
Salt-Tolerant Compatible Microbial Inoculants Modulate Physio-Biochemical Responses Enhance Plant Growth, Zn Biofortification and Yield of Wheat Grown in Saline-Sodic Soil

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media, Chemical Reagents and Planting Materials
2.2. Salt-Tolerant Microbial Strains
2.3. In Vitro Screening for Salt Tolerance and Plant Growth-Promoting Traits
2.4. Mass Multiplication and Development of Bioformulations
2.5. Evaluation of Microbial Inoculants
Effect of Seed Bio-Priming on Seed Germination and Vigour Indices under Nethouse Conditions
2.6. Evaluation of Microbial Inoculants under Field Conditions
2.6.1. Experimental Set-Up
2.6.2. Estimation of Log CFU Count
2.6.3. Estimation of IAA in Rhizosphere Soil
2.6.4. Effect of Bioagents on Membrane Thermostability, Chlorophyll Content and Accumulation of Biomolecules and Organic Solutes
2.6.5. Effect of Bioagents on Lipid Peroxidation and Antioxidant Enzymes Activity
2.6.6. Gene Expression Analyses
2.6.7. Estimation of Zn Bio-Accumulation in Wheat
2.6.8. Plant Growth Promotion and Yield
2.6.9. Effect of Bioagents on Na+, K+ and Ca++ Uptake in Plant
2.7. Statistical Analyses
3. Results
3.1. In Vitro Screening for Salt Tolerance and Plant Growth-Promoting Traits
3.2. Effect of Seed Bio-Priming on Seed Germination and Vigour Indices
3.3. Estimation of Log CFU Count
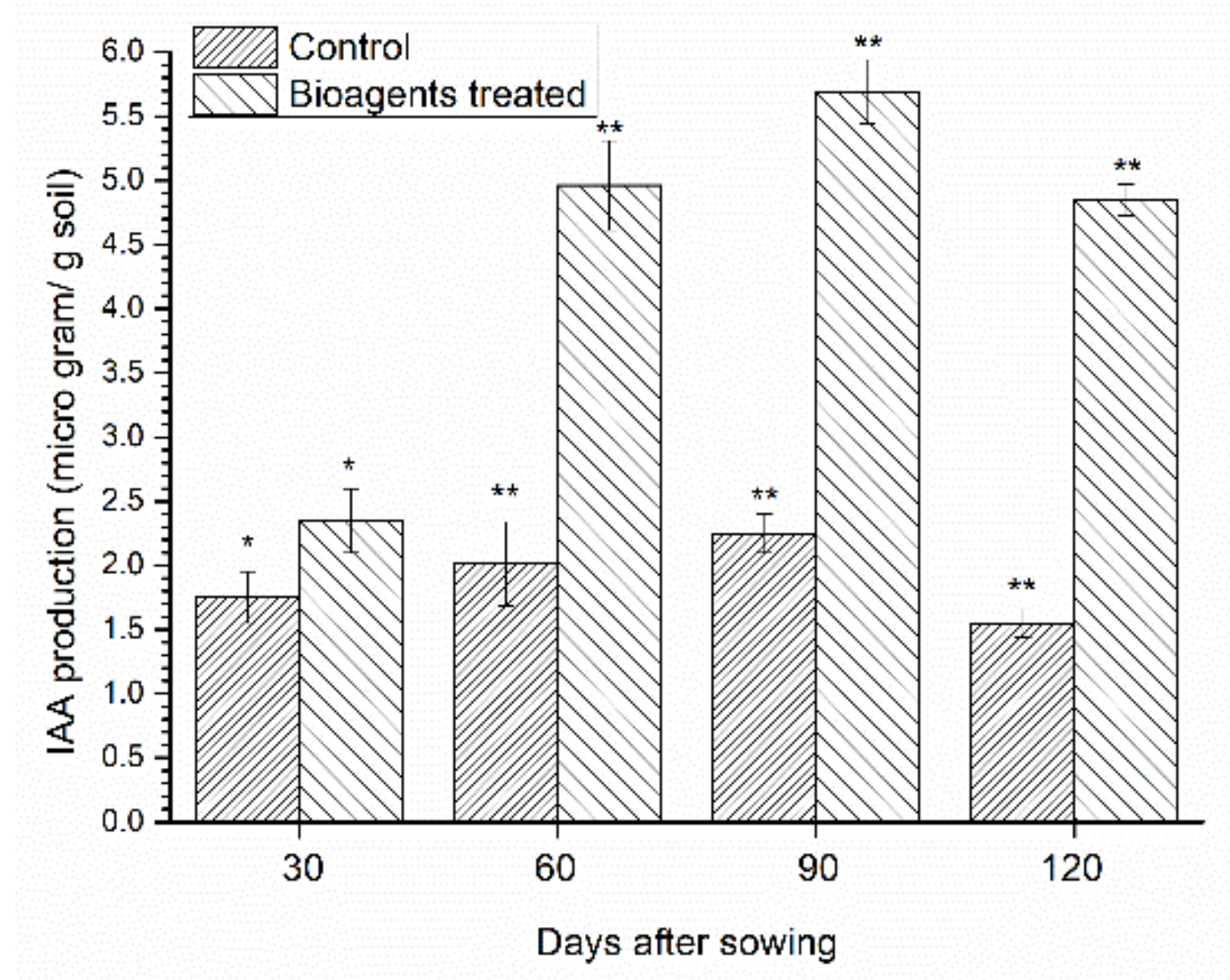
3.4. Estimation of IAA
3.5. Effect of Microbial Inoculants on Plant Growth Promotion
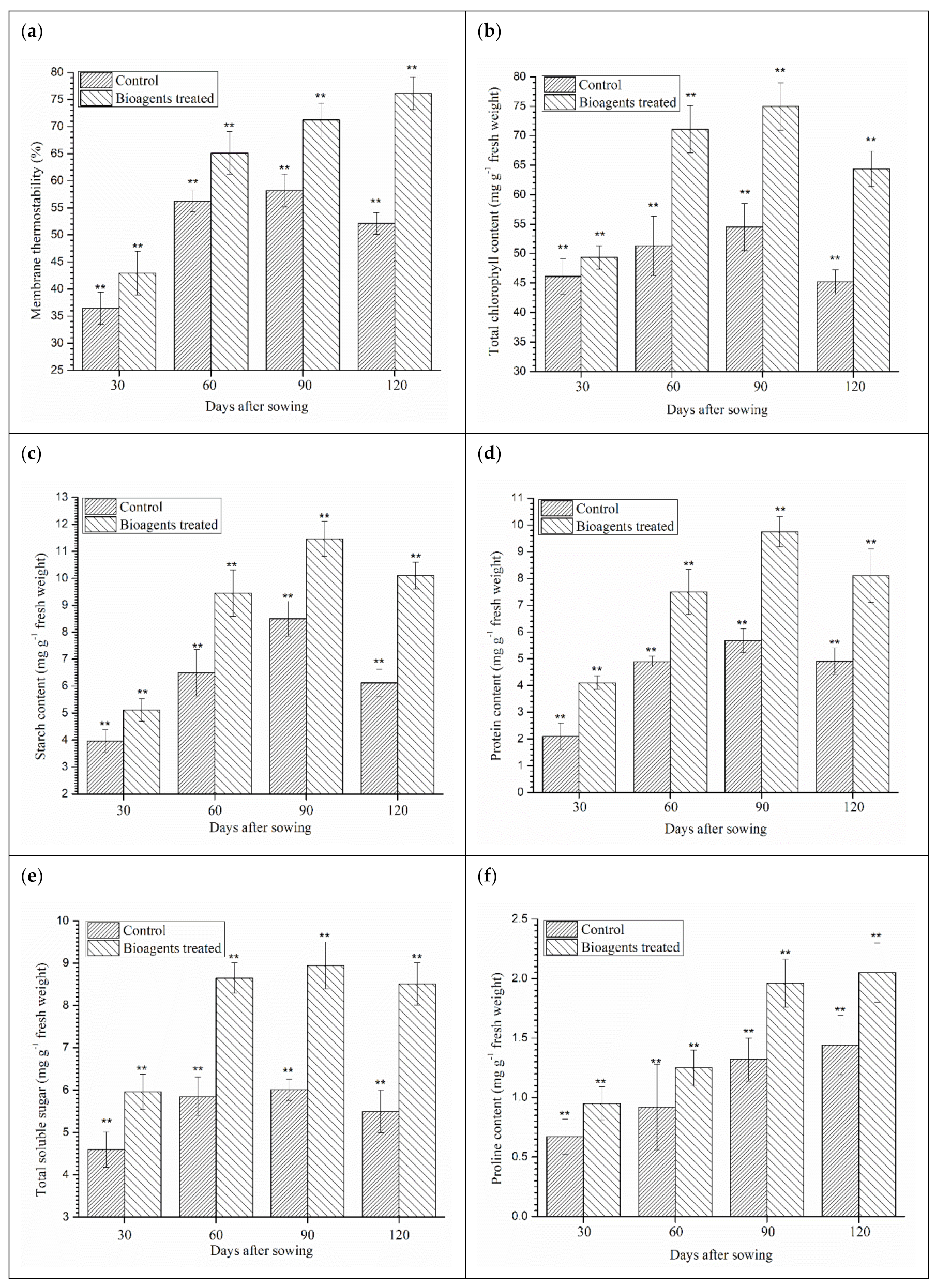
3.6. Effect of Microbial Inoculation on Membrane Thermostability, Chlorophyll Content, Accumulation of Biomolecules and Organic Solutes
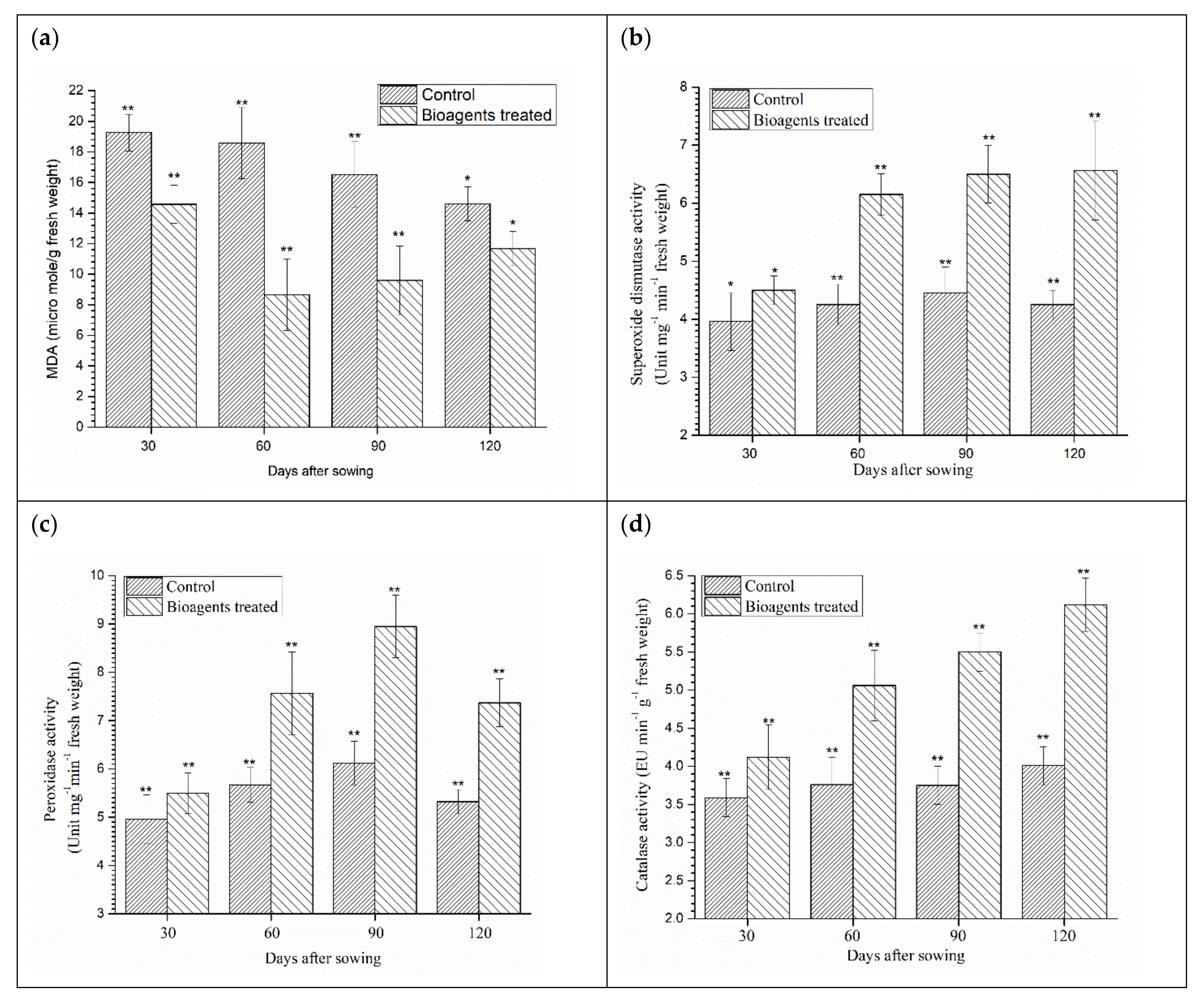
3.7. Effect of Microbial Inoculation on Lipid Peroxidation and Antioxidant Enzymes
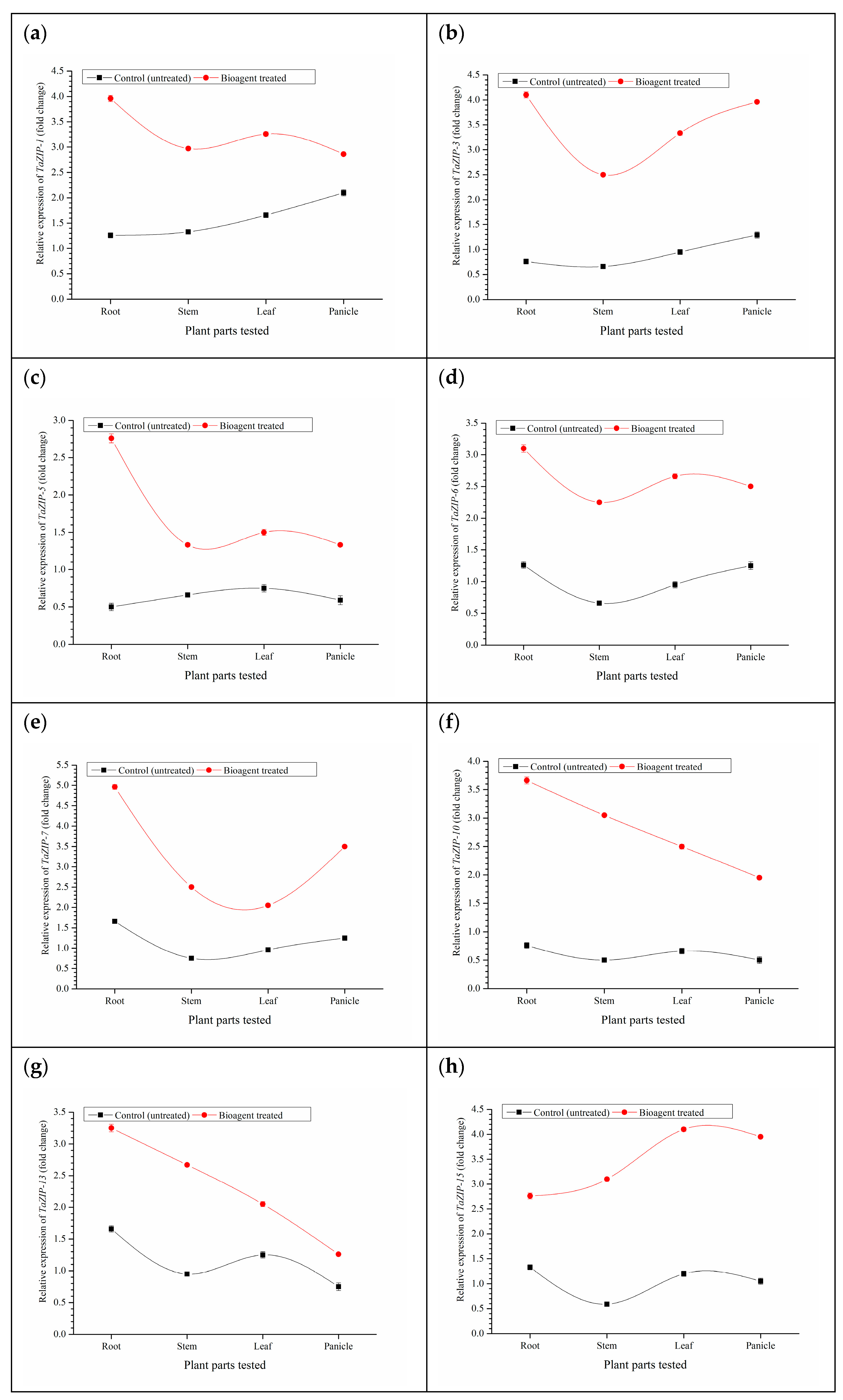
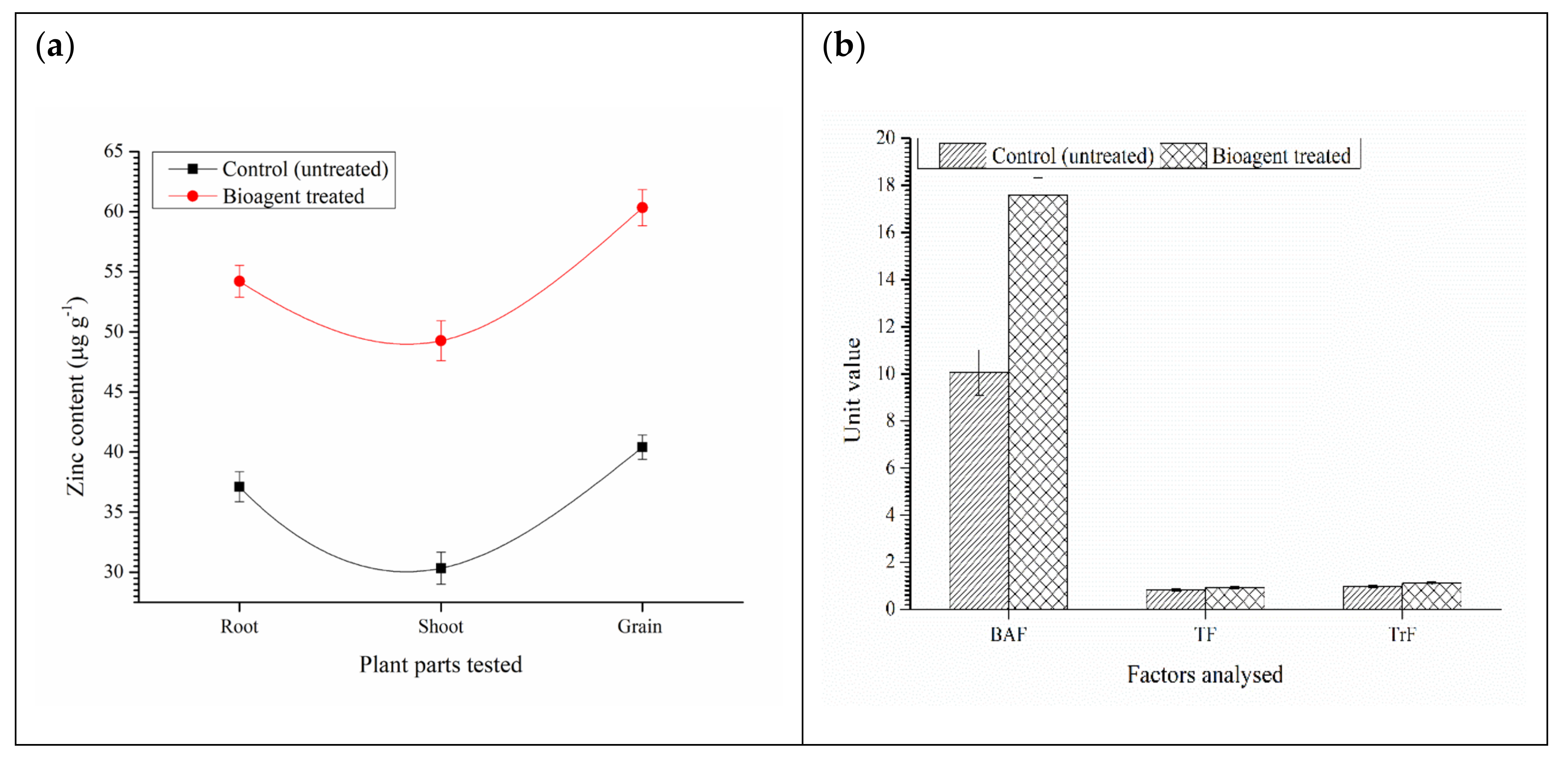
3.8. Expression Analyses of Zn Transporters and Biofortification of Wheat
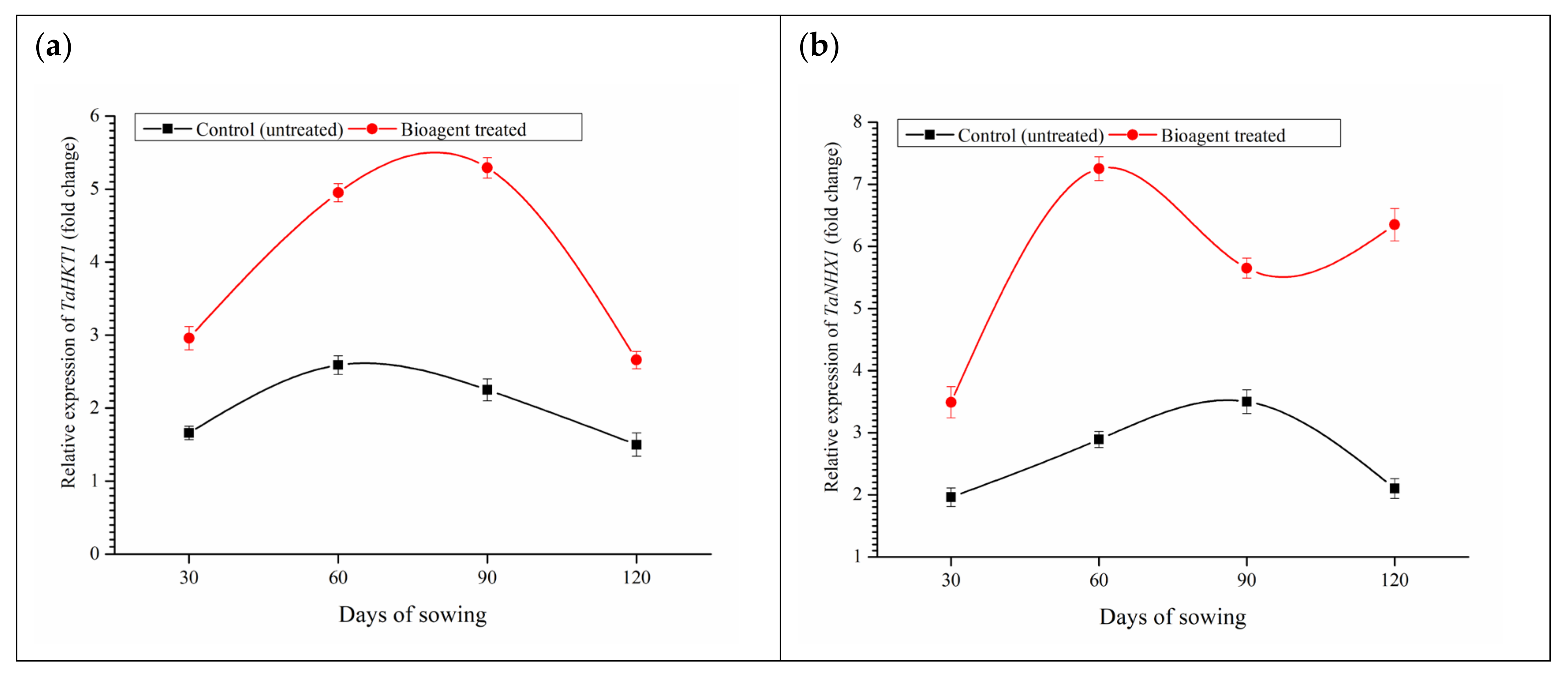
3.9. Effect of Microbial Inoculation on Expression of TaHKT-1 and TaNHX-1 and Uptake of Na+, K+ and Ca2+
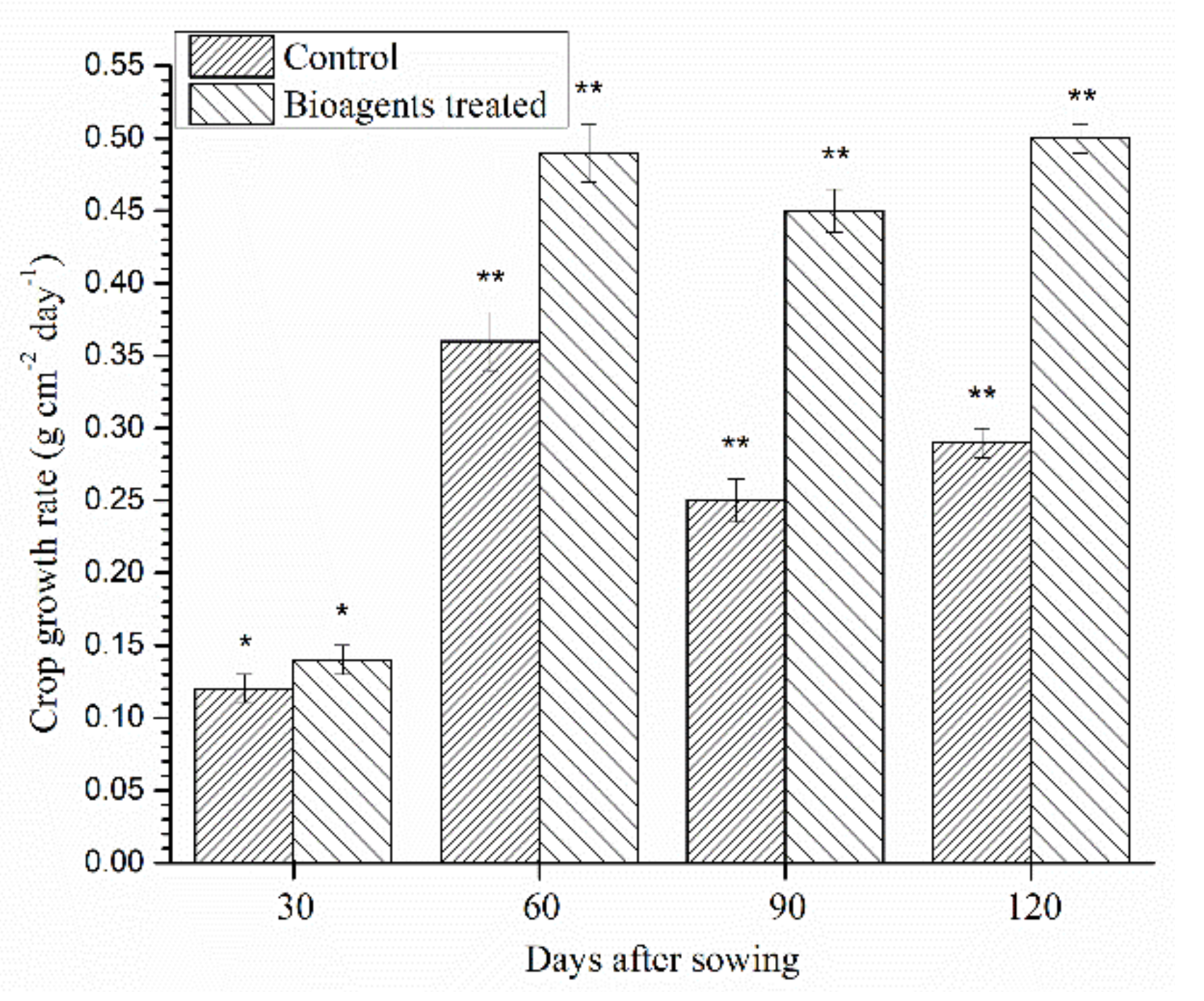
3.10. Effect of Microbial Inoculation on Yield and Yield Attributing Traits
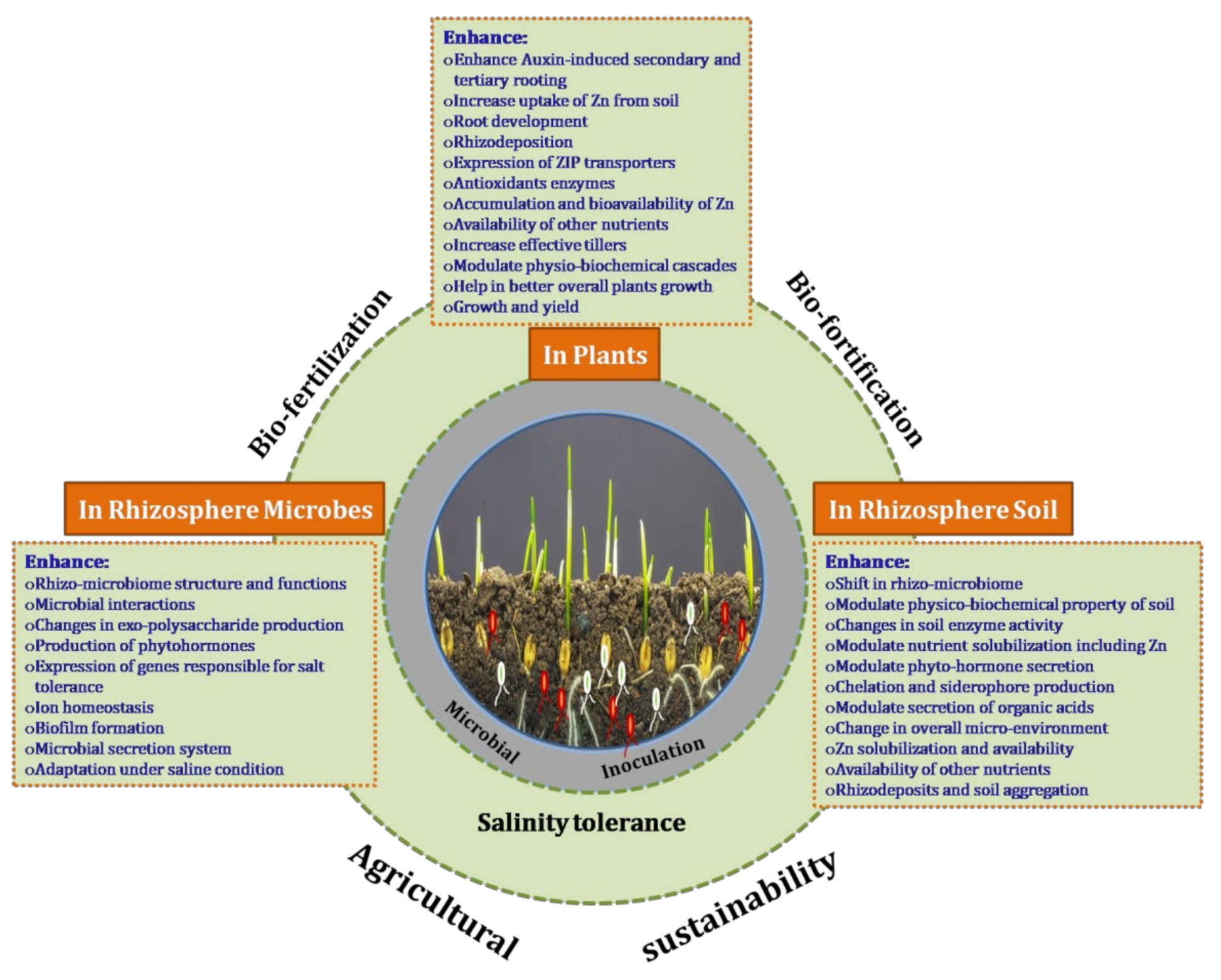
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flowers, T.J.; Garcia, A.; Koyama, M.; Yeo, A.R. Breeding for salt tolerance in crop plants—the role of molecular biology. Acta Physiol. 1997, 19, 427–433. [Google Scholar] [CrossRef]
- Szabolcs, I. Soils and salinisation. In Handbook of Plant and Crop Stress; Pessarakali, M., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1994; pp. 3–11. [Google Scholar]
- Das, D.K. Introductory Soil Science; Kalyani Publishers: New Delhi, India, 2009; pp. 194–214. [Google Scholar]
- National Remote Sensing Application and Associates. Mapping of Salt Affected Soils of India. 1:250000 Map Sheets, Legend; NRSA: Hyderabad, India, 1996. [Google Scholar]
- Zhang, S.; Gan, Y.; Xu, B. Application of plant-growth-promoting fungi Trichoderma longibrachiatum T6 enhances tolerance of wheat to salt stress through improvement of antioxidative defense system and gene expression. Front. Plant Sci. 2016, 7, 1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.H.; Kang, Y.H. Changes of soil microbial characteristics in saline-sodic soils under drip irrigation. J. Soil Sci. Plant Nutr. 2014, 14, 139–150. [Google Scholar] [CrossRef]
- Wong, V.N.; Greene, R.S.; Dalal, R.C.; Murphy, B.W. Soil carbon dynamics in saline and sodic soils: A review. Soil Use Manag. 2010, 26, 2–11. [Google Scholar] [CrossRef]
- Dargan, K.S.; Gaul, B.L. For paddy in alkali soil seedling age and planting date vital. Indian Farming 1974, 245, 11–12. [Google Scholar]
- Mishra, N.; Khan, S.S.; Sundari, S.K. Native isolate of Trichoderma: A biocontrol agent with unique stress tolerance properties. World J. Microbiol. Biotechnol. 2016, 32, 1–23. [Google Scholar] [CrossRef]
- Kumar, K.; Manigundan, K.; Amaresan, N. Influence of salt tolerant Trichoderma spp. on growth of maize (Zea mays L.) under different salinity conditions. J. Basic Microbiol. 2017, 56, 1–10. [Google Scholar]
- Liu, X.; Zhao, H.; Chen, S. Colonization of maize and rice plants by strain Bacillus megateriumC4. Curr. Microbiol. 2006, 52, 186–190. [Google Scholar] [CrossRef]
- Aravind, P.; Prasad, M.N.V. Zinc protects chloroplasts and associated photochemical functions in cadmium exposed Ceratophyllumdemersum L., a fresh water macrophyte. Plant Sci. 2004, 166, 1321–1327. [Google Scholar] [CrossRef]
- Ramesh, A.; Sharma, S.K.; Sharma, M.P.; Yadav, N.; Joshi, O.P. Inoculation of zinc solubilizing Bacillus aryabhattai strains for improved growth, mobilization and biofortification of zinc in soybean and wheat cultivated in Vertisols of central India. Appl. Soil Ecol. 2014, 73, 87–96. [Google Scholar] [CrossRef]
- Gunes, A.; Inal, A.; Alpaslan, M. Effect of salinity on stomatal resistance, proline and mineral composition of pepper. J. Plant Nutr. 1996, 19, 389–396. [Google Scholar] [CrossRef]
- Jamalomidi, M.; Esfahani, M.; Carapetian, J. Zinc and salinity interaction on agronomical traits, chlorophyll and proline content in lowland rice (Oryza sativa L.) genotypes. Pak. J. Biol. Sci. 2006, 9, 1315–1319. [Google Scholar] [CrossRef] [Green Version]
- Khoshgoftarmanesh, A.H.; Shariatmadari, H.; Karimian, N.; Khajehpour, M.R. Responses of wheat genotypes to zinc fertilization under saline soil conditions. J. Plant Nutr. 2006, 29, 1543–1556. [Google Scholar] [CrossRef]
- Tinker, P.B.; Lauchli, A. Advances in Plant Nutrition; Academic Publishers: San Diego, CA, USA, 1984. [Google Scholar]
- Genc, Y.; McDonald, G.K.; Graham, R.D. The interactive effects of zinc and salt on growth of wheat. In Plant Nutrition for Food Security, Human Health and Environmental Protection; Li, C.J., Zhang, F.-S., Doberman, A., Hinsinger, P., Lambers, H., Li, X.L., Marschner, P., Maene, L., McGrath, S.P., Oenema, O., et al., Eds.; Tsinghua University Press: Beijing, China, 2005; pp. 548–549. [Google Scholar]
- Alpaslan, M.; Inal, A.; Gunes, A.; Çikili, Y.; Ozcan, H. Effect of zinc treatment on the alleviation of sodium and chloride injury in tomato (Lycopersicum esculentum (L.) Mill. cv. Lale) grown under salinity. Turk. J. Agric. For. 1999, 23, 1–6. [Google Scholar]
- Cakmak, I. Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef]
- Joshi, A.K.; Kumari, M.; Singh, V.P.; Reddy, C.M.; Kumar, S.; Rane, J.; Chand, R. Stay green trait: Variation, inheritance and its association with spot blotch resistance in spring wheat (Triticum aestivum L.). Euphytica 2007, 153, 59–71. [Google Scholar] [CrossRef]
- Joshi, A.K.; Mishra, B.; Chatrath, R.; Ferrara, G.O.; Singh, R.P. Wheat improvement in India: Present status, emerging challenges and future prospects. Euphytica 2007, 157, 431–446. [Google Scholar] [CrossRef]
- Joshi, A.K.; Chand, R. Progress of researches done to understand host-pathogen relationship for spot blotch pathogen of wheat. J. Wheat Res. 2011, 3, 1–7. [Google Scholar]
- Singh, C.; Singh, P.; Singh, R. (Eds.) Modern Techniques of Raising Field Crops; Oxford & IBH Publishing CO. PVT. LTD.: New Delhi, India, 2008; pp. 355–362. [Google Scholar]
- Singh, R.P.; Jha, P.; Jha, P.N. The plant-growth-promoting bacterium Klebsiella sp. SBP-8 confers induced systemic tolerance in wheat (Triticum aestivum) under salt stress. J. Plant Physiol. 2015, 184, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.P.; Mishra, V.K.; Singh, S.; Sharma, D.K.; Singh, D.; Singh, U.S.; Singh, R.K.; Haefele, S.M.; Ismail, A.M. Productivity of sodic soils can be enhanced through the use of salt tolerant rice varieties and proper agronomic practices. Field Crops Res. 2016, 190, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Ebel, C.; BenFeki, A.; Hanin, M.; Solano, R.; Chini, A. Characterization of wheat (Triticum aestivum) TIFY family and role of Triticum Durum TdTIFY11a in salt stress tolerance. PLoS ONE 2018, 13, 0200566. [Google Scholar] [CrossRef] [Green Version]
- Abhinandan, K.; Skori, L.; Stanic, M.; Hickerson, N.M.N.; Jamshed, M.; Samuel, M.A. Abiotic stress signaling in wheat–an inclusive overview of hormonal interactions during abiotic stress responses in wheat. Front. Plant Sci. 2018, 9, 734. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Wasiullah; Singh, S.; Imran, M.; Pathak, N.; Alam, M.; Rai, J.P.; Singh, R.K.; Sarma, B.K.; et al. Compatible salt-tolerant rhizosphere microbe-mediated induction of phenylpropanoid cascade and induced systemic responses against Bipolaris sorokiniana (Sacc.) Shoemaker causing spot blotch disease in wheat (Triticum aestivum L.). Appl. Soil Ecol. 2016, 108, 300–306. [Google Scholar]
- Siddaiah, C.N.; Satyanarayana, N.R.; Mudili, V.; Gupta, V.K.; Gurunathan, S.; Rangappa, S.; Huntrike, S.S.; Srivastava, R.K. Elicitation of resistance and associated defense responses in Trichoderma hamatum induced protection against pearl millet downy mildew pathogen. Sci. Rep. 2017, 7, 43991. [Google Scholar] [CrossRef]
- Glick, B.R.; Cheng, Z.; Czarny, J.; Duan, J. Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur. J. Plant Pathol. 2007, 119, 329–339. [Google Scholar] [CrossRef]
- Orhan, F. Alleviation of salt stress by halotolerant and halophilic plant growth-promoting bacteria in wheat (Triticum aestivum). Braz. J. Microbiol. 2016, 47, 621–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Zhang, H.; Kim, M.S.; Sun, Y.; Dowd, S.E.; Shi, H.; Paré, P.W. Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol. Plant Microbe Interact. 2008, 21, 737–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, D.; Lade, H. Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: A review. Agron. Sustain. Dev. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Boil. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, U.B.; Trivedi, M.; Sahu, P.K.; Paul, S.; Paul, D.; Saxena, A.K. Seed biopriming with salt-tolerant endophytic Pseudomonas geniculata-modulated biochemical responses provide ecological fitness in maize (Zea mays L.) grown in saline sodic soil. Int. J. Environ. Res. Public Health 2019, 17, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Singh, U.B.; Trivdi, M.; Malviya, D.; Sahu, P.K.; Roy, M.; Sharma, P.K.; Singh, H.V.; Manna, M.C.; Saxena, A.K. Restructuring the cellular responses: Connecting microbial intervention with ecological fitness and adaptiveness to the maize (Zea mays L.) grown in saline–sodic soil. Front. Microbiol. 2021, 11, 568325. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Sugawara, M.; Okunou, K.; Nakayama, H.; Schroeder, J.I.; Shinmyo, A.; Yoshida, K. Functions of HKT transporters in sodium transport in roots and in protecting leaves from salinity stress. Plant Biotech. 2008, 25, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Brick, J.M.; Bostock, R.M.; Silversone, S.E. Rapid in situ assay for indole acetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Dey, R.K.; Pal, K.K.; Bhatt, D.M.; Chauhan, S.M. Growth promotion and yield enhancement of peanut (Arachis hypogaea L.) by application of plant growth-promoting rhizobacteria. Microbiol. Res. 2004, 159, 371–394. [Google Scholar] [CrossRef]
- Gaur, A.C. Physiological Functions of Phosphate Solubilizing Micro-Organisms. Phosphate Solubilizing Micro-Organisms as Biofertilizers; Omega Scientific Publishing: New Delhi, India, 1990; pp. 16–72. [Google Scholar]
- Boller, T.; Mauch, F. Colorimetric assay for chitinase. Methods Enzymol. 1988, 161, 430–435. [Google Scholar]
- Whitman, W.B.; Goodfellow, M.; Kämpfer, P.; Busse, H.-J.; Trujillo, M.E.; Ludwig, W.; Suzuki, K.-i. (Eds.) Bergey’s Manual of Systematic Bacteriology Parts A and B, 2nd ed.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Singh, U.B.; Malviya, D.; Wasiullah; Singh, S.; Pradhan, J.K.; Singh, B.P.; Roy, M.; Imram, M.; Pathak, N.; Baisyal, B.M.; et al. Bio-protective microbial agents from rhizosphere eco-systems triggering plant defense responses provide protection against sheath blight disease in rice (Oryza sativa L.). Microbiol. Res. 2016, 192, 300–312. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Khan, W.; Singh, S.; Karthikeyan, N.; Imran, M.; Rai, J.P.; Sarma, B.K.; Manna, M.C.; Chaurasia, R.; et al. Earthworm grazed-Trichoderma harzianum biofortified spent mushroom substrates modulate accumulation of natural antioxidants and bio-fortification of mineral nutrients in tomato. Front. Plant Sci. 2018, 9, 1017. [Google Scholar] [CrossRef]
- ISTA. Seed Testing International, ISTA Secretariat, CH – Switzerland, ISTA News Bulletin No. 126, 2003. Available online: https://www.seedtest.org/upload/cms/user/STI126 (accessed on 11 August 2021).
- Udayashankar, A.C.; Nayaka, S.C.; Reddy, M.S.; Srinivas, C. Plant growth-promoting rhizobacteria mediate induced systemic resistance in rice against bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae. Biol. Control. 2011, 59, 114–122. [Google Scholar] [CrossRef]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor determination in soybean seed by multiple criteria 1. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Kharb, R.S.; Lather, B.S.; Deswal, D.P. Prediction of field emergence through heritability and genetic advance of vigour parameters. Seed Sci. Technol. 1994, 22, 461–466. [Google Scholar]
- Thimmaiah, S.R. Standard Methods of Biochemical Analysis; Kalyani Publishers: New Delhi, India, 2012; p. 545. [Google Scholar]
- Sadasivam, S. Biochemical Methods; New Age International: New Delhi, India, 1996; p. 256. [Google Scholar]
- Fokar, M.; Henry, T.N.; Blum, A. Heat tolerance in spring wheat I. Estimating cellular thermo-tolerance and its heritability. Euphytica 1998, 104, 1–8. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Mittal, S.B. Evaluation of chemical methods for estimating available zinc and response of green gram (Phaseolusaureus roxb) to applied zinc in non-calcareous soils. Plant Soil 1981, 63, 477–484. [Google Scholar] [CrossRef]
- Ma, L.Q.; Komar, K.M.; Tu, C.; Zhang, W.; Cai, Y.; Kenelly, E.D. A Fern that hyperaccumulates arsenic. Nature 2001, 409, 579–582. [Google Scholar] [CrossRef]
- Baker, A.J.M.; Brooks, R.R. Terrestrial higher plants which hyperaccumulate metal elements. A review of their distribution, ecology and phytochemistry. Biorecovery 1989, 1, 81–126. [Google Scholar]
- Watson, D.J. The physiological basis of variation in yield. Adv. Agron. 1952, 4, 101–105. [Google Scholar]
- The Food and Agriculture Organization of the United Nations (FAO). Crops and Drops: Making the Best Use of Water for Agriculture; FAO: Rome, Italy, 2002; Available online: http://www.fao.org/docrep/w5146e/w5146e0a.htm (accessed on 11 August 2021).
- Minasny, B.; Hong, S.Y.; Hartemink, A.E.; Kim, Y.H.; Kang, S.S. Soil pH increase under paddy in South Korea between 2000 and 2012. Agric. Ecosyst. Environ. 2016, 221, 205–213. [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; Nafady, N.A.; Bashandy, S.R.; Hassan, A.A. Mitigation of effect of salt stress on the nodulation, nitrogen fixation and growth of chickpea (Cicer arietinum L.) by triple microbial inoculation. Rhizosphere 2019, 10, 100148. [Google Scholar] [CrossRef]
- Ramadoss, D.; Lakkineni, V.K.; Bose, P.; Ali, S.; Annapurna, K. Mitigation of salt stress in wheat seedlings by halotolerant bacteria isolated from saline habitats. SpringerPlus 2013, 2, 6–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qurashi, A.W.; Sabri, A.N. Osmolyte accumulation in moderately halophilic bacteria improves salt tolerance of chickpea. Pak. J. Bot. 2013, 45, 1011–1016. [Google Scholar]
- Shrivastava, U.P.; Kumar, A. Characterization and optimization of 1-aminocyclopropane-1-carboxylate deaminase (ACCD) activity in different rhizospheric PGPR along with Microbacterium sp. strain ECI-12A. Int. J. Appl. Sci. Biotechnol. 2013, 1, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Essghaier, B.; Dhieb, C.; Rebib, H.; Ayari, S.; Boudabous, A.R.; Sadfi-Zouaoui, N. Antimicrobial behavior of intracellular proteins from two moderately halophilic bacteria: Strain J31 of Terribacillus halophilus and strain M3-23 of Virgibacillus marismortui. J. Plant Pathol. Microbiol. 2014, 5, 214. [Google Scholar] [CrossRef] [Green Version]
- Marakana, T.; Sharma, M.; Sangani, K. Isolation and characterization of halotolerant bacteria and it’s effects on wheat plant as PGPR. Pharma Innov. J. 2018, 7, 102–110. [Google Scholar]
- Ahkami, A.H.; White, R.A., III; Handakumbura, P.P.; Jansson, C. Rhizosphere engineering: Enhancing sustainable plant ecosystem productivity. Rhizosphere 2017, 3, 233–243. [Google Scholar] [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and fungal contributions to carbon sequestration in agro-ecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Wang, S.; Wu, H.; Qiao, J.; Ma, L.; Liu, J.; Xia, Y.; Gao, X. Molecular mechanism of plant growth promotion and induced systemic resistance to tobacco mosaic virus by Bacillus spp. J. Microbiol. Biotechnol. 2009, 19, 1250–1258. [Google Scholar] [CrossRef]
- Zhang, H.; Kim, M.S.; Krishnamachari, V.; Payton, P.; Sun, Y.; Grimson, M.; Farag, M.A.; Ryu, C.M.; Allen, R.; Melo, I.S.; et al. Rhizobacterial volatile emissions regulate auxin homeostasis and cell expansion in Arabidopsis. Planta 2007, 226, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D. Alleviation of salt stress by plant growth regulators and IAA producing bacteria in wheat. Acta Physiol. Plant. 2009, 31, 861–864. [Google Scholar] [CrossRef]
- Pietri, J.A.; Brookes, P.C. Relationships between soil pH and microbial properties in a UK arable soil. Soil Biol. Biochem. 2008, 40, 1856–1861. [Google Scholar] [CrossRef]
- Boiero, L.; Perrig, D.; Masciarelli, O.; Penna, C.; Cassan, F.; Luna, V. Phytohormone production by three strains of Bradyrhizobium japonicum and possible physiological and technological implications. Appl. Microbiol. Biotechnol. 2007, 74, 874–880. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Davranov, K.; Wirth, S.; Hashem, A.; Abd-Allah, E.F. Impact of soil salinity on the plant-growth–promoting and biological control abilities of root associated bacteria. Saudi J. Biol. Sci. 2017, 24, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.A.; Bakken, L.R. Viability of soil bacteria: Optimization of plate-counting technique and comparison between total counts and plate counts within different size groups. Microb. Ecol. 1987, 13, 59–74. [Google Scholar] [CrossRef]
- Singh, D.; Rajawat, M.V.S.; Kaushik, R.; Prasanna, R.; Saxena, A.K. Beneficial role of endophytes in biofortification of Zn in wheat genotypes varying in nutrient use efficiency grown in soils sufficient and deficient in Zn. Plant Soil 2017, 416, 107–116. [Google Scholar] [CrossRef]
- Velmourougane, K.; Prasanna, R.; Singh, S.; Chawla, G.; Kumar, A.; Saxena, A.K. Modulating rhizosphere colonization, plant growth, soil nutrient availability and plant defense enzyme activity through Trichoderma viride-Azotobacter chroococcum biofilm inoculation in chickpea. Plant Soil 2017, 421, 157–174. [Google Scholar] [CrossRef]
- Singh, D.; Geat, N.; Rajawat, M.V.S.; Prasanna, R.; Kar, A.; Singh, A.M.; Saxena, A.K. Prospecting endophytes from different Fe or Zn accumulating wheat genotypes for their influence as inoculants on plant growth, yield, and micronutrient content. Ann. Microbiol. 2018, 68, 815–833. [Google Scholar] [CrossRef]
- Singh, D.; Geat, N.; Rajawat, M.V.S.; Mahajan, M.M.; Prasanna, R.; Singh, S.; Kaushik, R.; Singh, R.N.; Kumar, K.; Saxena, A.K. Deciphering the Mechanisms of Endophyte-Mediated Biofortification of Fe and Zn in Wheat. J. Plant Growth Regul. 2018, 37, 174–182. [Google Scholar] [CrossRef]
- Lucas, J.A.; García-Cristobal, J.; Bonilla, A.; Ramos, B.; Gutierrez-Manero, J. Beneficial rhizobacteria from rice rhizosphere confers high protection against biotic and abiotic stress inducing systemic resistance in rice seedlings. Plant Physiol. Biochem. 2014, 82, 44–53. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, X.; Zhang, X.; Dong, L.; Zhang, J.; Wei, Y. Improved plant growth and Zn accumulation in grains of rice (Oryza sativa L.) by inoculation of endophytic microbes isolated from a Zn hyperaccumulator, Sedum alfredii H. J. Agric. Food Chem. 2014, 62, 1783–1791. [Google Scholar] [PubMed]
- White, P.J.; Broadley, M.R. Physiological limits to zinc biofortification of edible crops. Front. Plant Sci. 2011, 2, 80. [Google Scholar] [CrossRef] [Green Version]
- Naz, I.; Ahmad, H.; Khokhar, S.N.; Khan, K.; Shah, A.H. Impact of zinc solubilizing bacteria on zinc contents of wheat. Am. Euras. J. Agric. Environ. Sci. 2016, 16, 449–454. [Google Scholar]
- Yadav, R.C.; Sharma, S.K.; Ramesh, A.; Sharma, K.; Sharma, P.K.; Varma, A. Contribution of Zinc-Solubilizing and-Mobilizing Microorganisms (ZSMM) to enhance zinc bioavailability for better soil, plant, and human health. In Rhizosphere Microbes; Springer: Singapore, 2020; pp. 357–386. [Google Scholar]
- Yadav, R.; Ror, P.; Rathore, P.; Kumar, S.; Ramakrishna, W. Bacillus subtilis CP4, isolated from native soil in combination with arbuscular mycorrhizal fungi promotes biofortification, yield and metabolite production in wheat under field conditions. J. Appl. Microbiol. 2020, 131, 339–359. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotox. Environ. Safe. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Ilangumaran, G.; Smith, D.L. Plant growth promoting rhizobacteria in amelioration of salinity stress: A systems biology perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef] [PubMed]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Murzello, C.; Sun, Y.; Kim, M.S.; Xie, X.; Jeter, R.M.; Zak, J.C.; Dowd, S.E.; Paré, P.W. Choline and osmotic-stress tolerance induced in Arabidopsis by the soil microbe Bacillus subtilis (GB03). Mol. Plant Microbe Interact. 2010, 23, 1097–1104. [Google Scholar] [CrossRef] [Green Version]
- Mahoney, A.K.; Yin, C.; Hulbert, S.H. Community structure, species variation, and potential functions of rhizosphere-associated bacteria of different winter wheat (Triticum aestivum) cultivars. Front. Plant Sci. 2017, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Phsiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Bashir, S.; Khan, Y. Plant growth promoting bacteriaas an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef]
- Johnson, H.E.; Broadhurst, D.; Goodacre, R.; Smith, A.R. Metabolic fingerprinting of salt-stressed tomatoes. Phytochemistry 2003, 62, 919–928. [Google Scholar] [CrossRef]
- Hichem, H.; El Naceur, A.; Mounir, D. Effects of salt stress on photosynthesis, PSII photochemistry and thermal energy dissipation in leaves of two corn (Zea mays L.) varieties. Photosynthetica 2009, 47, 517–526. [Google Scholar] [CrossRef]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic stress responses and microbe-mediated mitigation in plants: The omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Prasad, M.; Srinivasan, R.; Chaudhary, M.; Jat, L.K. Plant growth promoting rhizobacteria (PGPR) for sustainable agriculture: Perspectives and Challenges. In PGPR Amelioration in Sustainable Agriculture; Mukesh, C., Lokesh Kumar, J., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 129–157. [Google Scholar]
- Singh, S.B.; Gowtham, H.G.; Niranjana, S.R. ACC deaminase producing PGPR invoke changes in antioxidant systems to minimize the adverse effects of salt in sunflower. RJLBPCS 2019, 5, 1–15. [Google Scholar]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef] [Green Version]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 1980. [Google Scholar]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Tiwari, S.; Singh, P.; Tiwari, R.; Meena, K.K.; Yandigeri, M.; Singh, D.P.; Arora, D.K. Salt-tolerant rhizobacteria-mediated induced tolerance in wheat (Triticum aestivum) and chemical diversity in rhizosphere enhance plant growth. Biol. Fertil. Soils. 2011, 47, 907–916. [Google Scholar] [CrossRef]
- Xu, Z.H.; Saffigna, P.G.; Farquhar, G.D.; Simpson, J.A.; Haines, R.J.; Walker, S.; Osborne, D.O.; Guinto, D. Carbon isotope discrimination and oxygen isotope composition in clones of the F1 hybrid between slash pine and Caribbean pine in relation to tree growth, water-use efficiency and foliar nutrient concentration. Tree Physiol. 2000, 20, 1209–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodd, I.C.; Pérez-Alfocea, F. Microbial amelioration of crop salinity stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, P.J.; Broadley, M.R. Chloride in soils and its uptake and movement within the plant: A review. Ann. Bot. 2001, 88, 967–988. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.M.; Naqvi, S.S.M.; Ansari, R.A. Impact of soil pH on nutrient uptake by crop plants. In Handbook of Plant and Crop Stress; CRC Press: Boca Raton, FL, USA, 1999; Volume 2, pp. 51–60. [Google Scholar]
- Al-Maliki, S.; AL-Masoudi, M. Interactions between Mycorrhizal fungi, tea wastes, and algal biomass affecting the microbial community, soil structure, and alleviating of salinity stress in corn yield (Zea mays L.). Plants 2018, 7, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, S.; Nee Sabharwal, V.P.; Kushwaha, H.R.; Sopory, S.K.; Singla-Pareek, S.L.; Pareek, A. Transcriptome map for seedling stage specific salinity stress response indicates a specific set of genes as candidate for saline tolerance in Oryza sativa L. Funct. Integr. Genomics. 2009, 9, 109–123. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sc. 2014, 19, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Shahbaz, M.; Ashraf, M. Improving salinity tolerance in cereals. Crit. Rev. Plant Sci. 2013, 32, 237–249. [Google Scholar] [CrossRef]
- Zhang, M.; Cao, Y.; Wang, Z.; Wang, Z.Q.; Shi, J.; Liang, X.; Song, W.; Chen, Q.; Lai, J.; Jiang, C. A retrotransposon in an HKT1 family sodium transporter causes variation of leaf Na+ exclusion and salt tolerance in maize. New Phytol. 2017, 217, 1161–1176. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Liang, X.; Wang, L.; Cao, Y.; Song, W.; Shi, J.; Lai, J.; Jiang, C. A HAK family Na+ transporter confers natural variation of salt tolerance in maize. Nat.Plants 2019, 5, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene expression profiling of plants under salt stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Bouis, H.E.; Welch, R.M. Biofortification—A sustainable agricultural strategy for reducing micronutrient malnutrition in the global South. Crop Sci. 2010, 50, S20–S32. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Q.; Sun, Y.X.; Ye, Y.L.; RezaulKarim, M.; Xue, Y.F.; Yan, P.; Meng, Q.F.; Cui, Z.L.; Cakmak, I.; Zhang, F.S.; et al. Zinc biofortification of wheat through fertilizer applications in different locations of china. Field Crop Res. 2012, 125, 1–7. [Google Scholar] [CrossRef]
- Cakmak, I.; Pfeiffer, W.H.; McClafferty, B. Biofortification of durum wheat with zinc and iron. Cereal Chem. 2010, 87, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; McLaughlin, M.J.; White, P. Zinc for better crop production and human health. Plant Soil 2017, 411, 1–4. [Google Scholar] [CrossRef]
- Francisco, G.C.; Poblaciones, M.J.; Almeida, A.S.; Cakmak, I. Zinc (Zn) concentration of bread wheat grown under Mediterranean conditions as affected by genotype and soil/foliar Zn application. Plant Soil 2016, 401, 331–346. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Gene Symbol | Forward Primer | Reverse Primer |
|---|---|---|---|
| (A) Zn transporter (TaZIP) genes | |||
| 1. | TaZIP1 | GTCCCCCTACTTCTACCGCT | TGGTTGACCCTCTGCCTGTT |
| 2. | TaZIP3 | GGGAAATGGAGAACYCCTGGATG | GGCATAGAGATCTTGAAAGCAATTGC |
| 3. | TaZIP5 | AGGTTTCGCCTCAAGTCTGTCTTG | GGCTATTCTCGTCGTAAGCAGAG |
| 4. | TaZIP6 | GTCATCATCTCTGAAACTGAAGAAGG | CCCTCTATACATTTCACTATGRCC |
| 5. | TaZIP7 | ACAGGCAGTATGTTSGGACGTAG | CAGCAAGTGATGGCCTATGTCG |
| 6. | TaZIP10 | GTGGATCTCATTGCTGCTGA | AGCCCAAATAGCCAGTGATG |
| 7. | TaZIP13 | CGCAAGCSTACAACATGAAACAGT | CTTYAGACACGCTACTGGGTTGG |
| 8. | TaZIP13 | CGCGAGCCTACAACTTGAAACAG | CTTYAGACACGCTACTGGGTTGG |
| 9. | TaZIP15 | CTCTCTGCGCTGGTTGCTTT | TGGGAGGACTCCGGCAACAG |
| (B) Salinity stress related genes | |||
| 10. | TaHKT-1 | CAAAGGTGAAGGAGCTGAGG | GAGCTGAGCCCATCAAAGAC |
| 11. | TaNHX-1 | GAATGCCACTCAGATCCAGC | GCTGCTGGGTGGCTTAGTGC |
| (C) Housekeeping genes | |||
| 12. | TaActin3 | GACGCACAACAGGTATCGTGTTG | CAGCGAGGTCAAGACGAAGGATG |
| 13. | TaSuccDH | TTTGCTCTCCGTGGTGCCTTTGG | GAAGATGTGTAGCTCCTTGCTTGC |
| Treatments | Germination (%) | VigourIndex I | VigourIndex II |
|---|---|---|---|
| T. harzianum UBSTH-501 | 87.23 ± 2.25 a | 3413.22 ± 10.21 a | 1.95 ± 0.45 a |
| B. amyloliquefaciens B-16 | 85.26 ± 2.50 b | 3324.16 ± 9.26 b | 1.87 ±0.25 a |
| Control (untreated) | 80.92 ± 3.52 c | 3112.25± 8.20 c | 1.75 ± 0.25 b |
| Treatments | Fungi | Bacteria | Actinomycetes | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 30 DAS † | 60 DAS | 90 DAS | 120 DAS | 30 DAS | 60 DAS | 90 DAS | 120 DAS | 30 DAS | 60 DAS | 90 DAS | 120 DAS | |
| Control (untreated) | 2.02 ± 0.25 b | 3.01 ± 0.15 a | 2.92 ± 0.50 b | 2.62 ± 0.57 b | 2.15 ± 0.50 b | 3.46 ± 0.16 b | 3.92 ± 0.15 b | 3.67 ± 0.25 b | 1.61 ± 0.10 a | 1.92 ± 0.15 b | 2.01 ± 0.33 b | 1.72 ± 0.11 b |
| Bioagents treated | 2.85 ± 0.33 a | 3.01 ± 0.36 a | 3.96 ± 0.33 a | 3.30 ± 0.66 a | 3.56 ± 0.20 a | 6.96 ± 0.66 a | 7.12 ± 1.15 a | 4.92 ± 0.50 a | 1.88 ± 0.33 a | 2.67 ± 0.20 a | 2.46 ± 0.45 a | 2.42 ± 0.25 a |
| Treatments | Plant Height (cm) | Number of Tillers Plant−1 | Plant Biomass on Dry wt. Basis (g) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 30 DAS † | 60 DAS | 90 DAS | 120 DAS | 30 DAS | 60 DAS | 90 DAS | 120 DAS | 30 DAS | 60 DAS | 90 DAS | 120 DAS | |
| Control (untreated) | 17.50 ± 1.02 b | 38.60 ± 2.25 b | 70.30 ± 2.50 b | 75.05 ± 1.57 b | 2.05 ± 0.55 b | 4.30 ± 0.36 b | 4.90 ± 0.25 b | 4.96 ± 0.45 b | 1.89 ± 0.22 b | 4.80 ± 0.50 b | 5.21 ± 0.33 b | 6.88 ± 0.21 b |
| Bioagents treated | 24.70 ± 2.01 a | 61.90 ± 2.36 a | 95.40 ± 3.50 a | 100.30 ± 2.65 a | 3.25 ± 0.22 a | 8.40 ± 1.02 a | 8.90 ± 0.66 a | 8.90 ± 0.40 a | 2.35 ± 0.11 a | 6.54 ± 0.25 a | 8.20 ± 0.25 a | 10.50 ± 0.75 a |
| Treatments | Na+ Uptake | K+ Uptake | Ca2+ Uptake | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 30 DAS † | 60 DAS | 90 DAS | 120 DAS | 30 DAS | 60 DAS | 90 DAS | 120 DAS | 30 DAS | 60 DAS | 90 DAS | 120 DAS | |
| Control (untreated) | 0.44 ± 0.01 a | 3.06 ± 0.25 a | 5.93 ± 0.55 a | 8.68 ± 1.02 a | 0.58 ± 0.01 b | 2.36 ± 0.20 b | 3.90 ± 0.66 b | 6.50 ± 0.80 b | 0.45 ± 0.01 a | 2.20 ± 0.19 b | 3.77 ± 0.45 b | 7.00 ± 0.95 b |
| Bioagents treated | 0.54 ± 0.05 a | 2.82 ± 0.33 b | 5.71 ± 0.82 b | 7.10 ± 0.66 b | 1.06 ± 0.15 a | 5.54 ± 0.55 a | 9.36 ± 1.33 a | 13.09 ± 1.66 a | 0.75 ± 0.01 a | 4.25 ± 0.25 a | 7.86 ± 0.75 a | 11.44 ± 4.25 a |
| Parameters | Control (Untreated) | Bioagents Treated |
|---|---|---|
| Spike length (cm) | 9.10 ± 1.02 b | 11.20 ± 1.15 a |
| Spike weight (g) | 1.66 ± 0.20 b | 2.10 ± 0.21 a |
| Spikelet spike −1 | 17.05 ± 1.25 b | 19.50 ± 1.45 a |
| Number of seeds spike −1 | 44.80 ± 2.01 b | 50.70 ± 3.02 a |
| Seed weight (g) spike −1 | 1.50 ± 0.11 b | 1.70 ± 0.33 a |
| Test weight (g) | 37.30 ± 1.33 b | 39.20 ± 1.36 a |
| Seed yield (q ha −1) | 22.80 ± 1.03 b | 27.20 ± 1.25 a |
| Straw yield (q ha −1) | 33.20 ± 1.25 b | 41.70 ± 1.54 a |
| Biological yield (q ha −1) | 56.00 ± 2.25 b | 68.90 ± 2.66 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, U.B.; Malviya, D.; Singh, S.; Singh, P.; Ghatak, A.; Imran, M.; Rai, J.P.; Singh, R.K.; Manna, M.C.; Sharma, A.K.; et al. Salt-Tolerant Compatible Microbial Inoculants Modulate Physio-Biochemical Responses Enhance Plant Growth, Zn Biofortification and Yield of Wheat Grown in Saline-Sodic Soil. Int. J. Environ. Res. Public Health 2021, 18, 9936. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18189936
Singh UB, Malviya D, Singh S, Singh P, Ghatak A, Imran M, Rai JP, Singh RK, Manna MC, Sharma AK, et al. Salt-Tolerant Compatible Microbial Inoculants Modulate Physio-Biochemical Responses Enhance Plant Growth, Zn Biofortification and Yield of Wheat Grown in Saline-Sodic Soil. International Journal of Environmental Research and Public Health. 2021; 18(18):9936. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18189936
Chicago/Turabian StyleSingh, Udai B., Deepti Malviya, Shailendra Singh, Prakash Singh, Abhijeet Ghatak, Muhammad Imran, Jai P. Rai, Rajiv K. Singh, Madhab C. Manna, Arun K. Sharma, and et al. 2021. "Salt-Tolerant Compatible Microbial Inoculants Modulate Physio-Biochemical Responses Enhance Plant Growth, Zn Biofortification and Yield of Wheat Grown in Saline-Sodic Soil" International Journal of Environmental Research and Public Health 18, no. 18: 9936. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18189936