
Influence of Zooplankton and Environmental Factors on Clear-Water Phase in Lake Paldang, South Korea


Abstract
:1. Introduction
2. Materials and Methods
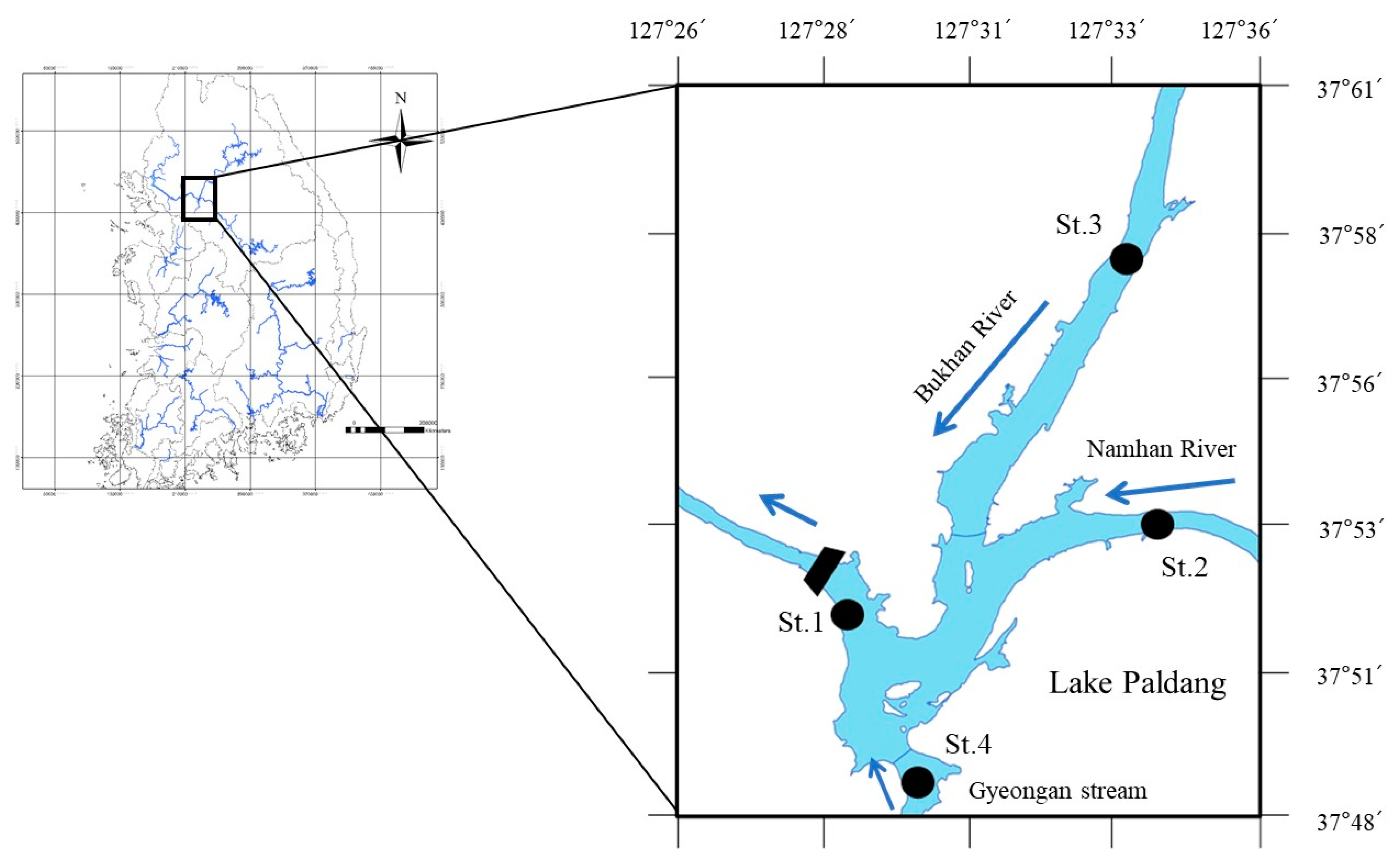
2.1. Study Sites and Period
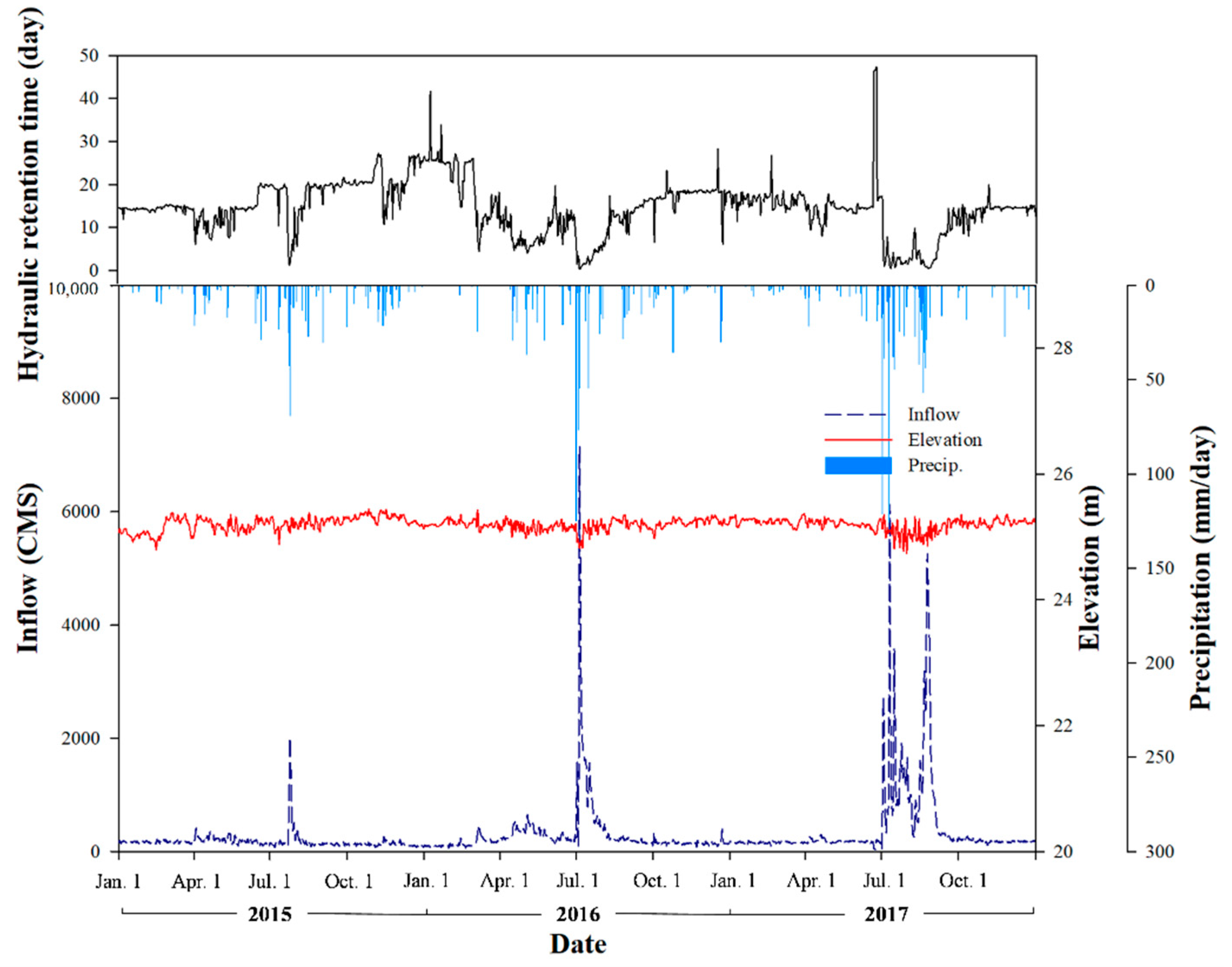
2.2. Precipitation and Hydrological Data
2.3. Sampling and Data Analysis
2.3.1. Physicochemical Factors
2.3.2. Biological Factors
2.3.3. CWP Occurrence Assessment
2.3.4. Statistical Analysis
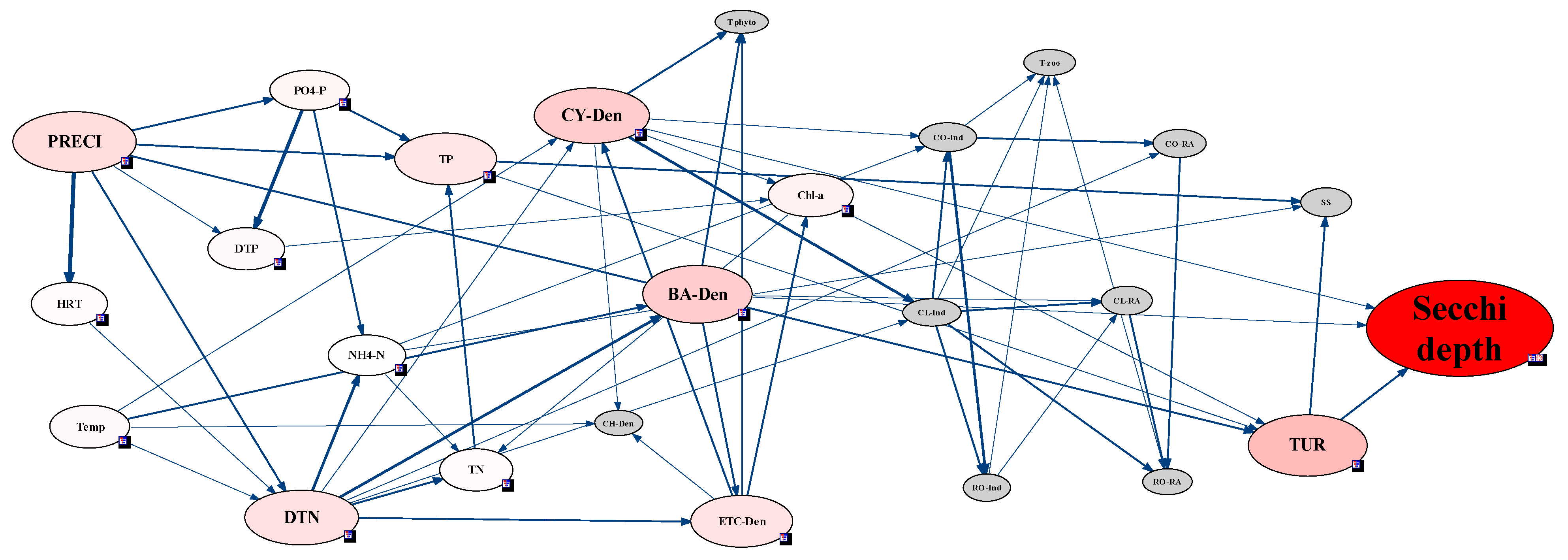
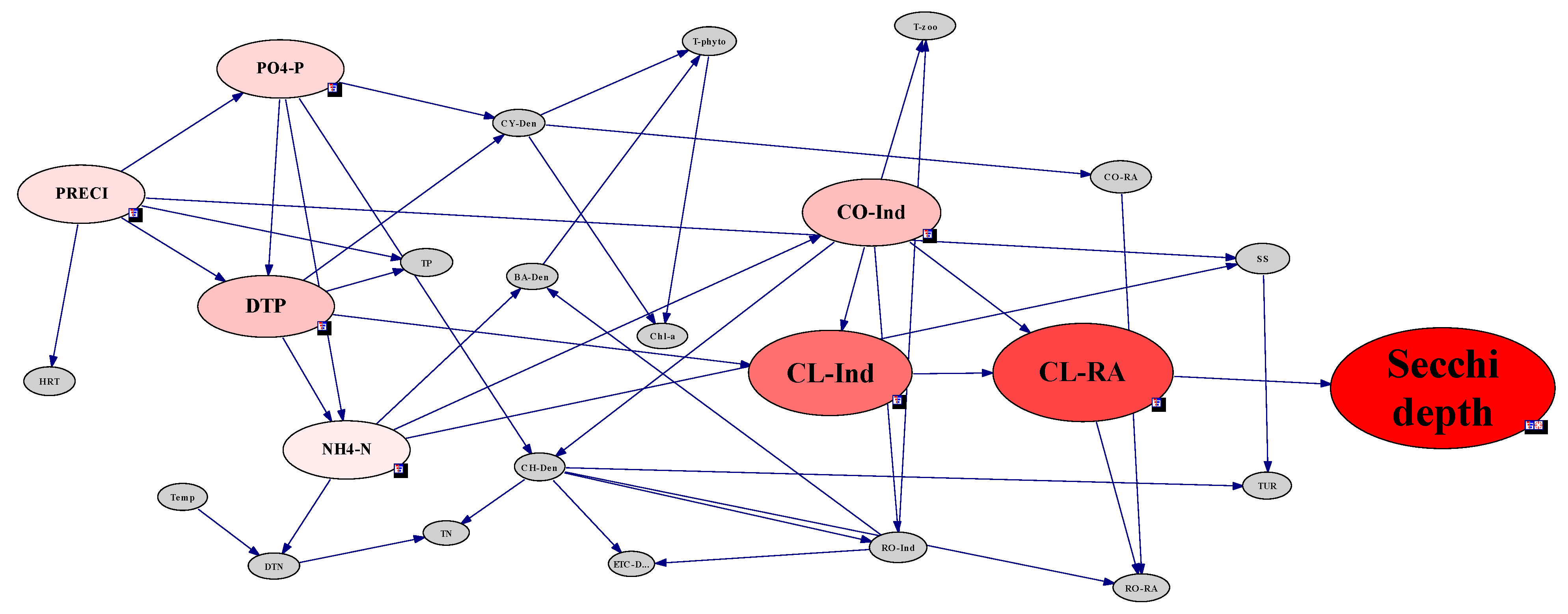
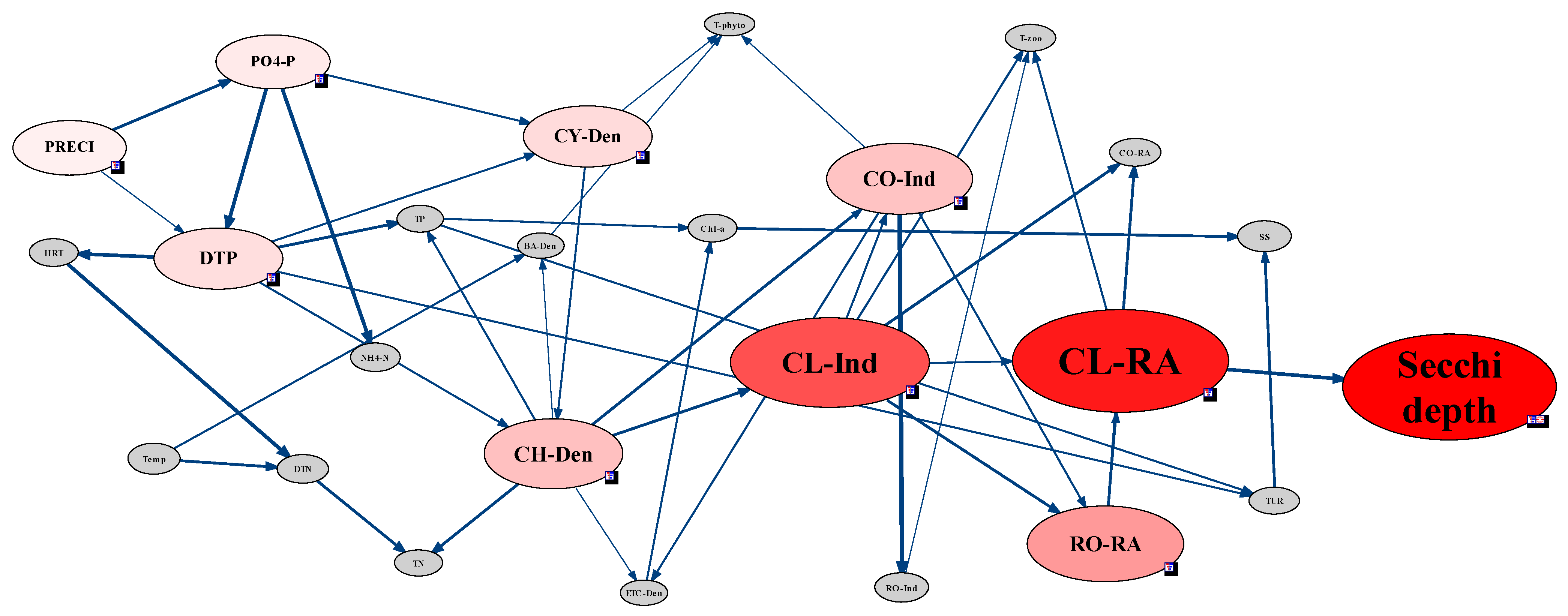
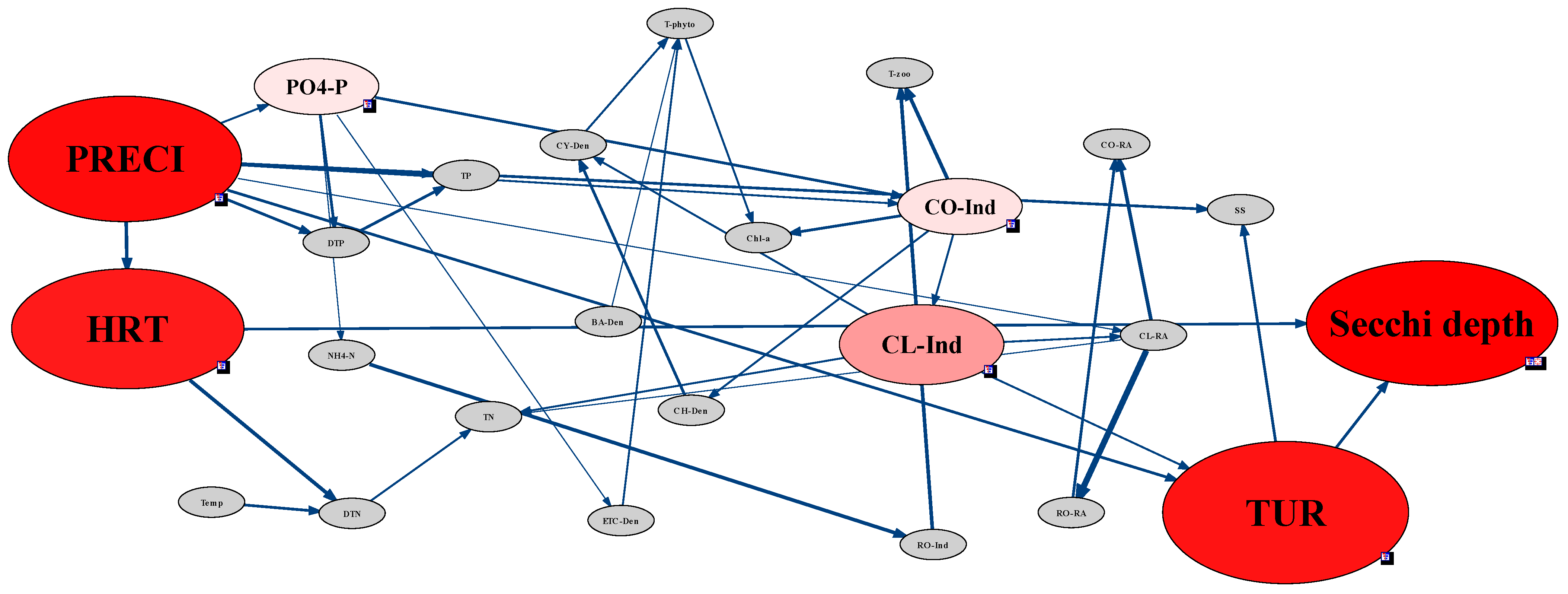
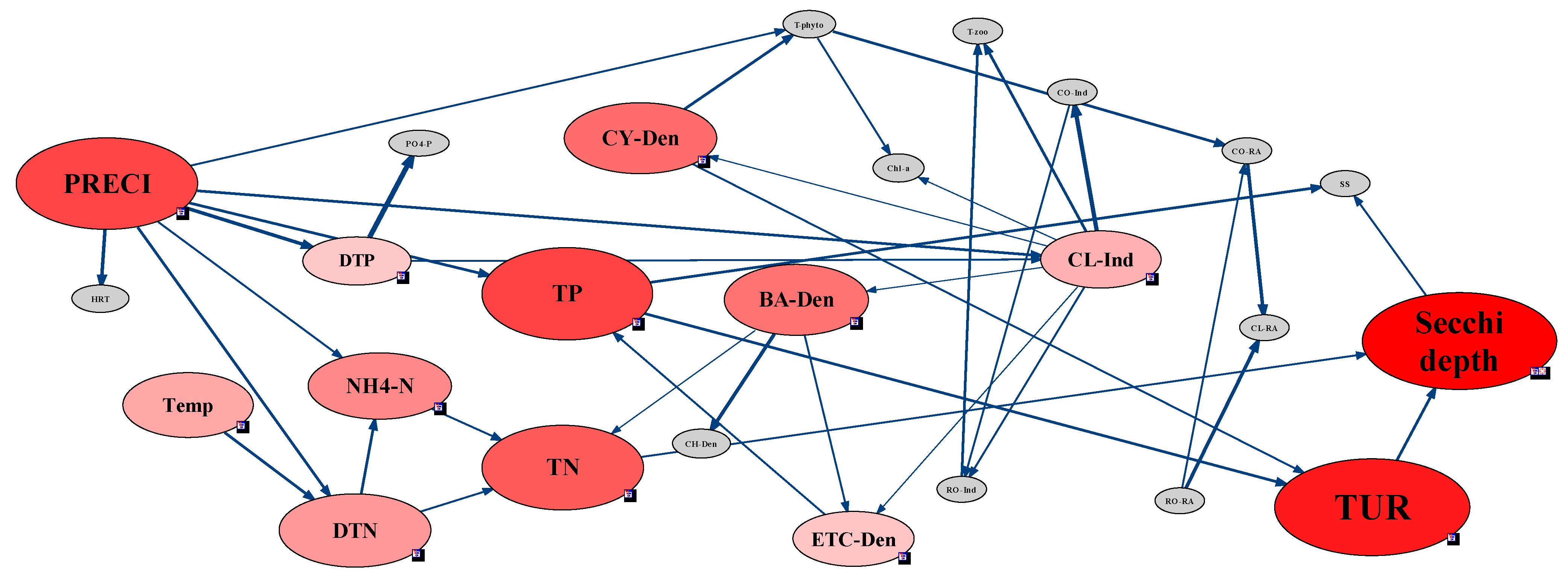
2.3.5. Analysis of Environmental Factors Affecting CWP Occurrence Using the Network Model
3. Results and Discussion
3.1. Precipitation and Hydrological Characteristics
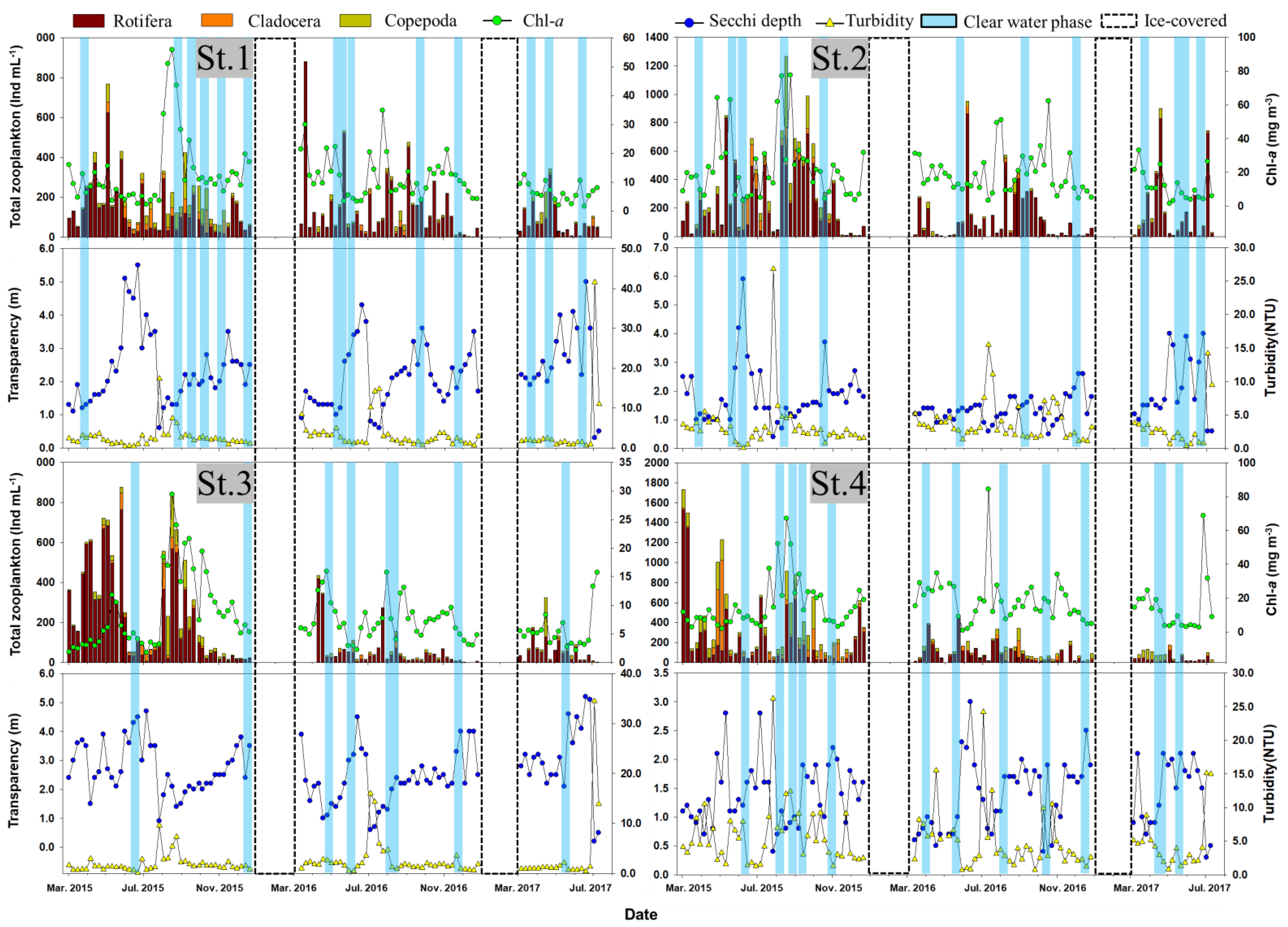
3.2. CWP Occurrence Pattern
3.3. Time Characteristics for CWP Occurrence
3.4. Analysis of CWP Occurrence Factors through the Network Model
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dröscher, I.; Finlay, K.; Patoine, A.; Leavitt, P.R. Daphnia control of the spring clear-water phase in six polymictic lakes of varying productivity and size. Int. Ver. Theor. Angew. Limnol. Verh. 2008, 30, 186–190. [Google Scholar] [CrossRef]
- Sommer, U.; Gliwicz, Z.M.; Lampert, W.; Duncan, A. The PEG-model of seasonal succession of planktonic events in fresh waters. Arch. Hydrobiol. 1986, 106, 433–471. [Google Scholar]
- George, D.G.; Talling, J.F.; Rigg, E. Factors influencing the temporal coherence of five lakes in the English Lake District. Freshw. Biol. 2000, 43, 449–461. [Google Scholar] [CrossRef]
- Lund, J. Studies on Asterionella formosa hass: II. Nutrient depletion and the spring maximum. J. Ecol. 1950, 38, 15–35. [Google Scholar] [CrossRef]
- Talling, J. The underwater light climate as a controlling factor in the production ecology of freshwater phytoplankton: With 14 figures in the text and on 1 folder. Int. Ver. Theor. Angew. Limnol. Mitt. 1971, 19, 214–243. [Google Scholar] [CrossRef]
- Talling, J.F. Comparative seasonal changes, and inter-annual variability and stability, in a 26-year record of total phytoplankton biomass in four English lake basins. Hydrobiologia 1993, 268, 65–98. [Google Scholar] [CrossRef]
- Maberly, S.; Hurley, M.; Butterwick, C.; Corry, J.; Heaney, S.; Irish, A.; Jaworski, G.; Lund, J.; Reynolds, C.; Roscoe, J. The rise and fall of Asterionella formosa in the South Basin of Windermere: Analysis of a 45-year series of data. Freshw. Biol. 1994, 31, 19–34. [Google Scholar] [CrossRef]
- George, D.G.; Hewitt, D.P.; Lund, J.W.G.; Smyly, W.J.P. The relative effects of enrichment and climate change on the long-term dynamics of Daphnia in Esthwaite Water, Cumbria. Freshw. Biol. 1990, 23, 55–70. [Google Scholar] [CrossRef]
- Lampert, W. Laboratory studies on zooplankton-cyanobacteria interactions. N. Zealand J. Mar. Freshw. Res. 1987, 21, 483–490. [Google Scholar] [CrossRef]
- Hambright, K.D.; Zohary, T.; Güde, H. Microzooplankton dominate carbon flow and nutrient cycling in a warm subtropical freshwater lake. Limnol. Oceanogr. 2007, 52, 1018–1025. [Google Scholar] [CrossRef]
- Talling, J. Phytoplankton—Zooplankton seasonal timing and the ‘clear-water phase’in some English lakes. Freshw. Biol. 2003, 48, 39–52. [Google Scholar] [CrossRef]
- Tailing, J. Freshwater phytoplankton—Accessible, microbial, influential population dynamics. Int. Ver. Theor. Angew. Limnol. Verh. 2002, 28, 7–28. [Google Scholar] [CrossRef]
- Lampert, W.; Fleckner, W.; Rai, H.; Taylor, B.E. Phytoplankton control by grazing zooplankton: A study on the spring clear-water phase. Limnol. Oceanogr. 1986, 31, 478–490. [Google Scholar] [CrossRef]
- Thornton, K.W.; Kimm, B.L.; Payne, F.E. Reservoir Limnology: Ecological Perspectives; Wiley & Sons: Hoboken, NJ, USA, 1990. [Google Scholar]
- Winston, W.; Criss, R. Geochemical variations during flash flooding, Meramec River basin, May 2000. J. Hydrol. 2002, 265, 149–163. [Google Scholar] [CrossRef]
- Kim, J.; Hong, W. Studies on the physical environmental factor analysis for water quality management in Man-made lake of Korea. J. Environ. Sci. Int. 1992, 1, 49–57. [Google Scholar]
- Kim, J.-M.; Park, J.-D.; Noh, H.-R.; Han, M.-S. Changes of seasonal and vertical water quality in Soyang and Paldang river-reservoir system, Korea. Korean J. Ecol. Environ. 2002, 35, 10–20. [Google Scholar]
- Kong, D.S. Limnological and ecological characteristics of a river-reservoir (Paldang), Korea. Korean J. Limnol. 1997, 30, 524–535. [Google Scholar]
- Park, H.-K.; Byeon, M.-S.; Kim, E.-K.; Lee, H.-J.; Chun, M.-J.; Jung, D.-I. Water quality and phytoplankton distribution pattern in upper inflow rivers of Lake Paldang. J. Environ. Sci. Int. 2004, 20, 615–624. [Google Scholar]
- Deevey, G.B. The zooplankton of the surface waters of the Delaware Bay region. Bull. Bingham Oceanogr. Collect. 1960, 17, 5–53. [Google Scholar]
- Sługocki, Ł.; Czerniawski, R.; Kowalska-Góralska, M.; Senze, M.; Reis, A.; Carrola, J.S.; Teixeira, C.A. The Impact of Land Use Transformations on Zooplankton Communities in a Small Mountain River (The Corgo River, Northern Portugal). Int. J. Environ. Res. Public Health 2018, 16, 20. [Google Scholar] [CrossRef] [Green Version]
- MOE. Standard Methods for the Examination Water Quality; The Korean Ministry of Environment: Sejong, Korea, 2014.
- Segers, H. Rotifera 2. The Lecanidae (Monogononta). Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; SPB Academic: The hague, The Netherlands, 1995. [Google Scholar]
- Segers, H. World Records of Lecanidae (Rotifera, Monogononta); Studiedocumenten van het KBIN Documents de Travail de l’IR Sc. NB; Koninklijk Belgisch Instituut voor Natuurwetenschappen: Brussels, Belgium, 1995. [Google Scholar]
- Chang, C.; Min, G. Key to the Korean Freshwater Cyclopoid Copepods and Their DNA Taxonomy; Junghaeng-Sa Publishing, Co.: Seoul, Korea, 2005; pp. 1–153. [Google Scholar]
- Jeong, H.; Kotov, A.; Lee, W. Checklist of the freshwater Cladocera (Crustacea: Branchiopoda) of South Korea. Proc. Boil. Soc. Wash. 2014, 127, 216–228. [Google Scholar] [CrossRef]
- John, D.; Whitton, B.; Brook, A. The freshwater algal flora of the British Isles. An identification guide to freshwater and terrestrial algae. Lond. Nat. 2002, 81, 234–235. [Google Scholar]
- Wehr, J.D.; Sheath, R.G.; Kociolek, J.P. Freshwater Algae of North America: Ecology and Classification; Academic Press: San Diego, CA, USA, 2015. [Google Scholar]
- BayesFusion, LLC. GeNIe Modeler; BayesFusion, LLC: Pittsburgh, PA, USA; Available online: https://support.bayesfusion.com/docs/GeNIe.pdf (accessed on 5 November 2020).
- Dodson, S.I. Introduction to limnology. J. N. Am. Benthol. Soc. 2004, 23, 661–662. [Google Scholar] [CrossRef]
- Deneke, R.; Nixdorf, B. On the occurrence of clear-water phases in relation to shallowness and trophic state: A comparative study. Hydrobiologia 1999, 408/409, 251–262. [Google Scholar] [CrossRef]
- Dröscher, I.; Patoine, A.; Finlay, K.; Leavitt, P. Climate control of the spring clear-water phase through the transfer of energy and mass to lakes. Limnol. Oceanogr. 2009, 54, 2469–2480. [Google Scholar] [CrossRef]
- Keckeis, S.; Baranyi, C.; Hein, T.; Holarek, C.; Riedler, P.; Schiemer, F. The significance of zooplankton grazing in a floodplain system of the River Danube. J. Plankton Res. 2003, 25, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Baranyi, C.; Hein, T.; Holarek, C.; Keckeis, S.; Schiemer, F. Zooplankton biomass and community structure in a Danube River floodplain system: Effects of hydrology. Freshw. Biol. 2002, 47, 473–482. [Google Scholar] [CrossRef]
- Muylaert, K.; Vyverman, W. Impact of a Flood Event on the Planktonic Food Web of the Schelde Estuary (Belgium) in Spring 1998. Hydrobiologia 2006, 559, 385–394. [Google Scholar] [CrossRef]
- Işkın, U.; Filiz, N.; Cao, Y.; Neif, É.M.; Öğlü, B.; Lauridsen, T.L.; Davidson, T.A.; Søndergaard, M.; Tavşanoğlu, Ü.N.; Beklioğlu, M.; et al. Impact of Nutrients, Temperatures, and a Heat Wave on Zooplankton Community Structure: An Experimental Approach. Water 2020, 12, 3416. [Google Scholar] [CrossRef]
- Arfè, A.; Quatto, P.; Zambon, A.; MacIsaac, H.J.; Manca, M. Long-Term Changes in the Zooplankton Community of Lake Maggiore in Response to Multiple Stressors: A Functional Principal Components Analysis. Water 2019, 11, 962. [Google Scholar] [CrossRef] [Green Version]
- You, K.-A.; Byeon, M.-S.; Hwang, S.-J. Effects of hydraulic-hydrological changes by monsoon climate on the zooplankton community in lake Paldang, Korea. Korean J. Ecol. Environ. 2012, 45, 278–288. [Google Scholar]
- Sim, Y.-B.; Jeong, H.-G.; Im, J.-K.; Youn, S.-J.; Byun, M.-S.; Yoo, S.-J. Spatial and Temporal Distribution of Zooplankton Communities in Lake Paldang. Korean J. Ecol. Environ. 2018, 51, 287–298. [Google Scholar] [CrossRef]
- Elser, J.J.; Goldman, C.R. Experimental separation of the direct and indirect effects of herbivorous zooplankton on phytoplankton in a subalpine lake. Int. Ver. Theor. Angew. Limnol. Verh. 1990, 24, 493–498. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Jensen, C.; Faafeng, B.; Hessen, D.O.; Søndergaard, M.; Lauridsen, T.; Brettum, P.; Christoffersen, K. The Impact of Nutrient State and Lake Depth on Top-down Control in the Pelagic Zone of Lakes: A Study of 466 Lakes from the Temperate Zone to the Arctic. Ecosystems 2003, 6, 313–325. [Google Scholar] [CrossRef]
- Matsuzaki, S.S.; Lathrop, R.C.; Carpenter, S.R.; Walsh, J.R.; Zanden, M.J.V.; Gahler, M.R.; Stanley, E.H. Climate and food web effects on the spring clear-water phase in two north-temperate eutrophic lakes. Limnol. Oceanogr. 2021, 66, 30–46. [Google Scholar] [CrossRef]
- Winder, M.; Schindler, D.E. Climatic effects on the phenology of lake processes. Glob. Chang. Biol. 2004, 10, 1844–1856. [Google Scholar] [CrossRef] [Green Version]
- Thackeray, S.J.; Henrys, P.A.; Feuchtmayr, H.; Jones, I.D.; Maberly, S.C.; Winfield, I.J. Food web de-synchronization in E ngland’s largest lake: An assessment based on multiple phenological metrics. Glob. Change Biol. 2013, 19, 3568–3580. [Google Scholar] [CrossRef] [Green Version]
- Luecke, C.; Vanni, M.J.; Magnuson, J.J.; Kitchell, J.F.; Jacobson, P.T. Seasonal regulation of Daphnia populations by planktivorous fish: Implications for the spring clear-water phase. Limnol. Oceanogr. 1990, 35, 1718–1733. [Google Scholar] [CrossRef]
- Conde-Porcuna, J.M.; Baquero, R.M.; Cruz-Pizarro, L. Effects of Daphnia longispina on rotifer populations in a natural environment: Relative importance of food limitation and interference competition. J. Plankton Res. 1994, 16, 691–706. [Google Scholar] [CrossRef]
- Fussmann, G. The importance of crustacean zooplankton in structuring rotifer and phytoplankton communities; an enclosure study. J. Plankton Res. 1996, 18, 1897–1915. [Google Scholar] [CrossRef] [Green Version]
- Lehman, J.T.; Platte, R.A.; Ferris, J.A. Role of hydrology in development of a vernal clear water phase in an urban impoundment. Freshw. Biol. 2007, 52, 1773–1781. [Google Scholar] [CrossRef]
- Berger, S.A.; Diehl, S.; Stibor, H.; Trommer, G.; Ruhenstroth, M. Water temperature and stratification depth independently shift cardinal events during plankton spring succession. Glob. Chang. Biol. 2010, 16, 1954–1965. [Google Scholar] [CrossRef]
- Abrantes, N.; Antunes, S.; Pereira, M.; Gonçalves, F. Seasonal succession of cladocerans and phytoplankton and their interactions in a shallow eutrophic lake (Lake Vela, Portugal). Acta Oecologica 2006, 29, 54–64. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temporal Tier 1 | Temporal Tier 2 | Temporal Tier 3 | Temporal Tier 4 | Temporal Tier 5 | Temporal Tier 6 |
|---|---|---|---|---|---|
| PRECI | HRT | CY | T-phyto-Den. | TN | SS |
| Temp | NH4–N | BA | T-zoo-Ind. | TP | SD |
| PO4–P | CH | RO-RA | Chl-a | TUR | |
| DTN | ETC-Den | CL-RA | |||
| DTP | RO-Ind | CO-RA | |||
| CL-Ind | |||||
| CO-Ind |
| Parameter (Unit) | Site | Before the CWP | After the CWP | t | p |
|---|---|---|---|---|---|
| Secchi depth (m) | St. 1 | 1.9 ± 0.5 | 2.6 ± 0.9 | −3.763 | 0.00271 * |
| St. 2 | 1.7 ± 1.0 | 2.7 ± 1.5 | −3.969 | 0.00265 * | |
| St. 3 | 2.4 ± 1.1 | 3.4 ± 1.1 | −3.417 | 0.01419 * | |
| St. 4 | 1.0 ± 0.4 | 1.7 ± 0.5 | −5.078 | 0.00021 * | |
| Turbidity (NTU) | St. 1 | 2.7 ± 1.4 | 1.7 ± 0.6 | 3.204 | 0.00758 * |
| St. 2 | 2.6 ± 1.2 | 1.6 ± 1.3 | 4.468 | 0.00120 * | |
| St. 3 | 2.4 ± 1.4 | 0.8 ± 0.6 | 3.234 | 0.01782 * | |
| St. 4 | 6.3 ± 2.9 | 3.6 ± 2.5 | 4.873 | 0.00030 * | |
| Total zooplankton (Ind L−1) | St. 1 | 96.4 ± 67.5 | 196.6 ± 138.0 | −5.838 | 0.00008 * |
| St. 2 | 166.9 ± 210.3 | 321.4 ± 347.7 | −2.410 | 0.03671 * | |
| St. 3 | 36.3 ± 22.7 | 72.0 ± 53.9 | −1.797 | 0.12243 | |
| St. 4 | 128.7 ± 152.2 | 288.5 ± 308.8 | −2.846 | 0.01377 * | |
| Chlorophyll-a (mg m−3) | St. 1 | 15.7 ± 10.0 | 9.2 ± 7.2 | 4.293 | 0.00105 * |
| St. 2 | 27.3 ± 22.8 | 10.3 ± 8.1 | 3.049 | 0.01227 * | |
| St. 3 | 9.0 ± 4.7 | 4.9 ± 2.6 | 2.842 | 0.02948 * | |
| St. 4 | 23.2 ± 14.9 | 11.5 ± 7.1 | 4.409 | 0.00071 * | |
| Total nitrogen (mg L−1) | St. 1 | 1.898 ± 0.470 | 1.799 ± 0.385 | 2.335 | 0.03774 * |
| St. 2 | 1.997 ± 0.456 | 1.963 ± 0.448 | 0.444 | 0.66658 | |
| St. 3 | 1.805 ± 0.285 | 1.818 ± 0.339 | −0.256 | 0.80627 | |
| St. 4 | 2.483 ± 1.073 | 2.133 ± 0.851 | 3.725 | 0.00255 * | |
| Total phosphorus (mg L−1) | St. 1 | 0.021 ± 0.008 | 0.016 ± 0.004 | 2.759 | 0.01733 * |
| St. 2 | 0.031 ± 0.018 | 0.026 ± 0.014 | 2.159 | 0.05617 | |
| St. 3 | 0.014 ± 0.007 | 0.012 ± 0.003 | 1.451 | 0.19707 | |
| St. 4 | 0.031 ± 0.010 | 0.023 ± 0.009 | 3.934 | 0.00171 * | |
| Hydraulic retention time (d) | St. 1 | 15.8 ± 4.7 | 16.9 ± 5.3 | −1.607 | 0.13396 |
| St. 2 | 13.9 ± 3.5 | 16.7 ± 6.3 | −1.652 | 0.12964 | |
| St. 3 | 12.2 ± 6.1 | 14.9 ± 6.5 | −2.447 | 0.04997 * | |
| St. 4 | 13.7 ± 4.9 | 14.4 ± 4.8 | −1.417 | 0.17995 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sim, Y.; Byeon, M.S.; Kim, K.; Yu, S.J.; Im, J.K. Influence of Zooplankton and Environmental Factors on Clear-Water Phase in Lake Paldang, South Korea. Int. J. Environ. Res. Public Health 2021, 18, 7205. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18137205
Sim Y, Byeon MS, Kim K, Yu SJ, Im JK. Influence of Zooplankton and Environmental Factors on Clear-Water Phase in Lake Paldang, South Korea. International Journal of Environmental Research and Public Health. 2021; 18(13):7205. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18137205
Chicago/Turabian StyleSim, Younbo, Myeong Seop Byeon, Keonhee Kim, Soon Ju Yu, and Jong Kwon Im. 2021. "Influence of Zooplankton and Environmental Factors on Clear-Water Phase in Lake Paldang, South Korea" International Journal of Environmental Research and Public Health 18, no. 13: 7205. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18137205