
Effects of Modified Clay on Phaeocystis globosa Growth and Colony Formation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Sampling and Analysis
2.3.1. Microscopy
2.3.2. pH and Nutrient Analysis
2.3.3. Polysaccharide
2.4. Statistical Analysis
3. Results
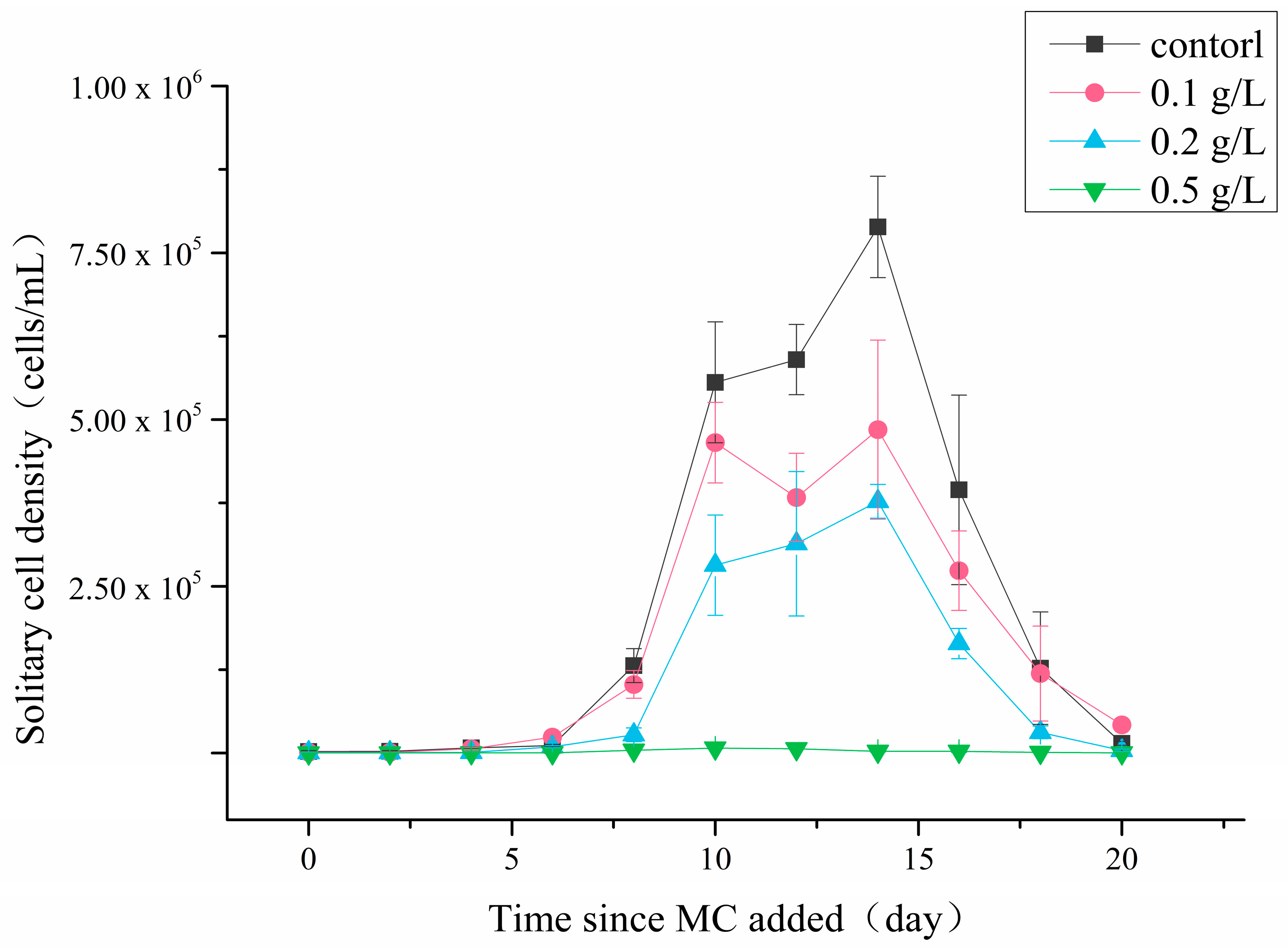
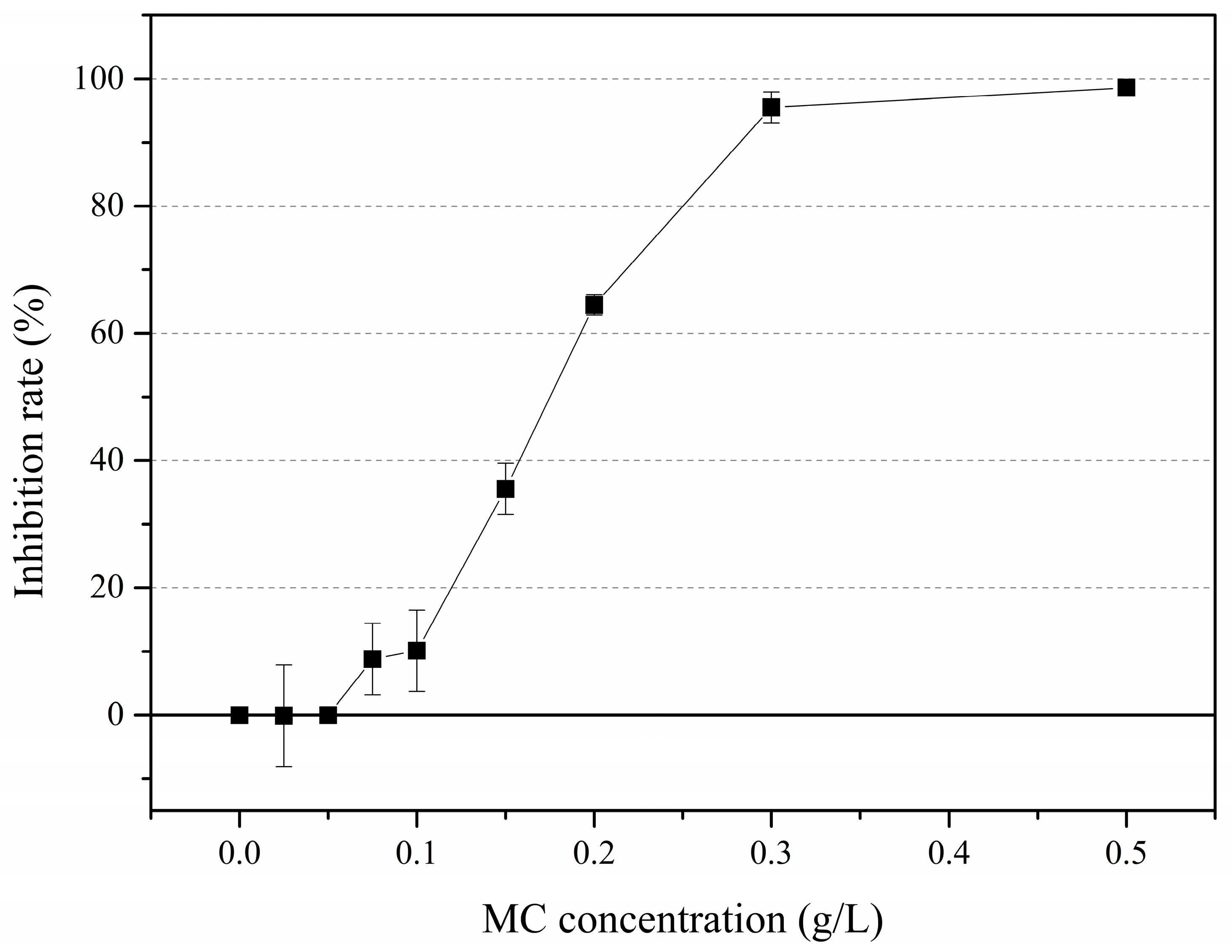
3.1. Solitary Cell Abundances
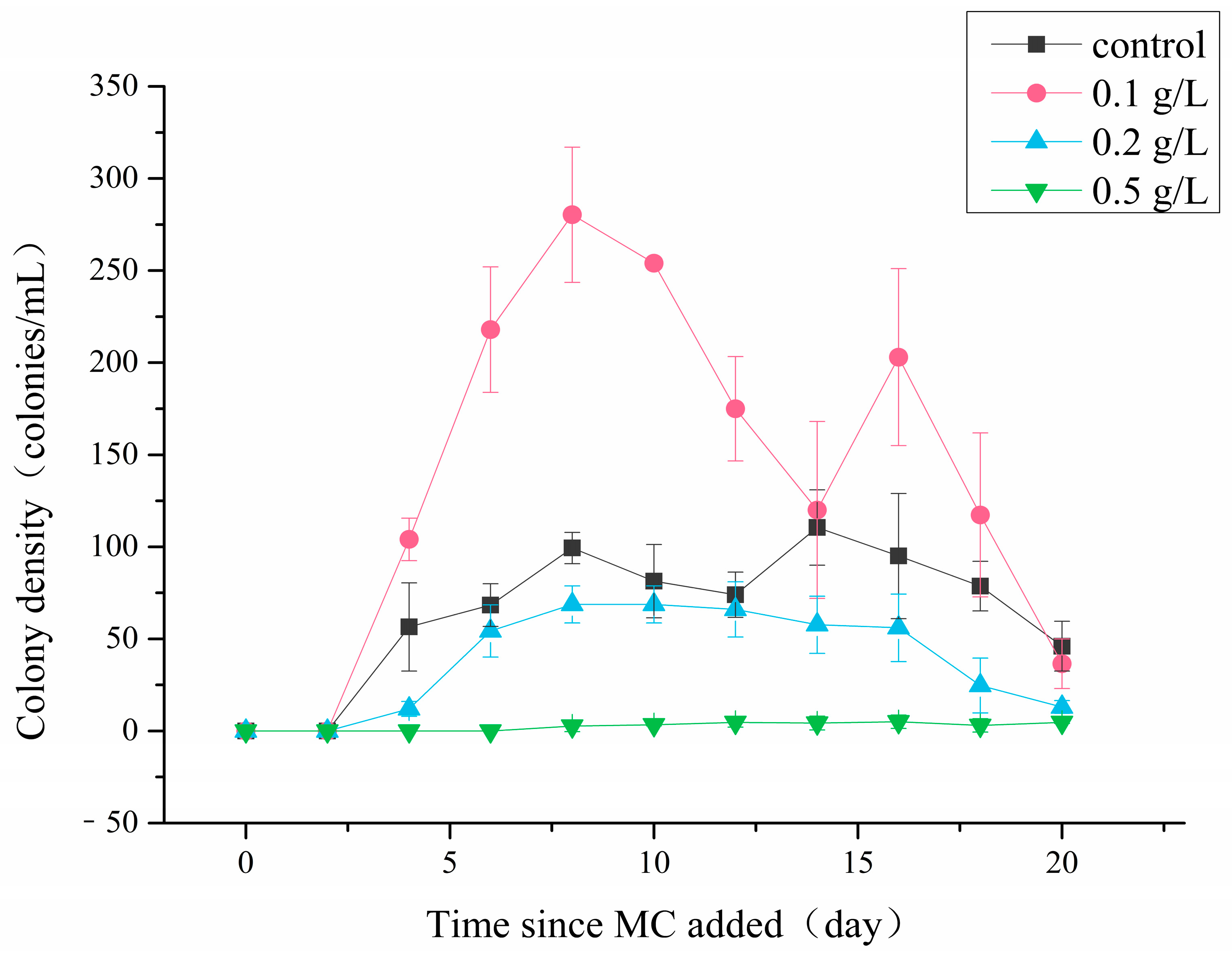
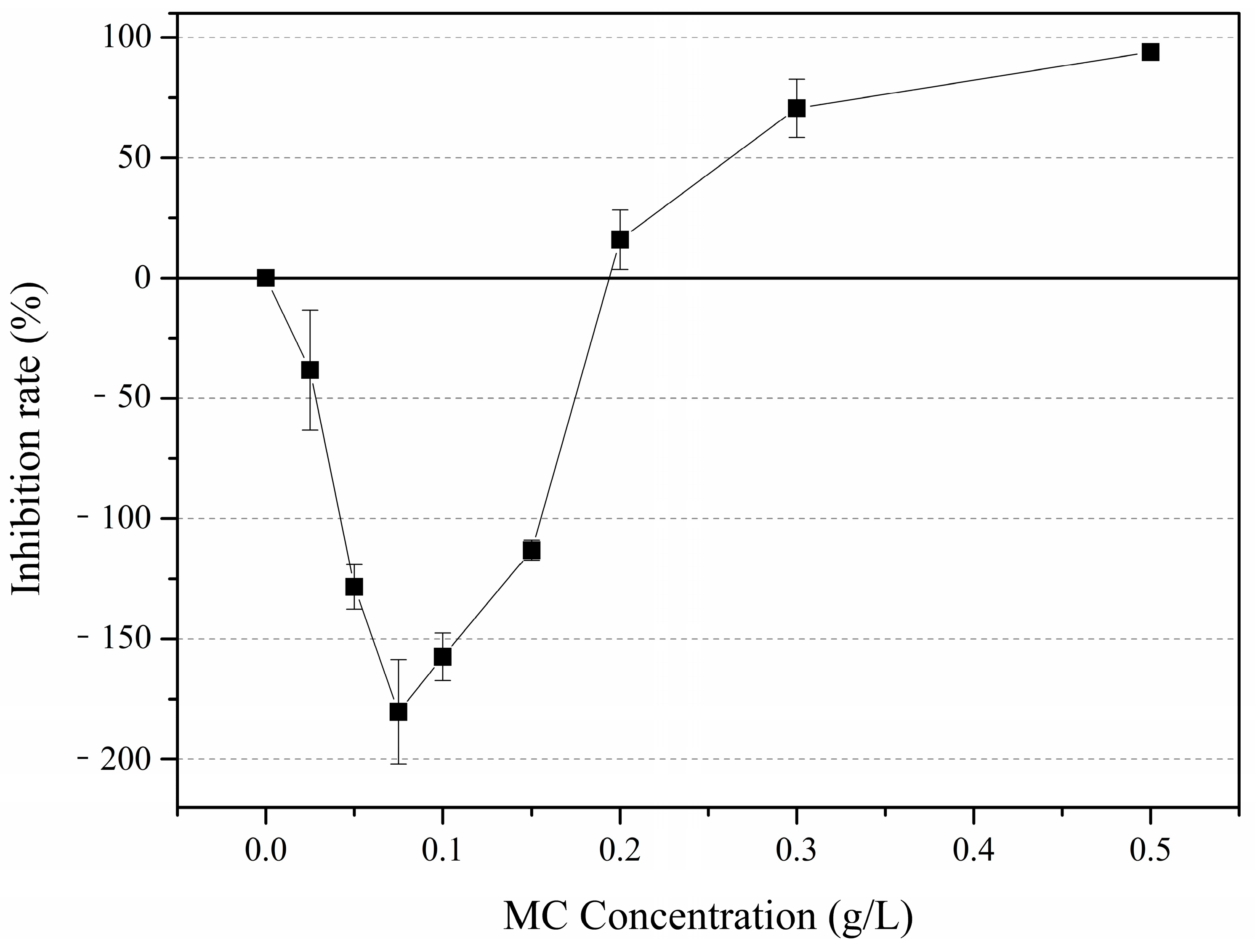
3.2. Colony Abundances
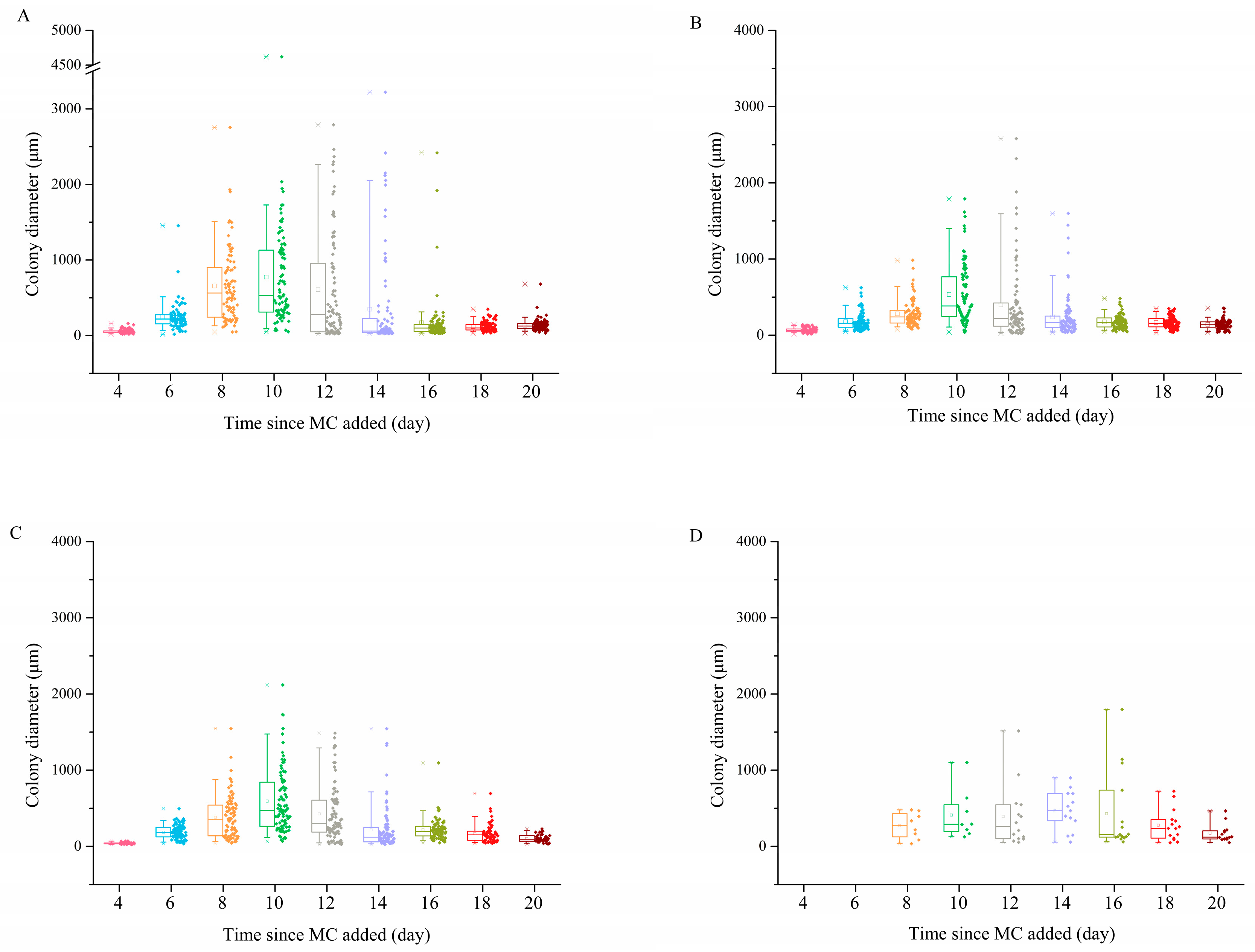
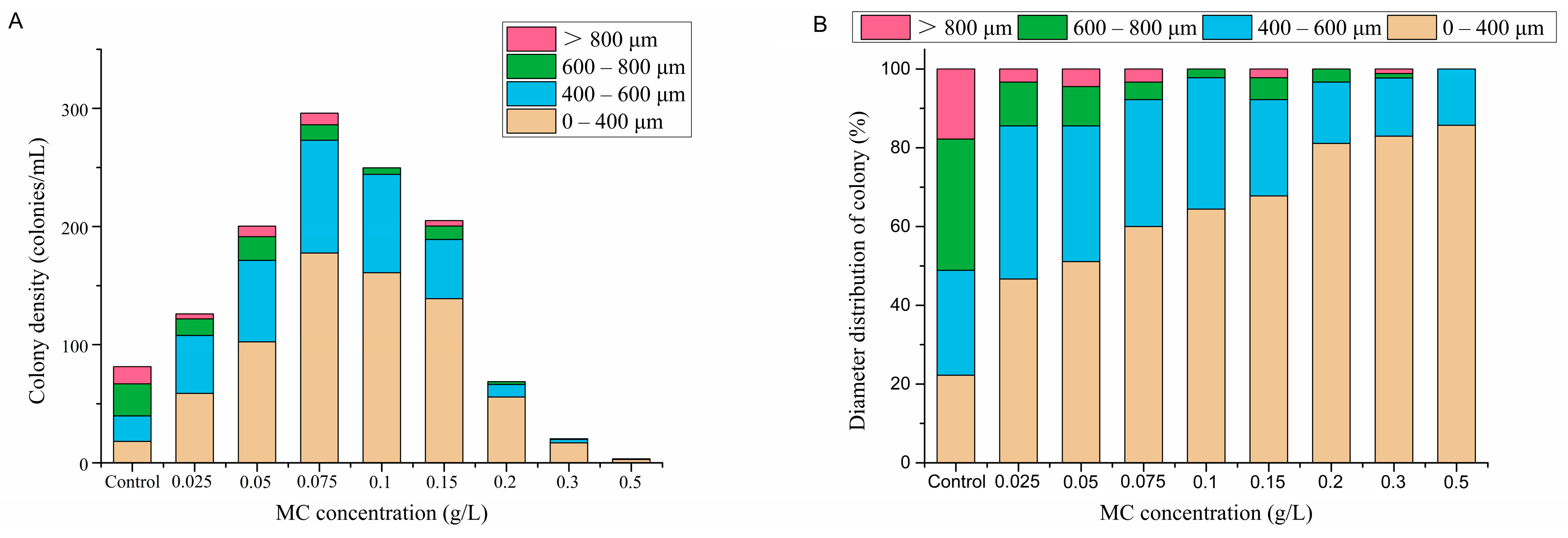
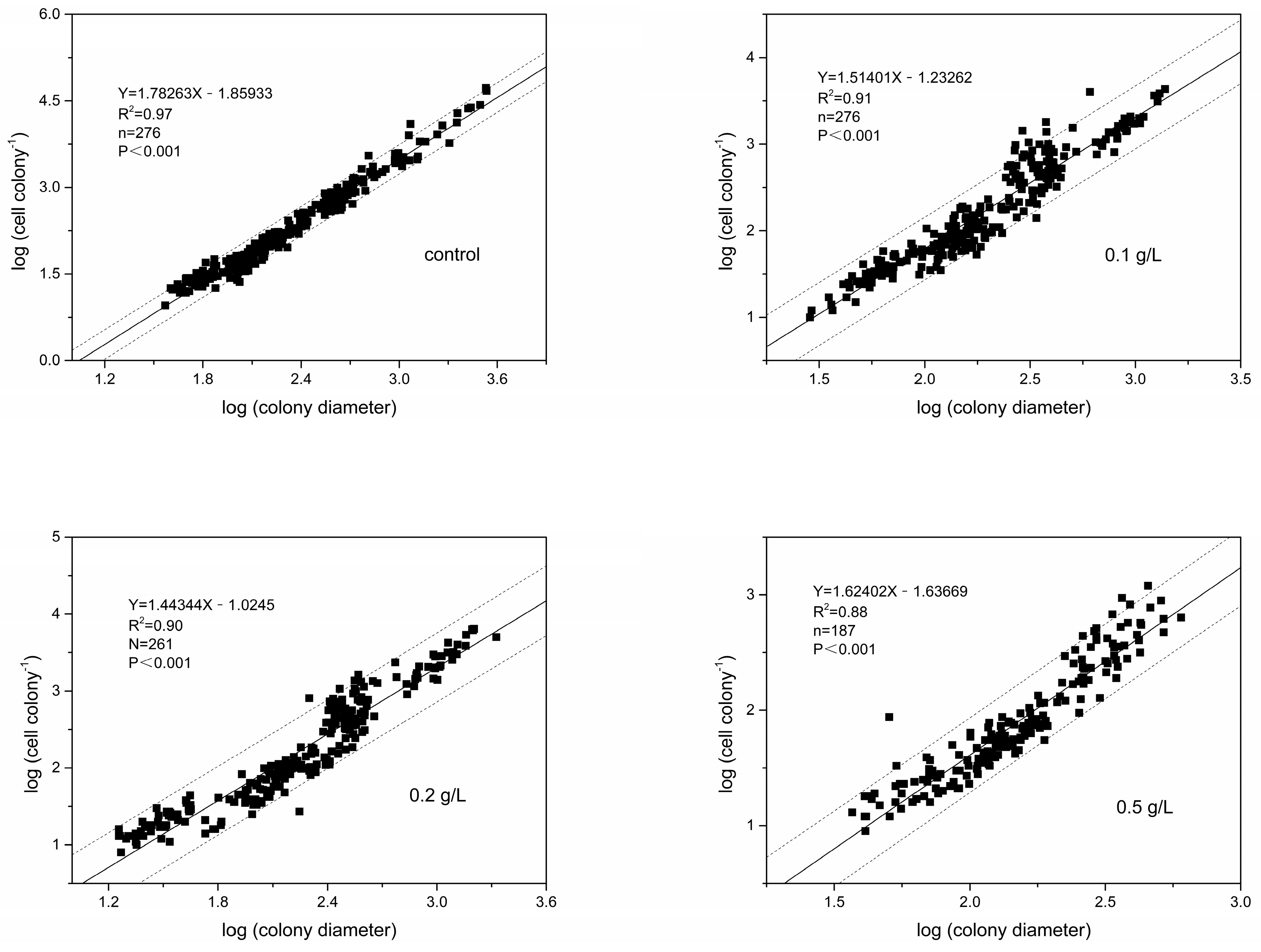
3.3. Colony Morphology
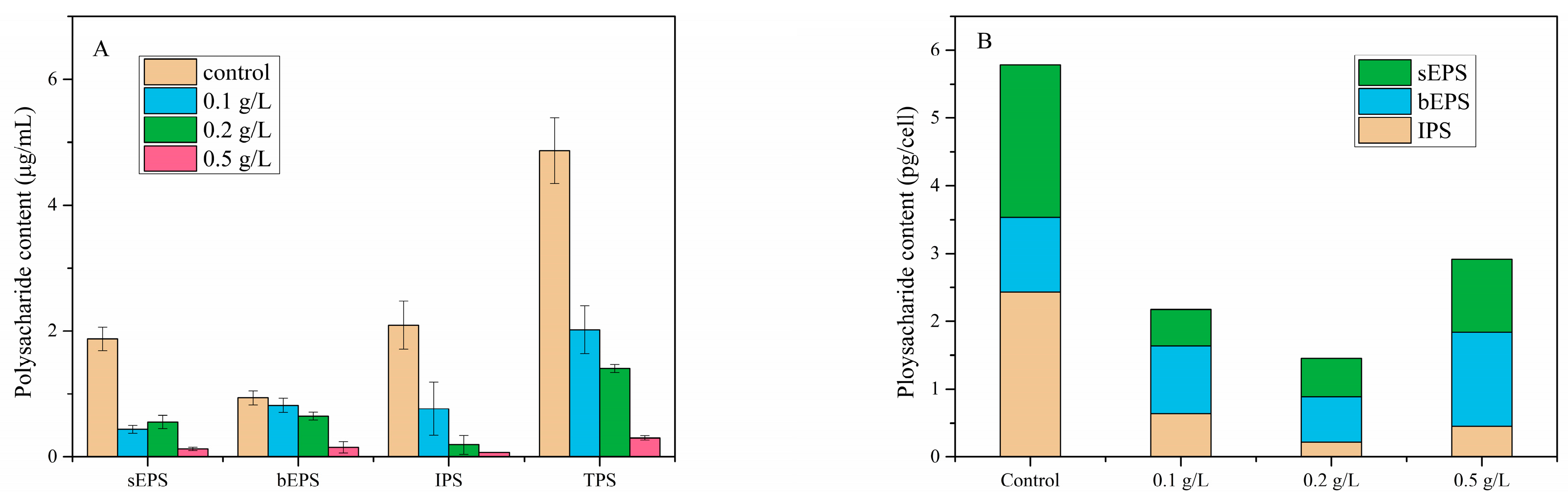
3.4. Polysaccharide Contents
3.5. Environmental Parameters
4. Discussion
4.1. Effects of MC on P. globosa Solitary Cell Abundances
4.2. Effects of MC on P. globosa Colony Abundances and Colony Morphology
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lancelot, C.; Mathot, S. Biochemical fractionation of primary production by phytoplankton in Belgian coastal waters during short- and long-term incubations with 14 C-bicarbonate. Mar. Biol. 1985, 86, 227–232. [Google Scholar] [CrossRef]
- Liss, P.S.; Malin, G.; Turner, S.M.; Holligan, P.M. Dimethyl sulphide and Phaeocystis: A review. J. Mar. Syst. 1994, 5, 41–53. [Google Scholar] [CrossRef]
- Schoemann, V.; Becquevort, S.; Stefels, J.; Rousseau, V.; Lancelot, C. Phaeocystis blooms in the global ocean and their controlling mechanisms: A review. J. Sea Res. 2005, 53, 43–66. [Google Scholar] [CrossRef]
- Buchan, A.; Lecleir, G.R.; Gulvik, C.A.; González, J.M. Master recyclers: Features and functions of bacteria associated with phytoplankton blooms. Nat. Rev. Microbiol. 2014, 12, 686–698. [Google Scholar] [CrossRef]
- Rousseau, V.; Vaulot, D.; Casotti, R.; Cariou, V.; Lenz, J.; Gunkel, J.; Baumann, M. The life cycle of Phaeocystis (Prymnesiophycaea): Evidence and hypotheses. J. Mar. Syst. 1994, 5, 23–39. [Google Scholar] [CrossRef]
- Rousseau, V.; Chrétiennot-Dinet, M.J.; Jacobsen, A.; Nejstgaard, J.C.; Verity, P.G. The life cycle of Phaeocystis: State of knowledge and presumptive role in ecology. Biogeochemistry 2007, 83, 29–47. [Google Scholar] [CrossRef]
- Rijssel, M.V.; Hamm, C.E.; Gieskes WW, C. Phaeocystis globosa (Prymnesiophyceae) colonies: Hollow structures built with small amounts of polysaccharides. Eur. J. Phycol. 1997, 32, 185–192. [Google Scholar]
- Chen, Y.Q.; Wang, N.; Zhang, P.; Zhou, H.; Qu, L.H. Molecular evidence identifies bloom-forming Phaeocystis (Prymnesiophyta) from coastal waters of southeast China as Phaeocystis globosa. Biochem. Syst. Ecol. 2002, 30, 15–22. [Google Scholar] [CrossRef]
- Alderkamp, A.C.; Buma, A.G.J.; Rijssel, M.V. The carbohydrates of Phaeocystis and their degradation in the microbial food web. Biogeochemistry 2007, 83, 99–118. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, H.H.; Tang, K.W. Effects of protozoan grazing on colony formation in Phaeocystis globose (Prymnesiophyceae) and the potential costs and benefits. Aquat. Microb. Ecol. 2002, 27, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Tang, K.W. Grazing and colony size development in Phaeocystis globosa (Prymnesiophyceae): The role of a chemical signal. J. Plankton Res. 2003, 25, 831–842. [Google Scholar] [CrossRef]
- Brussaard, C.P.D.; Kuipers, B.; Veldhuis, M.J.W. A mesocosm study of Phaeocystis globosa population dynamics: I. Regulatory role of viruses in bloom control. Harmful Algae 2005, 4, 875–893. [Google Scholar] [CrossRef]
- Brussaard, C.P.D.; Bratbak, G.; Baudoux, A.C.; Ruardij, P. Phaeocystis and its interaction with viruses. Biogeochemistry 2007, 83, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Brisbin, M.M.; Mitarai, S. Differential gene expression supports a resource-intensive, defensive role for colony production in the bloom-forming haptophyte, Phaeocystis globosa. J. Eukaryot. Microbiol. 2019, 66, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lancelot, C. Phaeocystis blooms and nutrient enrichment in the continental coastal zones of the North Sea. AMBIO 1987, 16, 38–46. [Google Scholar]
- Alderkamp, A.C.; Nejstgaard, J.C.; Verity, P.G.; Zirbel, M.J.; Sazhin, A.F.; Rijssel, M.V. Dynamics in carbohydrate composition of Phaeocystis pouchetii colonies during spring blooms in mesocosms. J. Sea Res. 2006, 55, 169–181. [Google Scholar] [CrossRef]
- Baumann, M.E.M.; Lancelot, C.; Brandini, F.P.; Sakshaugd, E.; Johne, D.M. The taxonomic identity of the cosmopolitan prymnesiophyte Phaeocystis: A morphological and ecophysiological approach. J. Mar. Syst. 1994, 5, 5–22. [Google Scholar] [CrossRef]
- Madhupratap, M.; Sawant, S.; Gauns, M. A first report on a bloom of the marine prymnesiophycean, Phaeocystis globosa from the Arabian Sea. Oceanol. Acta 2000, 23, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Spilmont, N.; Denis, L.; Artigas, L.F.; Caloin, F.; Courcot, L.; Creach, A.; Desroy, N.; Gevaert, F.; Hacquebart, P.; Hubas, C.; et al. Impact of the Phaeocystis globosa spring bloom on the intertidal benthic compartment in the eastern English Channel: A synthesis. Mar. Pollut. Bull. 2009, 58, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.H.; Yu, Z.M.; Qiu, L.X. Field experiment and emergent application of modified clays for Phaeocystis globosa blooms mitigation. Oceanologia et Limnologia Sinica. 2017, 48, 753–759, (In Chinese with English Abstract). [Google Scholar]
- Yu, Z.M.; Song, X.X.; Cao, X.H.; Liu, Y. Mitigation of harmful algal blooms using modified clays: Theory, mechanisms, and applications. Harmful Algae 2017, 69, 48–64. [Google Scholar] [CrossRef]
- Anderson, D.M. Turning back the harmful red tide. Nature 1997, 388, 513–514. [Google Scholar] [CrossRef]
- Sengco, M.R.; Anderson, D.M. Controlling Harmful Algal Blooms Through Clay Flocculation1. J. Eukaryot. Microbiol. 2004, 51, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, S.E.; Sengco, M.R.; Anderson, D.M. Using clay to control harmful algal blooms: Deposition and resuspension of clay/algal flocs. Harmful Algae 2005, 4, 123–138. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Zhang, H.; Liu, X.A.; Cai, D.Q.; Feng, H.Y.; Miao, C.G.; Wang, X.Q.; Wu, Z.Y.; Yu, Z.L. Flocculation of harmful algal blooms by modified attapulgite and its safety evaluation. Water Res. 2011, 45, 2855–2862. [Google Scholar] [CrossRef]
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in Understanding Harmful Algal Blooms: Paradigm Shifts and New Technologies for Research, Monitoring, and Management. Annu. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.M.; Zou, J.Z.; Ma, X.N. A new method to improve the capability of clays for removing red tide organisms. Oceanologia Et Limnologia Sinica. 1994, 25, 226–232, (In Chinese with English Abstract). [Google Scholar]
- Liu, Y.; Cao, X.H.; Yu, Z.M.; Song, X.X.; Qiu, L.X. Controlling harmful algae blooms using aluminum-modified clay. Mar. Pollut. Bull. 2016, 103, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Yu, Z.M.; Song, X.X.; Cao, X.H. Effects of modified clay on the physiological and photosynthetic activities of Amphidinium carterae Hulburt. Harmful Algae 2017, 70, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Yu, Z.M.; Song, X.X.; Cao, X.H. Physiological and photosynthetic responses of Karenia mikimotoi to the modified clay mitigation method. Mar. Pollut. Bull. 2018, 133, 491–499. [Google Scholar] [CrossRef]
- Zhu, J.N.; Yu, Z.M.; He, L.Y.; Cao, X.H.; Liu, S.Y.; Song, X.X. Molecular Mechanism of Modified Clay Controlling the Brown Tide Organism Aureococcus anophagefferens Revealed by Transcriptome Analysis. Environ. Sci. Technol. 2018, 52, 7006–7014. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, Z.; Song, X.; Yuan, Y.Q.; Cao, X.H. Effects of modified clay used for the control of harmful algal blooms on Alexandrium pacificum cysts. Harmful Algae 2018, 72, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Song, X.; Fan, X.; Yu, Z.M. Flocculation of Ulva microscopic propagules using modified clay: A mesocosm experiment. J. Oceanol. Limnol. 2020, 38, 1283–1291. [Google Scholar] [CrossRef]
- Xu, N.; Huang, B.Z.; Hu, Z.X.; Tang, Y.Z.; Duan, S.S.; Zhang, C.W. Effects of temperature, salinity, and irradiance on the growth of harmful algal bloom species Phaeocystis globosa Scherffel (Prymnesiophyceae) isolated from the South China Sea. Chin. J. Oceanol. Limnol. 2017, 035, 557–565. [Google Scholar] [CrossRef]
- Sun, Y.F.; Wang, Y.Y.; Lei, J.; Qian, C.C.; Zhu, X.X.; Akbar, S.; Huang, Y.; Yang, Z. Temperature-dependent elimination efficiency on Phaeocystis globosa by different initial population sizes of rotifer Brachionus plicatilis. Chemosphere 2018, 203, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Yu, Z.M.; Zou, J.Z.; Ma, X.N. Application of clays to removal of red tide organisms II. Coagulation of different species of red tide organisms with montmorillonite and effect of clay pretreatment. Chin. J. Oceanol. Limnol. 1994, 12, 316–324. [Google Scholar]
- Lu, G.Y.; Song, X.X.; Yu, Z.M.; Cao, X.H. Application of PAC-modified kaolin to mitigate Prorocentrum donghaiense: Effects on cell removal and phosphorus cycling in a laboratory setting. J. Appl. Phycol. 2017, 29, 1–12. [Google Scholar] [CrossRef]
- Lu, G.Y.; Song, X.X.; Yu, Z.M.; Cao, X.H.; Yuan, Y.Q. Effects of modified clay flocculation on major nutrients and diatom aggregation during Skeletonema costatum blooms in the laboratory. Chin. J. Oceanol. Limnol. 2015, 33, 1007–1019. [Google Scholar] [CrossRef]
- Wang, X.D.; Wang, Y.; Smith, O.W. The role of nitrogen on the growth and colony development of Phaeocystis globosa (Prymnesiophyceae). Eur. J. Phycol. 2011, 46, 305–314. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, F.X.; Shi, X.L.; Zhang, M.; Xing, P.; Cao, H.S. Changes in the morphology and polysaccharide content of Microcystis aeruginosa (Cyanobacteria) during flagellate grazing. J. Phycol. 2008, 44, 716–720. [Google Scholar] [CrossRef]
- Li, M.; Zhu, W.; Gao, L.; Lu, L. Changes in extracellular polysaccharide content and morphology of Microcystis aeruginosa at different specific growth rates. J. Appl. Phycol. 2013, 25, 1023–1030. [Google Scholar] [CrossRef]
- Qiu, L.X.; Yu, Z.M.; Cao, X.H.; Ji, H.N.; Song, X.X. Study on the Mechanism of a New Type of Modified Clay Controlling Phaeocystis globosa Growth. J. Oceanol. Limnol. 2020, 38, 1270–1282. [Google Scholar] [CrossRef]
- Zhu, J.N.; Yu, Z.M.; He, L.Y.; Cao, X.H.; Ji, H.N.; Song, X.X. Physiological response dynamics of the brown tide organism Aureococcus anophagefferens treated with modified clay. Harmful Algae. 2019, 86, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, M.J.; Smith, S.V. C:N:P ratios of benthic marine plants. Limnol. Oceanogr. 1983, 28, 568–574. [Google Scholar] [CrossRef]
- Gauthier, D.A.; Turpin, D.H. Interactions between inorganic phosphate (Pi) assimilation, photosynthesis and respiration in the Pi-limited green alga Selenastrum minutum. Plant Cell Environ. 1997, 20, 12–24. [Google Scholar] [CrossRef]
- Sebastian, L.; Dion, M.F.F.; Susan, I.B.; Stephanie, C.G.; Andrew, P.N. Influence of phosphorus limitation on toxicity and photosynthesis of alexandrium minutum (dinophyceae) monitored by in-line detection of variable chlorophyll fluorescence1. J. Phycol. 2003, 39, 320–331. [Google Scholar]
- Qi, Y.Z.; Chen, J.F.; Wang, Z.H.; Xu, N.; Wang, Y.; Shen, P.P.; Lu, S.H.; Hodgkiss, I.J. Some observations on harmful algal bloom (HAB) events along the coast of Guangdong, southern China in 1998. Hydrobiologia 2004, 512, 209–214. [Google Scholar] [CrossRef]
- Van Rijssel, M.; Janse, I.; Noordkamp, D.J.B.; Gieskes, W.W.C. An inventory of factors that affect polysaccharide production by Phaeocystis globosa. J. Sea Res. 2000, 43, 297–306. [Google Scholar] [CrossRef]
- Cariou, V.; Casotti, R.; Birrien, J.L.; Vaulot, D. The initiation of Phaeocystis colonies. J. Plankton Res. 1994, 16, 457–470. [Google Scholar] [CrossRef]
- Peperzak, L.; Gaebler-Schwarz, S. Current knowledge of the life cycles of phaeocystis globosa and phaeocystis antarctica (prymnesiophyceae). J. Phycol. 2012, 48, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Verity, P.G.; Whipple, S.J.; Nejstgaard, J.C.; Alderkamp, A.C. Colony size, cell number, carbon and nitrogen contents of Phaeocystis pouchetii from western Norway. J. Plankton Res. 2007, 29, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, V.; Lantoine, F.; Rodriguez, F.; Legall, F.; Chrétiennot-Dinet, M.J.; Lancelot, C. Characterization of Phaeocystis globosa (Prymnesiophyceae), the blooming species in the Southern North Sea. J. Sea Res. 2013, 76, 105–113. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zhang, S.; Peng, Y.; Li, Y.; Chen, Z.R.; Xu, H.; Yu, Z.M.; Zheng, W.; Zheng, T.L. Effects of marine actinomycete on the removal of a toxicity alga Phaeocystis globose in eutrophication waters. Front. Microbiol. 2015, 6, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, C.W.; Guo, X.Y.; Li, Y.; Zhang, H.J.; Lei, X.Q.; Cai, G.J.; Guo, J.J.; Yu, Z.M.; Zheng, T.L. Photoinhibition of Phaeocystis globosa resulting from oxidative stress induced by a marine algicidal bacterium Bacillus sp. LP-10. Sci. Rep. 2015, 5, 17002. [Google Scholar] [CrossRef]
- Veldhuis, M.J.W.; Admiraal, W. Influence of phosphate depletion on the growth and colony formation of Phaeocystis pouchetii. Mar. Biol. 1987, 95, 47–54. [Google Scholar] [CrossRef]
- Marina, P.; Thomas, K. Phytoplankton defence mechanisms: Traits and trade-offs. Biol. Rev. 2018, 93, 1269–1303. [Google Scholar]
- Smith, W.O.; Liu, X.; Tang, K.W.; Delizo, L.M.; Doan, N.H.; Nguyen, N.L.; Wang, X.D. Giantism and its role in the harmful algal bloom species Phaeocystis globosa. Deep Sea Res. Top. Stud. Oceanogr. 2014, 101, 95–106. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Solitary Cells | Colony Cells | Total Cells | ||
|---|---|---|---|---|---|
| Density (×103 Cell/mL) | Percentage (%) | Density (×103 Cell/mL) | Percentage (%) | Density (×103 Cell/mL) | |
| Control | 555.8 ± 90.6 | 65 | 300.8 ± 35.1 | 35 | 856.7 ± 157.3 |
| 0.1 g/L | 465.56 ± 60.3 | 66 | 238.1 ± 51.3 | 34 | 703.6 ± 17.9 |
| 0.2 g/L | 281.7 ± 75.2 | 66 | 143.0 ± 38.4 | 34 | 424.6 ± 57.9 |
| 0.5 g/L | 7.6 ± 0.8 | 76 | 2.4 ± 0.8 | 24 | 10.0 ± 1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, X.; Yu, Z.; Qiu, L.; Cao, X.; Song, X. Effects of Modified Clay on Phaeocystis globosa Growth and Colony Formation. Int. J. Environ. Res. Public Health 2021, 18, 10163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph181910163
Ren X, Yu Z, Qiu L, Cao X, Song X. Effects of Modified Clay on Phaeocystis globosa Growth and Colony Formation. International Journal of Environmental Research and Public Health. 2021; 18(19):10163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph181910163
Chicago/Turabian StyleRen, Xiangzheng, Zhiming Yu, Lixia Qiu, Xihua Cao, and Xiuxian Song. 2021. "Effects of Modified Clay on Phaeocystis globosa Growth and Colony Formation" International Journal of Environmental Research and Public Health 18, no. 19: 10163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph181910163