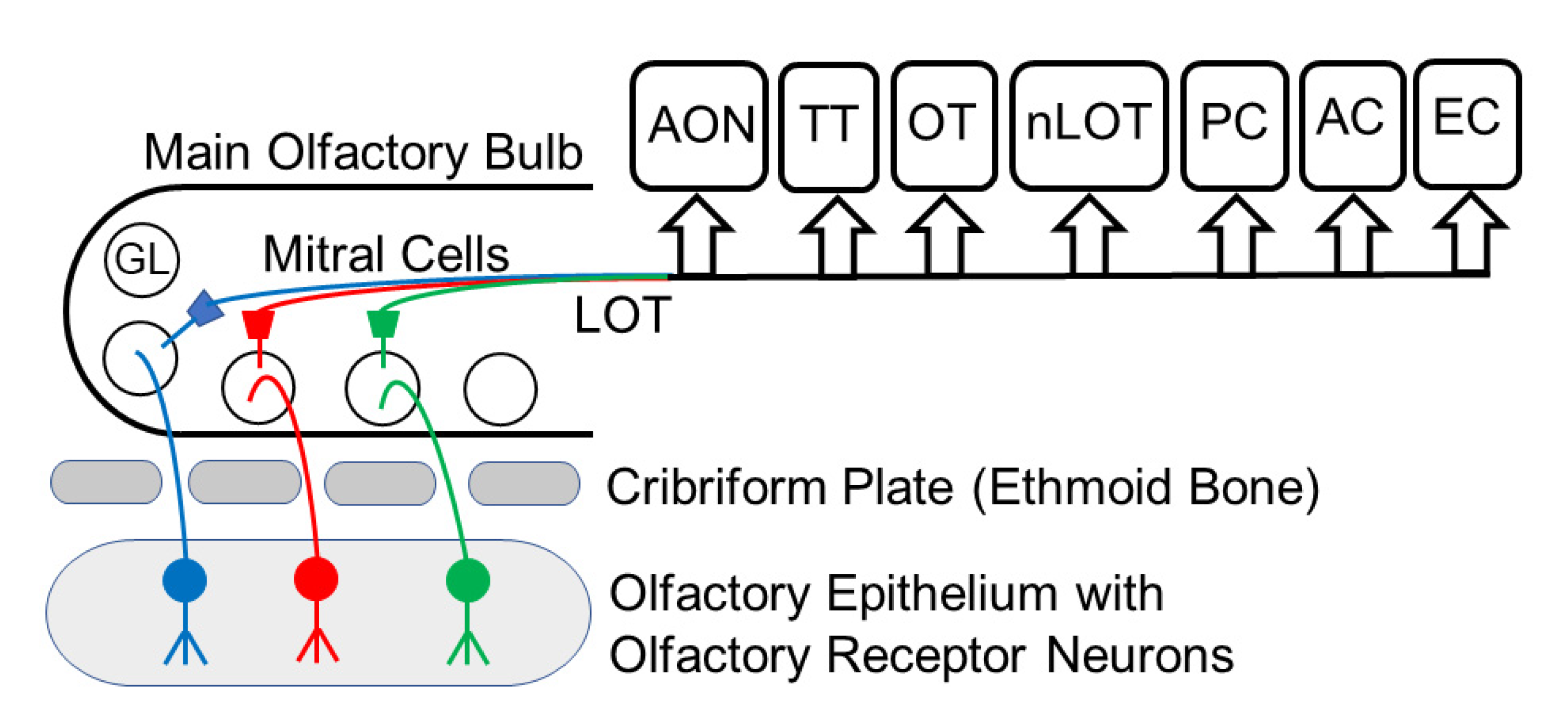
2. The Olfactory Pathway
3. Aging and Olfaction
4. Neuropsychiatric Disorders
5. Neurodegenerative Diseases
6. Olfactory Function, Memory, and Genes
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ennis, M.; Hamilton, K.A.; Hayar, A. Neurochemistry of the main olfactory system. In Handbook of Neurochemistry and Molecular Neurobiology: Sensory Neurochemistry; Springer: New York, NY, USA, 2007; pp. 137–204. [Google Scholar]
- Heinbockel, T.; Heyward, P.M. Glutamate synapses in olfactory neural circuits. In Amino Acid Receptor Research; Paley, B.F., Warfield, T.E., Eds.; Nova Science Publishers: New York, NY, USA, 2009; Volume 16, pp. 379–414. [Google Scholar]
- Shipley, M.T.; Ennis, M. Functional organization of olfactory system. J. Neurobiol. 1996, 30, 123–176. [Google Scholar] [CrossRef]
- Swanson, L.W. Brain Maps: Structure of the Rat Brain. A Laboratory Guide with Printed and Electronic Templates for Data, Models and Schematics, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Diaz, D.; Gomez, C.; Munoz-Castaneda, R.; Alfonso, J.R.; Weruaga, E. The olfactory system as a puzzle: Playing with its pieces. Anat. Rec. 2013, 296, 1383–1400. [Google Scholar] [CrossRef]
- Bryche, B.; St Albin, A.; Murri, S.; Lacôte, S.; Pulido, C.; Ar Gouilh, M.; Lesellier, S.; Servat, A.; Wasniewski, M.; Picard-Meyer, E.; et al. Massive transient damage of the olfactory epithelium associated with infection of sustentacular cells by SARS-CoV-2 in golden Syrian hamsters. Brain Behav. Immun. 2020, 89, 579–586. [Google Scholar] [CrossRef]
- Rey, N.L.; Wesson, D.W.; Brundin, P. The olfactory bulb as the entry site for prion-like propagation in neurodegenerative diseases. Neurobiol. Dis. 2018, 109, 226–248. [Google Scholar] [CrossRef]
- Bhatia-Dey, N.; Heinbockel, T. Neurological and Neuropsychiatric Disorders in Relation to Olfactory Dysfunction. In Sino-Nasal and Olfactory System Disorders; Heinbockel, T., Gendeh, B.S., Eds.; Intech Open: London, UK, 2020; Volume 7, p. 18. [Google Scholar] [CrossRef]
- Shepherd, G.M. The human sense of smell: Are we better than we think? PLoS Biol. 2004, 2, e146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadiou, H.; Aoudé, I.; Tazir, B.; Molinas, A.; Fenech, C.; Meunier, M.; Grosmaitre, X. Postnatal odorant exposure induces peripheral olfactory plasticity at the cellular level. J. Neurosci. 2014, 34, 4857. [Google Scholar] [CrossRef] [Green Version]
- Joussain, P.; Thevenet, M.; Rouby, C.; Bensafi, M. Effect of Aging on Hedonic Appreciation of Pleasant and Unpleasant Odors. PLoS ONE 2013, 8, e61376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joussain, P.; Rouby, C.; Bensafi, M. A pleasant familiar odor influences perceived stress and peripheral nervous system activity during normal aging. Front. Psychol. 2014, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Boesveldt, S.C.; de Graaf, C.; de Wijk, R.A. Dynamics of autonomic nervous system responses and facial expressions to odors. Front. Psychol. 2014, 5, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGann, J.P. Poor human olfaction is a 19th-century myth. Science 2017, 356, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Li, W.; Wu, X.; Li, J.; Yang, J.; Tu, C.; Ye, X.; Ling, S. Olfactory deficit is associated with mitral cell dysfunction in the olfactory bulb of P301S tau transgenic mice. Brain Res. Bull. 2019, 148, 34–45. [Google Scholar] [CrossRef]
- Bathini, P.; Brai, E.; Auber, L.A. Olfactory dysfunction in the pathophysiological continuum of dementia. Ageing Res. Rev. 2019, 55, 100956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadohisa, M. Effects of odor on emotion, with implications. Front. Syst. Neurosci. 2013, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, G.M.; Chen, W.R.; Greer, C.A. Olfactory Bulb. In Synaptic Organization of the Brain, 5th ed.; Shepherd, G.M., Ed.; Oxford University Press: New York, NY, USA, 2004; pp. 165–216. [Google Scholar]
- Pashkovski, S.L.; Lurilli, G.; Brann, D.; Chicharro, D.; Drummey, K.; Franks, K.; Panzeri, S.; Datta, S.R. Structure and flexibility in cortical representations of odour space. Nature 2020, 583, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Zamparo, I.; Francia, S.; Franchi, S.A.; Redolfi, N.; Costanzi, E.; Kerstens, A.; Fukutani, Y.; Battistutta, R.; de Laureto, P.P.; Munck, S.; et al. Axonal odorant receptors mediate axon targeting. Cell Rep. 2019, 29, 4334–4348.e7. [Google Scholar] [CrossRef] [Green Version]
- Rebholz, H.; Braun, R.J.; Ladage, D.; Knoll, W.; Kleber, C.; Hassel, W. Loss of Olfactory Function-Early Indicator for Covid-19, Other Viral Infections and Neurodegenerative Disorders. Front. Neurol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Hanchate, N.K.; Kondoh, K.; Lu, Z.; Kuang, D.; Ye, X.; Qiu, X.; Pachter, L.; Trapnell, C.; Buck, L.B. Single-cell transcriptomics reveals receptor transformations during olfactory neurogenesis. Science 2015, 350, 1251–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.; Li, Q.; Xie, X.S. Olfactory sensory neurons transiently express multiple olfactory receptors during development. Mol. Syst. Biol. 2015, 11, 844. [Google Scholar] [CrossRef]
- Brann, J.H.; Firestein, S.J. A lifetime of neurogenesis in the olfactory system. Front. Neurosci. 2014, 8, 182. [Google Scholar] [CrossRef]
- Brozzetti, L.; Sacchetto, L.; Cecchini, M.P.; Avesani, A.; Perra, D.; Bongianni, M.; Portioli, C.; Scupoli, M.; Ghetti, B.; Monaco, S.; et al. Neurodegeneration-Associated Proteins in Human Olfactory Neurons Collected by Nasal Brushing. Front. Neurosci. 2020, 14, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodato, M.A.; Rodin, R.E.; Bohrson, C.L.; Coulter, M.E.; Barton, A.R.; Kwon, M.; Yang, P.; Chittenden, T.W.; Hatem, N.E.; Ryu, S.C.; et al. Aging and neurodegeneration are associated with increased mutations in single human neurons. Science 2018, 359, 555–559. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Chen, Z.; Shang, C.; Yan, F.; Shi, Y.; Zhang, J.; Qu, B.; Han, H.; Wang, Y.; Li, D.; et al. IGF1-dependent synaptic plasticity of mitral cells in olfactory memory during social learning. Neuron 2017, 95, 106–122. [Google Scholar] [CrossRef] [Green Version]
- Doty, R.L. The olfactory system and its disorders. Semin. Neurol. 2009, 29, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Makowska, I.; Kloszewska, I.; Grabowska, A.; Szatkowska, I.; Rymarczyk, K. Olfactory deficits in normal aging and Alzheimer’s disease in the polish elderly population. Arch. Clin. Neuropsychol. 2011, 26, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Hüttenbrink, K.B.; Hummel, T.; Berg, D.; Gasser, T.; Hähner, A. Olfactory dysfunction: Common in later life and early warning of neurodegenerative disease. Dtsch Arztebl. Int. 2013, 110, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Teranishi, M.; Katayama, N.; Nakashima, T.; Sugiura, S.; Sone, M. Relationship between cognitive impairment and olfactory function among older adults with olfactory impairment. Auris Nasus Larynx 2021, 48, 420–427. [Google Scholar] [CrossRef]
- Tzeng, W.Y.; Figarella, K.; Garaschuk, O. Olfactory impairment in men and mice related to aging and amyloid-induced pathology. Pflug. Arch. 2021, 473, 805–821. [Google Scholar] [CrossRef] [PubMed]
- Mobley, A.S.; Rodriguez-Gil, D.J.; Imamura, F.; Greer, C.A. Aging in the olfactory system. Trends Neurosci. 2013, 37, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Doty, R.L.; Kamath, V. The influences of age on olfaction: A review. Front. Psychol. 2014, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Attems, J.; Walker, L.; Jellinger, K.A. Olfaction and aging: A mini-review. Gerontology 2015, 61, 485–490. [Google Scholar] [CrossRef]
- Kalmey, J.K.; Thewissen, J.G.; Dluzen, D.E. Age-related size reduction of foramina in the cribriform plate. Anat. Rec. 1998, 251, 326–329. [Google Scholar] [CrossRef]
- Paik, S.I.; Lehman, M.N.; Seiden, A.M.; Duncan, H.J.; Smith, D.V. Human Olfactory Biopsy: The Influence of Age and Receptor Distribution. Arch. Otolaryngol. Head Neck Surg. 1992, 118, 731–738. [Google Scholar] [CrossRef]
- Buschhuter, D.; Smitka, M.; Puschmann, S.; Gerber, J.C.; Witt, M.; Abolmaali, N.D.; Hummel, T. Correlation between olfactory bulb volume and olfactory function. Neuroimage 2008, 42, 498–502. [Google Scholar] [CrossRef]
- Yousem, D.M.; Geckle, R.J.; Bilker, W.B.; McKeown, D.A.; Doty, R.L. Posttraumatic olfactory dysfunction: MR and clinical evaluation. AJNR 1996, 17, 1171–1179. [Google Scholar]
- Yousem, D.M.; Geckle, R.J.; Bilker, W.B.; Kroger, H.; Doty, R.L. Posttraumatic smell loss: Relationship of psychophysical tests and volumes of the olfactory bulbs and tracts and the temporal lobes. Acad. Radiol. 1999, 6, 264–272. [Google Scholar] [CrossRef]
- Ueha, R.; Shichino, S.; Ueha, S.; Kondo, K.; Kikuta, S.; Nishijima, H.; Matsushima, K.; Yamasoba, T. Reduction of proliferating olfactory cells and low expression of extracellular matrix genes are hallmarks of the aged olfactory mucosa. Front. Aging Neurosci. 2018, 10, 1–13. [Google Scholar] [CrossRef]
- Segura, B.; Baggio, H.C.; Solana, E.; Palacios, E.M.; Vendrell, P.; Bargallo, N.; Junque, C. Neuroanatomical correlates of olfactory loss in normal aged subjects. Behav. Brain Res. 2013, 246, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.H.; Baxter, M.G. The ageing cortical synapse: Hallmarks and implications for cognitive decline. Nat. Rev. Neurosci. 2012, 13, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Habib, N.; McCabe, C.; Medina, S.; Varshavsky, M.; Kitsberg, D.; Dvir-Sternfeld, R.; Green, G.; Dionne, D.; Nguyen, L.; Marshall, J.L.; et al. Disease-associated astrocytes in Alzheimer’s disease and aging. Nat. Neurosci. 2020, 23, 701–706. [Google Scholar] [CrossRef]
- Rawson, N.E.; Gomez, G.; Cowart, B.J.; Kriete, A.; Pribitkin, E.; Restrepo, D. Age-associated loss of selectivity in human olfactory sensory neurons. Neurobiol. Aging 2012, 33, 1913–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trimmer, C.; Keller, A.; Murphy, N.R.; Snyder, L.L.; Willer, J.R.; Nagai, M.H.; Katsanis, N.; Vosshall, L.B.; Matsunami, H.; Mainland, J.D. Genetic variation across the human olfactory receptor repertoire alters odor perception. Proc. Natl. Acad. Sci. USA 2019, 16, 9475–9948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaffe, K.; Freimer, D.; Chen, H.; Asao, K.; Rosso, A.; Rubin, S.; Tranah, G.; Cummings, S.; Simonsick, E. Olfaction and risk of dementia in a biracial cohort of older adults. Neurology 2017, 88, 456–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Regemorter, V.; Hummel, T.; Rosenzweig, F.; Mouraux, A.; Rombaux, P.; Huart, C. Mechanisms Linking Olfactory Impairment and Risk of Mortality. Front. Neurosci. 2020, 14, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, S.M.; Parker, T.D.; Lane, C.A.; Keshavan, A.; Keuss, S.E.; Lu, K.; James, S.-N.; Murray-Smith, H.; Wong, A.; Nicholas, J.; et al. Olfactory testing does not predict β-amyloid, MRI measures of neurodegeneration or vascular pathology in the British 1946 birth cohort. J. Neurol. 2020, 267, 3329–3336. [Google Scholar] [CrossRef]
- Ryo, Y.; Takeuchi, M.; Ueda, N.; Ohi, K.; Kihara, H.; Shimada, T.; Uehara, T.; Kawasaki, Y. Olfactory function in neuropsychiatric disorders. Psychiatry Res. 2017, 252, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Luzzi, S.; Snowden, J.S.; Neary, D.; Coccia, M.; Provinciali, L.; Ralph, M.A.L. Distinct patterns of olfactory impairment in Alzheimer’s disease, semantic dementia, frontotemporal dementia, and corticobasal degeneration. Neuropsychologia 2007, 45, 1823–1831. [Google Scholar] [CrossRef]
- McLaughlin, N.C.; Westervelt, H.J. Odor identification deficits in frontotemporal dementia: A preliminary study. Arch. Clin. Neuropsychol. 2008, 23, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Taalman, H.; Wallace, C.; Milev, R. Olfactory Functioning and Depression: A Systematic Review. Front. Psychiatry 2017, 8, 190. [Google Scholar] [CrossRef] [Green Version]
- Zucco, G.M.; Bollini, F. Odour recognition memory and odour identification in patients with mild and severe major depressive disorders. Psychiatry Res. 2011, 190, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Croy, I.; Springborn, M.; Lötsch, J.; Johnston, A.N.; Hummel, T. Agreeable smellers and sensitivity neurotics—correlations among personality traits and sensory thresholds. PLoS ONE 2011, 6, e18701. [Google Scholar] [CrossRef] [Green Version]
- Kohli, P.; Soler, Z.M.; Nguyen, S.A.; Muus, J.S.; Schlosser, R.J. The association between olfaction and depression: A systematic review. Chem. Senses 2016, 41, 479–486. [Google Scholar] [CrossRef] [Green Version]
- Dileo, J.F.; Brewer, W.J.; Hopwood, M.; Anderson, V.; Creamer, M. Olfactory identification dysfunction, aggression and impulsivity in war veterans with post-traumatic stress disorder. Psychol. Med. 2008, 38, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Negoias, S.; Hummel, T.; Symmank, A.; Schellong, J.; Joraschky, P.; Croy, I. Olfactory bulb volume predicts therapeutic outcome in major depression disorder. Brain Imaging Behav. 2015, 10, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Orasji, S.S.S.; Mulder, J.L.; de Bruijn, S.F.T.M.; Wirtz, P.W. Olfactory dysfunction in behavioral variant frontotemporal dementia. Clin. Neurol. Neurosurg. 2016, 141, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Carnemolla, S.E.; Hsieh, J.W.; Sipione, R.; Landis, B.N.; Kumfor, F.; Piguet, O.; Manuel, A.L. Olfactory dysfunction in frontotemporal dementia and psychiatric disorders: A systematic review. Neurosci. Biobehav. Rev. 2020, 118, 588–611. [Google Scholar] [CrossRef] [PubMed]
- Rochet, M.; El-Hage, W.; Richa, S.; Kazour, F.; Atanasova, B. Depression, olfaction, and quality of life: A mutual relationship. Brain Sci. 2018, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savic, I.; Gulyas, B.; Larsson, M.; Roland, P. Olfactory functions are mediated by parallel and hierarchical processing. Neuron 2000, 26, 735–745. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.K.; Christoff, K.; Stappen, I.; Panitz, D.; Ghahremani, D.G.; Glover, G.; Gabrieli, J.D.E.; Sobel, N. Dissociated neural representations of intensity and valence in human olfaction. Nat. Neurosci. 2003, 6, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Kiracanski, K.; Joormann, J.; Gotlib, I.H. Cognitive aspects of depression: Cognitive aspects of depression. Wiley Interdiscip. Rev. Cogn. Sci. 2012, 3, 301–313. [Google Scholar]
- Wicker, B.; Monfardini, E.; Royet, J.-P. Olfactory processing in adults with autism spectrum disorders. Mol. Autism 2016, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kamath, V.; Paksarian, D.; Cui, L.; Moberg, P.J.; Turetsky, B.I.; Merikangas, K.R. Olfactory processing in bipolar disorder, major depression, and anxiety. Bipolar Disord. 2018, 20, 547–555. [Google Scholar] [CrossRef]
- Iakoucheva, L.M.; Muotri, A.R.; Sebat, J. Getting to the cores of autism. Cell 2019, 178, 1287–1298. [Google Scholar] [CrossRef]
- Dawson, P.A.; Lee, S.; Ewing, A.D.; Prins, J.B.; Heussler, H.S. Whole genome investigation of an atypical autism case identifies a novel ANOS1 mutation with subsequent diagnosis of Kallmann syndrome. Mol. Genet. Metabol. Rep. 2020, 23, 100593. [Google Scholar] [CrossRef] [PubMed]
- Briscoe, J.; Marin, O. Looking at neurodevelopment through a big data lens. Science 2020, 369. [Google Scholar] [CrossRef] [PubMed]
- Satterstrom, F.K.; Kosmicki, J.A.; Wang, J.; Breen, M.S.; De Rubeis, S.; An, J.-Y.; Peng, M.; Collins, R.; Grove, J.; Klei, L.; et al. Large-scale exome sequencing study implicates both developmental and functional changes in the neurobiology of autism. Cell 2020, 180, 568–584. [Google Scholar] [CrossRef] [PubMed]
- Geramita, M.A.; Wen, J.A.; Rannals, M.D.; Urban, N.N. Decreased amplitude and reliability of odor-evoked responses in two mouse models of autism. J. Neurophys. 2020, 123, 1283–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Hu, Y.; Cadman, S.; Bouloux, P. Diversity in fibroblast growth factor receptor1 regulation: Learning from the investigation of Kallmann syndrome. J. Neuroendocrinol. 2008, 20, 141–163. [Google Scholar] [CrossRef] [PubMed]
- Riley, J.D.; Delahunty, C.; Alsadah, A.; Mazzola, S. Further evidence of GABRA4 and TOP3B as autism susceptibility genes. Eur. J. Med. Genet. 2020, 63, 103876. [Google Scholar] [CrossRef]
- Benarroch, E.E. Olfactory system: Functional organization and involvement in neurodegenerative disease. Neurology 2010, 75, 1104–1109. [Google Scholar] [CrossRef]
- Acebes, A.; Martín-Peña, A.; Chevalier, V.; Ferrús, A. Synapse loss in olfactory local interneurons modifies perception. J. Neurosci. 2011, 31, 2734–2745. [Google Scholar] [CrossRef] [Green Version]
- Barresi, M.; Ciurleo, R.; Giacoppo, S.; Foti, C.V.; Celi, D.; Bramanti, P. Evaluation of olfactory dysfunction in neurodegenerative diseases. J. Neurol. Sci. 2012, 323, 16–24. [Google Scholar] [CrossRef]
- Bhatia-Dey, N.; Kanherkar, R.R.; Stair, S.E.; Makarev, E.O.; Csoka, A.B. Cellular Senescence as the Causal Nexus of Aging. Front. Genet. 2016, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef]
- Khosla, S.; Farr, J.N.; Tchkonia, T.; Kirkland, J.L. The role of cellular senescence in ageing and endocrine disease. Nat. Rev. Endocrinol. 2020, 16, 263–275. [Google Scholar] [CrossRef]
- Campisi, J.; Kapahi, P.; Lithgow, G.J.; Melov, S.; Newman, J.C.; Verdin, E. From discoveries in ageing research to therapeutics for healthy ageing. Nature 2019, 571, 183–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.L.; Sierra, F.; Howcroft, K.; Singer, D.S.; Sharpless, N.; Hodes, R.J.; Wilder, E.L.; Anderson, J.M. A Blueprint for Characterizing Senescence. Cell 2020, 183, 1143–1146. [Google Scholar] [CrossRef]
- Hoang, M.L.; Kinde, I.; Tomasetti, C.; McMahon, K.W.; Rosenquist, T.A.; Grollman, A.P.; Kinzlera, K.W.; Vogelstein, B.; Papadopoulos, N. Genome-wide quantification of rare somatic mutations in normal human tissues using massively parallel sequencing. Proc. Natl. Acad. Sci. USA 2016, 113, 9846–9851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olender, T.; Waszak, S.M.; Viavant, M.; Khen, M.; Ben-Asher, E.; Reyes, A.; Nativ, N.; Wysocki, C.J.; Ge, D.; Lancet, D. Personal receptor repertoires: Olfaction as a model. BMC Genom. 2012, 13, 414. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, J.; Jones-Gotman, M.; De Sousa, K.; Chertkow, H. Olfaction in patients with mild cognitive impairment and Alzheimer’s disease. Neurobiol. Aging 2008, 29, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L. Olfactory dysfunction in neurodegenerative diseases: Is there a common pathological substrate? Lancet Neurol. 2017, 16, 478–488. [Google Scholar] [CrossRef]
- Marin, C.; Vilas, D.; Langdon, C.; Alobid, I.; López-Chacón, M.; Haehner, A.; Hummel, T.; Mullol, J. Olfactory dysfunction in neurodegenerative diseases. Curr. Allergy Asthma Rep. 2018, 18, 42. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L.; Hawkes, C.H. Chemosensory dysfunction in neurodegenerative diseases. Smell Taste 2019, 4, 325–360. [Google Scholar]
- Sedghizadeh, M.J.; Hojjati, H.; Ezzatdoost, K.; Aghajan, H.; Vahabi, Z.; Tarighatnia, H. Olfactory response as a marker for Alzheimer’s disease: Evidence from perceptual and frontal lobe oscillation coherence deficit. PLoS ONE 2020, 15, e0243535. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.H.V.; Chaudhuri, K.R.; Jenner, P. Non-motor features of Parkinson’ disease. Nat. Rev. Neurosci. 2017, 18, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.C.G.; Silva, F.G.; Costa, L.O.P.; Freitas, S.M.S.F. Smell tests to distinguish Parkinson’s disease from other neurological disorders: A systematic review and meta-analysis. Expert Rev. Neurother. 2021, 21, 365–379. [Google Scholar] [CrossRef]
- Scheltens, P.; Blennow, K.; Breteler, M.M.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet 2016, 388, 505–517. [Google Scholar] [CrossRef]
- Tromp, D.; Dufour, A.; Lithfous, S.; Pebayle, T.; Despres, O. Episodic memory in normal aging and Alzheimer disease: Insights from imaging and behavioral studies. Ageing Res. Rev. 2015, 24, 232–262. [Google Scholar] [CrossRef]
- Ubeda-Bañon, I.; Saiz-Sanchez, D.; Flores-Cuadrado, A.; Rioja-Corroto, E.; Gonzalez-Rodriguez, M.; Villar-Conde, S.; Astillero-Lopez, V.; Cabello-de la Rosa, J.P.; Gallardo-Alcañiz, M.J.; Vaamonde-Gamo, J.; et al. The human olfactory system in two proteinopathies: Alzheimer’s and Parkinson’s diseases. Transl. Neurodegener. 2020, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Franks, K.H.; Chuah, M.I.; King, A.E.; Vickers, J.C. Connectivity of Pathology: The Olfactory System as a Model for Network-Driven Mechanisms of Alzheimer’s Disease Pathogenesis. Front. Aging Neurosci. 2015, 7, 234. [Google Scholar] [CrossRef] [Green Version]
- Ter Laak, H.J.; Renkawek, K.; van Workum, F.P. The olfactory bulb in Alzheimer disease: A morphologic study of neuron loss, tangles, and senile plaques in relation to olfaction. Alzheimer Dis. Assoc. Disord. 1994, 8, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Mundinano, I.C.; Caballero, M.C.; Ordonez, C.; Hernandez, M.; DiCaudo, C.; Marcilla, I.; Erro, M.E.; Tunon, M.T.; Luquin, M.R. Increased dopaminergic cells and protein aggregates in the olfactory bulb of patients with neurodegenerative disorders. Acta Neuropathol. 2011, 122, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.A.; Ostaszewski, M.; Matsuoka, Y.; Ghosh, S.; Glaab, E.; Trefois, C.; Crespo, I.; Perumal, T.M.; Jurkowsky, W.; Antony, P.M.A.; et al. Integrating pathways of Parkinson’s disease in a molecular interaction map. Mol. Neurobiol. 2014, 49, 88–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doty, R.L. Olfactory dysfunction in Parkinson disease. Nat. Rev. Neurol. 2012, 8, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Ponsen, M.M.; Stoffers, D.; Booij, J.; van Eck-Smit, B.L.; Wolters, E.; Berendse, H.W. Idiopathic hyposmia as a preclinical sign of Parkinson’s disease. Ann. Neurol. 2004, 56, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Wattendorf, E.; Welge-Lüssen, A.; Fiedler, K.; Bilecen, D.; Wolfensberger, M.; Fuhr, P.; Hummel, T.; Westermann, B. Olfactory impairment predicts brain atrophy in Parkinson’s disease. J. Neurosci. 2009, 29, 15410–15413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Yu, C.; Fan, F.; Zhang, K.; Zhu, C.; Wu, T.; Li, K.; Chan, P. Correlation between progressive changes in piriform cortex and olfactory performance in early Parkinson’s disease. Eur. Neurol. 2011, 66, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Spillantini, M.G.; Del Tredici, K.; Braak, H. 100 years of Lewy pathology. Nat. Rev. Neurol. 2013, 9, 13–24. [Google Scholar] [CrossRef]
- Takeda, A.; Baba, T.; Kikuchi, A.; Hasegawa, T.; Sugeno, N.; Konno, M.; Miura, E.; Mori, E. Olfactory dysfunction and dementia in Parkinson’s disease. J. Parkinsons Dis. 2014, 4, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Fullard, M.E.; Morley, J.F.; Duda, J.E. Olfactory dysfunction as an early biomarker in Parkinson’s disease. Neurosci. Bull. 2017, 33, 515–525. [Google Scholar] [CrossRef]
- Espay, A.J.; Kalia, L.V.; Gan-Or, Z. Disease modification and biomarker development in Parkinson disease: Revision or reconstruction? Neurology 2020, 94, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Pearce, R.K.; Hawkes, C.H.; Daniel, S.E. The anterior olfactory nucleus in Parkinson’s disease. Mov. Disord. 1995, 10, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Huisman, E.; Uylings, H.B.; Hoogland, P.V. A 100% increase of dopaminergic cells in the olfactory bulb may explain hyposmia in Parkinson’s disease. Mov. Disord. 2004, 19, 687–692. [Google Scholar] [CrossRef]
- Price, J.L. Olfactory System. In The Human Nervous System; Paxinos, G., Ed.; Academic Press: San Diego, CA, USA, 1990; pp. 979–998. [Google Scholar]
- Crosby, E.C.; Humphrey, T. Studies of the vertebrate telencephalon. II. The nuclear pattern of the anterior olfactory nucleus, tuberculum olfactorium and the amygdaloid complex in adult man. J. Comp. Neurol. 1941, 74, 193–352. [Google Scholar] [CrossRef] [Green Version]
- Ubeda-Bañon, I.; Saiz-Sanchez, D.; de la Rosa-Prieto, C.; Argandona-Palacios, L.; Garcia-Munozguren, S.; Martinez-Marcos, A. alpha-Synucleinopathy in the human olfactory system in Parkinson’s disease: Involvement of calcium binding protein and substance P-positive cells. Acta Neuropathol. 2010, 119, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Ubeda-Bañon, I.; Flores-Cuadrado, A.; Saiz-Sanchez, D.; Martinez-Marcos, A. Differential Effects of Parkinson’s Disease on Interneuron Subtypes within the Human Anterior Olfactory Nucleus. Front. Neuroanat. 2017, 11, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiz-Sanchez, D.; Ubeda-Bañon, I.; de la Rosa-Prieto, C.; Argandoña-Palacios, L.; Garcia-Munozguren, S.; Insausti, R.; Martinez-Marcos, A. Somatostatin, tau, and β-amyloid within the anterior olfactory nucleus in Alzheimer disease. Exp. Neurol. 2010, 223, 347–350. [Google Scholar] [CrossRef]
- Meldolesi, J. Astrocytes: News about Brain Health and Diseases. Biomedicines 2020, 8, 394. [Google Scholar] [CrossRef]
- Stevenson, T.J.; Murray, H.C.; Turner, C.; Turner, C.; Faull, R.L.M.; Dieriks, B.V.; Curtis, M.A. α-synuclein inclusions are abundant in non-neuronal cells in the anterior olfactory nucleus of the Parkinson’s disease olfactory bulb. Sci. Rep. 2020, 10, 6682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, Z.; Zheng, S.; Zhang, X.; Mai, Y.; Pan, J.; Hummel, T.; Hähner, A.; Zou, L. Olfactory impairment as an early marker of Parkinson’s disease in REM sleep behavior disorder: A systematic review and meta-analysis. J. Neurol. Neurosurg. Psychiatry 2021, 92, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Winchester, R.L.; Martyn, K. Could Early Identification of Changes in Olfactory Function Be an Indicator of Preclinical Neurodegenerative Disease? A Systematic Review. Neurol. Ther. 2020, 9, 243–263. [Google Scholar] [CrossRef]
- Hedner, M.; Nilsson, L.G.; Olofsson, J.K.; Bergman, O.; Eriksson, E.; Nyberg, L.; Larsson, M. Age-related olfactory decline is associated with the BDNF Val66met polymorphism: Evidence from a population-based study. Front. Aging Neurosci. 2010, 2, 24. [Google Scholar] [CrossRef]
- Le, X.T.; Pham, H.T.N.; Do, P.T.; Fuziwara, H.; Tanka, K.; Li, F.; Nguyen, T.V.; Nguyen, K.M.; Matsumoto, K. Bacopa monnieri Ameliorates Memory Deficits in Olfactory Bulbectomized Mice: Possible Involvement of Glutamatergic and Cholinergic Systems. Neurochem. Res. 2013, 38, 2201–2215. [Google Scholar] [CrossRef]
- Olender, T.; Keydar, I.; Pinto, J.M.; Tatarskyy, P.; Alkelai, A.; Chien, M.-S.; Fishilevich, S.; Restrepo, D.; Matsunami, H.; Gilad, Y.; et al. The human olfactory transcriptome. BMC Genom. 2016, 17, 619. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, S.; Shimizu, A.; Kondo, M.; Asano, H.; Ueno, N.; Nakayama, H.; Sato, N.; Komeno, M.; Ogita, H.; Kurokawa-Seo, M. Anosmin-1 activates vascular endothelial growth factor receptor and its related signaling pathway for olfactory bulb angiogenesis. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Kaufman, C.S.; Genovese, A.; Butler, M.G. Deletion of TOP3B Is Associated with Cognitive Impairment and Facial Dysmorphism. Cytogenet. Genom. Res. 2016, 150, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Wang, W. Roles of Topoisomerases in Heterochromatin, Aging, and Diseases. Genes 2019, 10, 884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucassen, E.B.; Turel, A.; Knehans, A.; Huang, X.; Eslinger, P. Olfactory dysfunction in multiple sclerosis: A scoping review of the literature. Mult. Scler. Relat. Disord. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- da Silva, A.M.; Torres, C.; Ferreira, I.; Moreira, I.; Samões, R.; Sousa, A.P.; Santos, E.; Teixeira-Pinto, A.; Cavaco, S. Prognostic value of odor identification impairment in multiple sclerosis: 10-Years follow-up. Mult. Scler. Relat. Disord. 2020, 46, 102486. [Google Scholar] [CrossRef] [PubMed]
- Brann, D.H.; Datta, S. R Finding the brain in the nose. Ann. Rev. Neurosci. 2020, 4, 277–295. [Google Scholar] [CrossRef]
- Sorokowska, A.; Sorokowski, P.; Hummel, T.; Huanca, T. Olfaction and Environment: Tsimane’ of Bolivian Rainforest Have Lower Threshold of Odor Detection Than Industrialized German People. PLoS ONE 2013, 8, e69203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajmani, G.S.; Suh, H.H.; Pinto, J.M. Effects of ambient air pollution exposure on olfaction: A review. Environ. Health Perspect. 2016, 124, 1683–1693. [Google Scholar] [CrossRef]
- Chronister, W.D.; Burbulis, I.E.; Wierman, M.B.; Wolpert, M.J.; Haakenson, M.F.; Smith, A.C.B.; Kleinman, J.E.; Hyde, T.M.; Weinberger, D.R.; Bekiranov, S.; et al. Neurons with Complex Karyotypes Are Rare in Aged Human Neocortex. Cell Rep. 2019, 26, 825–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doty, R.L.; Petersen, I.; Mensah, N.; Christensen, K. Genetic and environmental influences on odor identification ability in the very old. Psychol. Aging 2011, 26, 864–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Young, J.M.; Friedman, C.; Williams, E.M.; Ross, J.A.; Tonnes-Priddy, L.; Trask, B.J. Different evolutionary processes shaped the mouse and human olfactory receptor gene families. Hum. Mol. Genet. 2002, 5, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Olender, T.; Jones, T.E.; Bruford, E.; Lancet, D.A. A unified nomenclature for vertebrate olfactory receptors. BMC Evol. Biol. 2020, 20, 42. [Google Scholar] [CrossRef] [Green Version]
- Niimura, Y. Evolutionary dynamics of olfactory receptor genes in chordates: Interaction between environments and genomic contents. Hum. Genom. 2009, 4, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bear, D.M.; Lassance, J.M.; Hoekstra, H.E.; Datta, S.R. The evolving neural and genetic architecture of vertebrate olfaction. Curr. Biol. 2016, 26, R1039–R1049. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, L.R.; Riveros-McKay, F.; Mezzavilla, M.; Abou-Moussa, E.H.; Arayata, C.J.; Makhlouf, M.; Trimmer, C.; Ibarra-Soria, X.; Khan, M.; Van Gerven, L.; et al. A transcriptomic atlas of mammalian olfactory mucosae reveals an evolutionary influence on food odor detection in humans. Sci. Adv. 2019, 5, eaax0396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Child, K.M.; Herrick, D.B.; Schwob, J.E.; Holbrook, E.H.; Jang, W. The neurodegenerative capacity of olfactory stem cells is not limitless: Implications for aging. J. Neurosci. 2018, 38, 6806–6824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dibattista, M.; Pifferi, S.; Menini, A.; Reisert, J. Alzheimer’s disease: What Can We Learn from the Peripheral Olfactory System? Front. Neurosci. 2020, 14, 440. [Google Scholar] [CrossRef]
- O’Leary, T.P.; Stover, K.R.; Mantolino, H.M.; Darvesh, H.; Brown, R.E. Intact olfactory memory in the 5xFAD mouse model of Alzheimer’s disease from 3 to 15 months of age. Behav. Brain Res. 2020, 393, 112731. [Google Scholar] [CrossRef] [PubMed]
- Prediger, R.D.; Schamne, M.G.; Sampaio, T.B.; Moreira, E.L.G.; Rial, D. Animal models of olfactory dysfunction in neurodegenerative diseases. Handb. Clin. Neurol. 2019, 164, 431–452. [Google Scholar]
- Senf, K.; Karius, J.; Stumm, R.; Neuhaus, E.M. Chemokine signaling is required for homeostatic and injury-induced neurogenesis in the olfactory epithelium. STEM CELLS 2021, 39, 617–635. [Google Scholar] [CrossRef] [PubMed]
- Bhatia-Dey, N.; Heinbockel, T. Endocannabinoid-mediated neuromodulation in the olfactory bulb: Functional and therapeutic significance. Int. J. Mol. Sci. 2020, 21, 2850. [Google Scholar] [CrossRef] [Green Version]
- Vonderwalde, I.; Azimi, A.; Rolvink, G.; Ahlfors, J.-E.; Shoichet, M.S.; Morshead, C.M. Transplantation of directly reprogrammed human neural precursor cells following stroke promotes synaptogenesis and functional recovery. Transl. Stroke Res. 2020, 11, 93–107. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhatia-Dey, N.; Heinbockel, T. The Olfactory System as Marker of Neurodegeneration in Aging, Neurological and Neuropsychiatric Disorders. Int. J. Environ. Res. Public Health 2021, 18, 6976. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18136976
Bhatia-Dey N, Heinbockel T. The Olfactory System as Marker of Neurodegeneration in Aging, Neurological and Neuropsychiatric Disorders. International Journal of Environmental Research and Public Health. 2021; 18(13):6976. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18136976
Chicago/Turabian StyleBhatia-Dey, Naina, and Thomas Heinbockel. 2021. "The Olfactory System as Marker of Neurodegeneration in Aging, Neurological and Neuropsychiatric Disorders" International Journal of Environmental Research and Public Health 18, no. 13: 6976. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18136976