
The Function of the Kynurenine Pathway in the Placenta: A Novel Pharmacotherapeutic Target?


Abstract
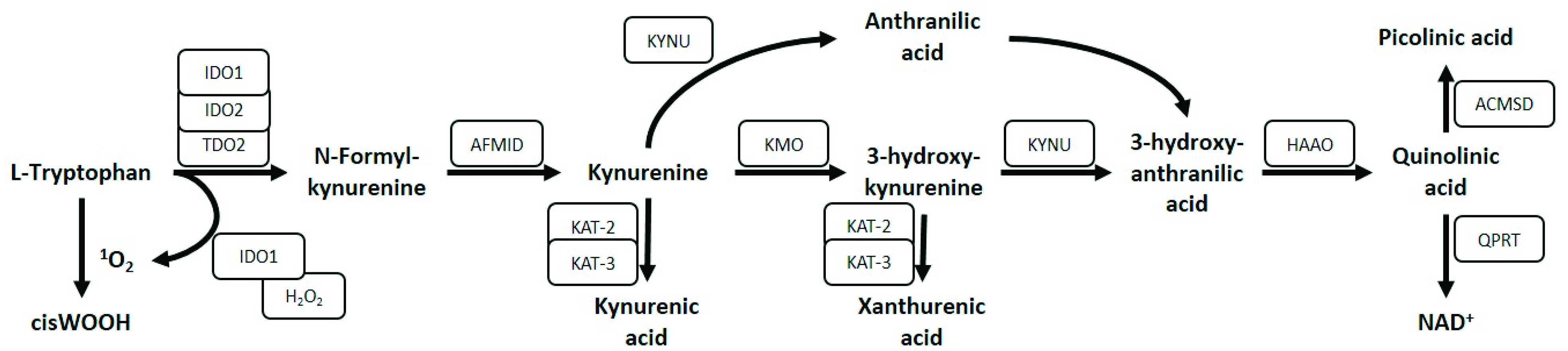
:1. Introduction
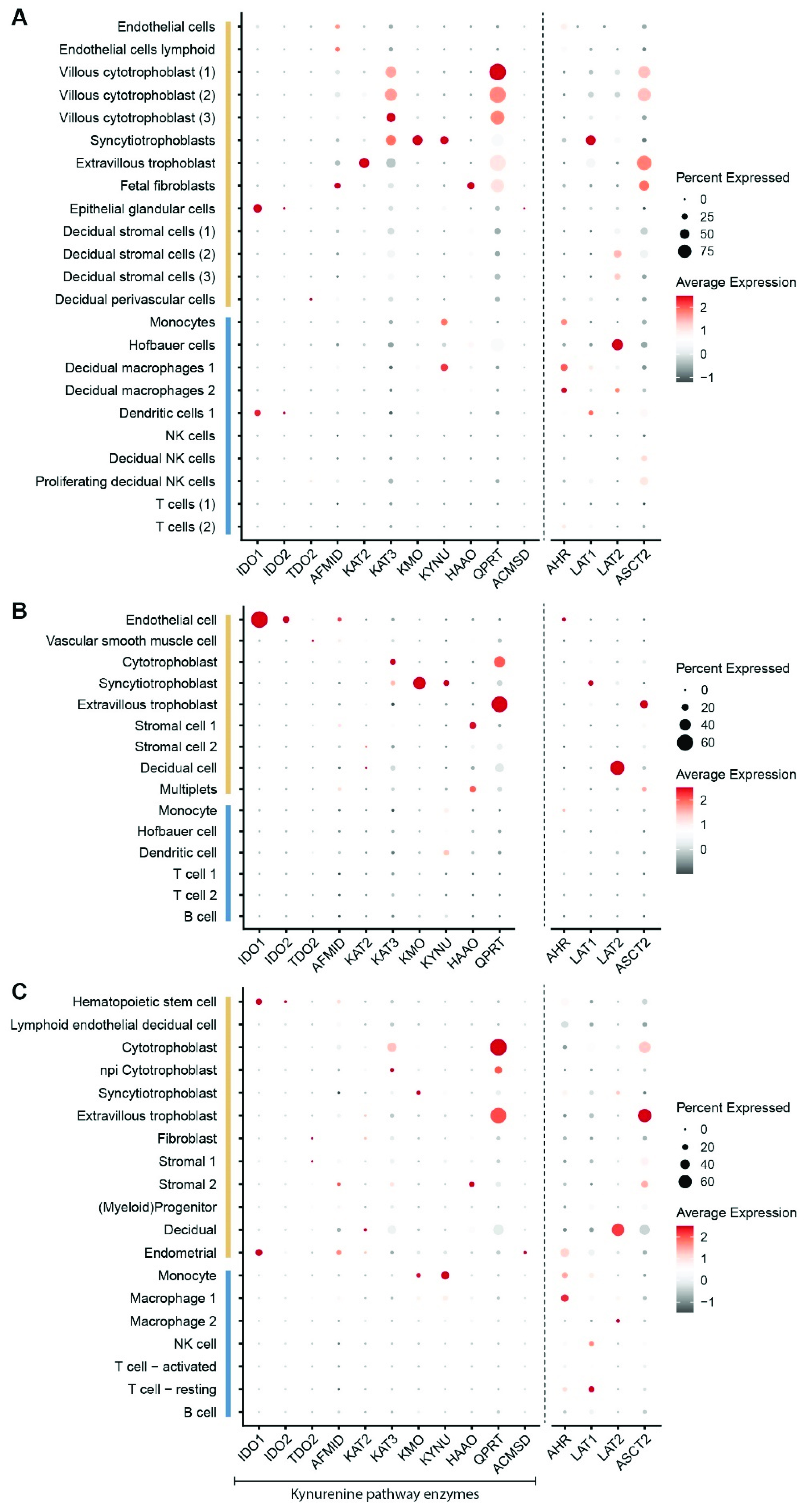
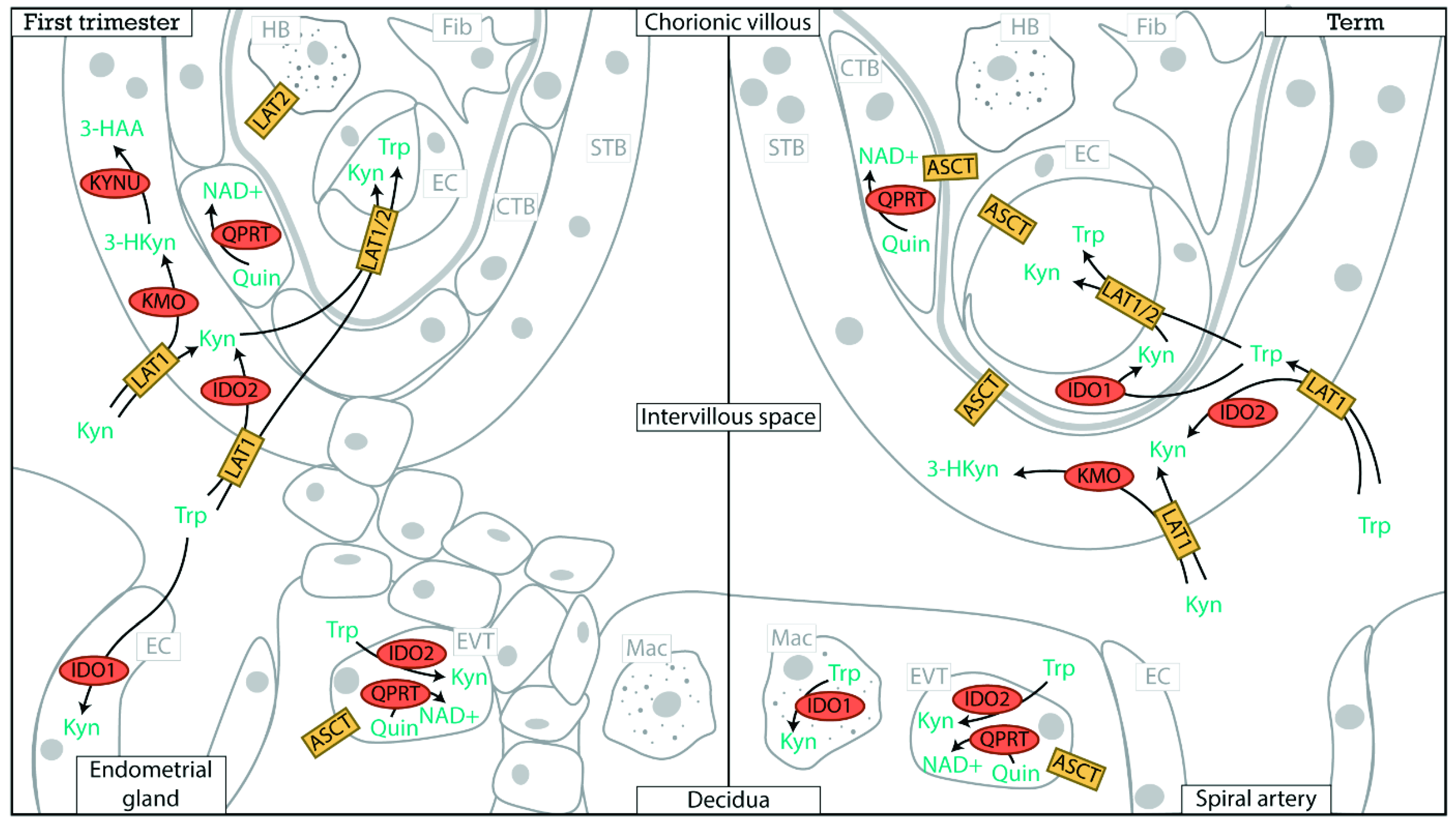
2. Placental Expression of Kynurenine Pathway Enzymes
3. Placental Tryptophan Transport
4. Kynurenine Pathway Functions
4.1. Immune Regulation
4.2. Vasodilation
4.3. Placental Vascular Development
4.4. Pro- and Anti-Oxidant Roles of the Kynurenine Pathway
5. Pathological Pregnancies
5.1. Recurrent Miscarriage
5.2. Preterm Birth
5.3. Preeclampsia
5.4. Fetal Growth Restriction
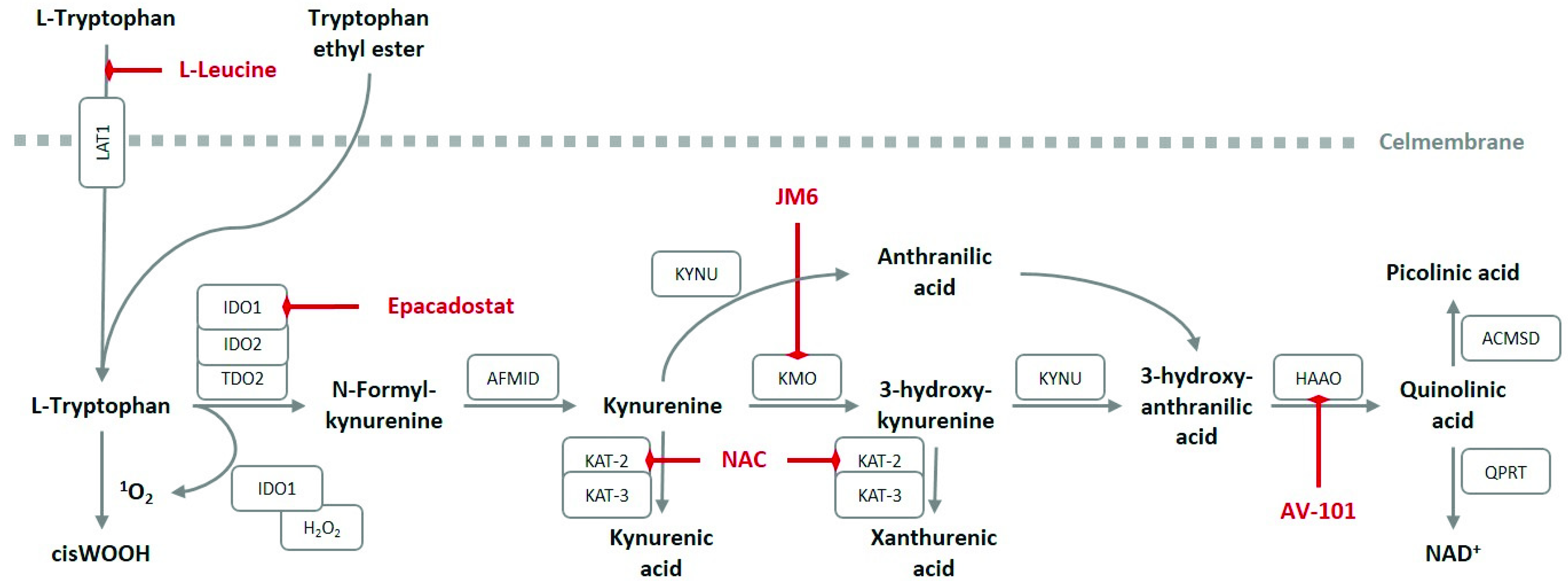
6. Pharmacological Interventions
7. Preclinical Models
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | Year | Dataset Title | Database and Identifier |
|---|---|---|---|
| Pique-Regi R, Romero R, Tarca AL, Sendler ED, Xu Y, Garcia-Flores V, Leng Y, Luca F, Hassan SS, Gomez-Lopez N. | 2019 | Single cell transcriptional signatures of the human placenta in term and preterm parturition. | NIH dbGAP repository, phs001886.v1.p1 |
| Vento-Tormo R, Efremova M, Botting RA, Turco MY, Vento-Tormo M, Meyer KB, Park JE, Stephenson E, Polański K, Goncalves A, Gardner L, Holmqvist S, Henriksson J, Zou A, Sharkey AM, Millar B, Innes B, Wood L, Wilbrey-Clark A, Payne RP, Ivarsson MA, Lisgo S, Filby A, Rowitch DH, Bulmer JN, Wright GJ, Stubbington MJT, Haniffa M, Moffett A, Teichmann SA. | 2018 | Single-cell reconstruction of the early maternal-fetal interface in humans. | ArrayExpress, E-MTAB-6701 |
| Tsang JCH, Vong JSL, Ji L, Poon LCY, Jiang P, Lui KO, Ni YB, To KF, Cheng YKY, Chiu RWK, Lo YMD. | 2017 | Integrative single-cell and cell-free plasma RNA transcriptomics elucidates placental cellular dynamics. | European Genome-Phenome Archive, EGAS00001002449 |
Acknowledgments
Conflicts of Interest
Glossary
| IDO | indoleamine 2,3-dioxygenase |
| TDO | tryptophan 2,3-dioxygenase |
| AFMID | arylformamidase |
| KAT-2 | aminoadipate aminotransferase |
| KAT-3 | kynurenine aminotransferase 3 |
| KMO | kynurenine 3-monooxygenase |
| KYNU | kynureninase |
| HAAO | 3-hydroxyanthranilate 3,4-dioxygenase |
| QPRT | quinolinate phosphoribosyltransferase |
| ACMSD | aminocarboxymuconate semialdehyde decarboxylase |
| NAD+ | oxidized nicotinamide adenine dinucleotide |
References
- Peters, J.C. Tryptophan nutrition and metabolism: An overview. Adv. Exp. Med. Biol. 1991, 294, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Ball, H.J.; Sanchez-Perez, A.; Weiser, S.; Austin, C.J.; Astelbauer, F.; Miu, J.; McQuillan, J.A.; Stocker, R.; Jermiin, L.S.; Hunt, N.H. Characterization of an indoleamine 2,3-dioxygenase-like protein found in humans and mice. Gene 2007, 396, 203–213. [Google Scholar] [CrossRef]
- Metz, R.; Duhadaway, J.B.; Kamasani, U.; Laury-Kleintop, L.; Muller, A.J.; Prendergast, G.C. Novel tryptophan catabolic enzyme IDO2 is the preferred biochemical target of the antitumor indoleamine 2,3-dioxygenase inhibitory compound D-1-methyl-tryptophan. Cancer Res. 2007, 67, 7082–7087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyland, E.; Williams, D.C. The metabolism of tryptophan. 2. The metabolism of tryptophan in patients suffering from cancer of the bladder. Biochem. J. 1956, 64, 578–582. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, G.C.; Malachowski, W.P.; DuHadaway, J.B.; Muller, A.J. Discovery of IDO1 Inhibitors: From Bench to Bedside. Cancer Res. 2017, 77, 6795–6811. [Google Scholar] [CrossRef] [Green Version]
- Munn, D.H.; Zhou, M.; Attwood, J.T.; Bondarev, I.; Conway, S.J.; Marshall, B.; Brown, C.; Mellor, A.L. Prevention of Allogeneic Fetal Rejection by Tryptophan Catabolism. Science 1998, 281, 1191–1193. [Google Scholar] [CrossRef]
- Pfefferkorn, E.R.; Rebhun, S.; Eckel, M. Characterization of an indoleamine 2,3-dioxygenase induced by gamma-interferon in cultured human fibroblasts. J. Interferon Res. 1986, 6, 267–279. [Google Scholar] [CrossRef]
- Ozaki, Y.; Edelstein, M.P.; Duch, D.S. The actions of interferon and antiinflammatory agents of induction of indoleamine 2,3-dioxygenase in human peripheral blood monocytes. Biochem. Biophys. Res. Commun. 1987, 144, 1147–1153. [Google Scholar] [CrossRef]
- Badawy, A.A.B. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. 2017, 10, 1178646917691938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, F.; Kuroiwa, T.; Takikawa, O.; Kido, R. Human indolylamine 2,3-dioxygenase. Its tissue distribution, and characterization of the placental enzyme. Biochem. J. 1985, 230, 635–638. [Google Scholar] [CrossRef]
- Munn, D.H.; Shafizadeh, E.; Attwood, J.T.; Bondarev, I.; Pashine, A.; Mellor, A.L. Inhibition of T cell proliferation by macrophage tryptophan catabolism. J. Exp. Med. 1999, 189, 1363–1372. [Google Scholar] [CrossRef]
- Badawy, A.A.; Namboodiri, A.M.; Moffett, J.R. The end of the road for the tryptophan depletion concept in pregnancy and infection. Clin. Sci. 2016, 130, 1327–1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffett, J.R.; Namboodiri, M.A. Tryptophan and the immune response. Immunol. Cell Biol. 2003, 81, 247–265. [Google Scholar] [CrossRef]
- Yeung, A.W.; Terentis, A.C.; King, N.J.; Thomas, S.R. Role of indoleamine 2,3-dioxygenase in health and disease. Clin. Sci. 2015, 129, 601–672. [Google Scholar] [CrossRef] [PubMed]
- Mondal, A.; Smith, C.; DuHadaway, J.B.; Sutanto-Ward, E.; Prendergast, G.C.; Bravo-Nuevo, A.; Muller, A.J. IDO1 is an Integral Mediator of Inflammatory Neovascularization. EBioMedicine 2016, 14, 74–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.; Chang, M.Y.; Parker, K.H.; Beury, D.W.; DuHadaway, J.B.; Flick, H.E.; Boulden, J.; Sutanto-Ward, E.; Soler, A.P.; Laury-Kleintop, L.D.; et al. IDO is a nodal pathogenic driver of lung cancer and metastasis development. Cancer Discov. 2012, 2, 722–735. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, H.; McKenzie, G.; Witting, P.K.; Stasch, J.P.; Hahn, M.; Changsirivathanathamrong, D.; Wu, B.J.; Ball, H.J.; Thomas, S.R.; et al. Kynurenine is an endothelium-derived relaxing factor produced during inflammation. Nat. Med. 2010, 16, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Stanley, C.P.; Maghzal, G.J.; Ayer, A.; Talib, J.; Giltrap, A.M.; Shengule, S.; Wolhuter, K.; Wang, Y.; Chadha, P.; Suarna, C.; et al. Singlet molecular oxygen regulates vascular tone and blood pressure in inflammation. Nature 2019, 566, 548–552. [Google Scholar] [CrossRef] [Green Version]
- Broekhuizen, M.; Klein, T.; Hitzerd, E.; de Rijke, Y.B.; Schoenmakers, S.; Sedlmayr, P.; Danser, A.H.J.; Merkus, D.; Reiss, I.K.M. l-Tryptophan-Induced Vasodilation Is Enhanced in Preeclampsia: Studies on Its Uptake and Metabolism in the Human Placenta. Hypertension 2020, 76, 184–194. [Google Scholar] [CrossRef]
- Xu, K.; Liu, G.; Fu, C. The Tryptophan Pathway Targeting Antioxidant Capacity in the Placenta. Oxidative Med. Cell. Longev. 2018, 2018, 1054797. [Google Scholar] [CrossRef]
- Christen, S.; Peterhans, E.; Stocker, R. Antioxidant activities of some tryptophan metabolites: Possible implication for inflammatory diseases. Proc. Natl. Acad. Sci. USA 1990, 87, 2506–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Liu, H.; Bai, M.; Gao, J.; Wu, X.; Yin, Y. Redox Properties of Tryptophan Metabolism and the Concept of Tryptophan Use in Pregnancy. Int. J. Mol. Sci. 2017, 18, 1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Zhang, M.; Ding, Y.; Wang, Q.; Zhang, W.; Song, P.; Zou, M.H. Activation of NAD(P)H oxidase by tryptophan-derived 3-hydroxykynurenine accelerates endothelial apoptosis and dysfunction in vivo. Circ. Res. 2014, 114, 480–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worton, S.A.; Greenwood, S.L.; Wareing, M.; Heazell, A.E.; Myers, J. The kynurenine pathway: A new target for treating maternal features of preeclampsia? Placenta 2019, 84, 44–49. [Google Scholar] [CrossRef]
- Sedlmayr, P.; Blaschitz, A.; Stocker, R. The role of placental tryptophan catabolism. Front. Immunol. 2014, 5, 230. [Google Scholar] [CrossRef] [Green Version]
- Kudo, Y.; Boyd, C.A. Human placental indoleamine 2,3-dioxygenase: Cellular localization and characterization of an enzyme preventing fetal rejection. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1500, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Karahoda, R.; Abad, C.; Horackova, H.; Kastner, P.; Zaugg, J.; Cerveny, L.; Kucera, R.; Albrecht, C.; Staud, F. Dynamics of Tryptophan Metabolic Pathways in Human Placenta and Placental-Derived Cells: Effect of Gestation Age and Trophoblast Differentiation. Front. Cell Dev. Biol. 2020, 8, 574034. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, S.; Eguchi, K.; Yonezawa, M.; Sekiba, K. Localization and developmental change of indoleamine 2,3-dioxygenase activity in the human placenta. Acta Med. Okayama 1991, 45, 135–139. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Zhou, C.; Wei, Y.Y.; Li, H.H.; Lei, W.; Boeldt, D.S.; Wang, K.; Zheng, J. Differential Distribution of Tryptophan-Metabolites in Fetal and Maternal Circulations During Normotensive and Preeclamptic Pregnancies. Reprod. Sci. 2021. [Google Scholar] [CrossRef]
- Holm, M.B.; Bastani, N.E.; Holme, A.M.; Zucknick, M.; Jansson, T.; Refsum, H.; Mørkrid, L.; Blomhoff, R.; Henriksen, T.; Michelsen, T.M. Uptake and release of amino acids in the fetal-placental unit in human pregnancies. PLoS ONE 2017, 12, e0185760. [Google Scholar] [CrossRef] [PubMed]
- Murthi, P.; Wallace, E.M.; Walker, D.W. Altered placental tryptophan metabolic pathway in human fetal growth restriction. Placenta 2017, 52, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Schulpis, K.H.; Vlachos, G.D.; Karikas, G.A.; Papakonstantinou, E.D.; Vlachos, D.G.; Papassotiriou, I.; Antsaklis, A.; Tsakiris, S. The effect of the mode of delivery on maternal-neonatal interleukin-6, biogenic amine and their precursor amino acid concentrations. Clin. Chem. Lab. Med. 2008, 46, 1624–1630. [Google Scholar] [CrossRef]
- Camelo, J.S., Jr.; Jorge, S.M.; Martinez, F.E. Amino acid composition of parturient plasma, the intervillous space of the placenta and the umbilical vein of term newborn infants. Braz. J. Med. Biol. Res. 2004, 37, 711–717. [Google Scholar] [CrossRef] [Green Version]
- Morita, I.; Kawamoto, M.; Yoshida, H. Difference in the concentration of tryptophan metabolites between maternal and umbilical foetal blood. J. Chromatogr. B Biomed. Sci. Appl. 1992, 576, 334–339. [Google Scholar] [CrossRef]
- Kamimura, S.; Eguchi, K.; Sekiba, K. Tryptophan and its metabolite concentrations in human plasma and breast milk during the perinatal period. Acta Med. Okayama 1991, 45, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Zanardo, V.; Casson, L.; Costa, C.; Biasiolo, M.; Allegri, G.; Vicenti, E.; Grella, P. Levels of Total and Free Tryptophan in Maternal and in Umbilical Blood of Infants Born by Vaginal Delivery and by Elective Cesarean Section. Neonatology 1985, 48, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Tricklebank, M.D.; Pickard, F.J.; de Souza, S.W. Free and bound tryptophan in human plasma during the perinatal period. Acta Pædiatrica 1979, 68, 199–204. [Google Scholar] [CrossRef]
- Takikawa, O.; Kuroiwa, T.; Yamazaki, F.; Kido, R. Mechanism of interferon-gamma action. Characterization of indoleamine 2,3-dioxygenase in cultured human cells induced by interferon-gamma and evaluation of the enzyme-mediated tryptophan degradation in its anticellular activity. J. Biol. Chem. 1988, 263, 2041–2048. [Google Scholar] [CrossRef]
- Kudo, Y.; Koh, I.; Sugimoto, J. Localization of Indoleamine 2,3-Dioxygenase-1 and Indoleamine 2,3-Dioxygenase-2 at the Human Maternal-Fetal Interface. Int. J. Tryptophan Res. 2020, 13, 1178646920984163. [Google Scholar] [CrossRef]
- Murthy, G.G.; Prideaux, M.A.; Armstrong, M.; Kenney, H.M.; Latchney, S.E.; Susiarjo, M.; Murphy, S.P. Characterization of the temporal, cell-specific and interferon-inducible patterns of indoleamine 2,3 dioxygenase 1 (IDO1) expression in the human placenta across gestation. Placenta 2021, 115, 129–138. [Google Scholar] [CrossRef]
- Santoso, D.I.; Rogers, P.; Wallace, E.M.; Manuelpillai, U.; Walker, D.; Subakir, S.B. Localization of indoleamine 2,3-dioxygenase and 4-hydroxynonenal in normal and pre-eclamptic placentae. Placenta 2002, 23, 373–379. [Google Scholar] [CrossRef]
- Sedlmayr, P.; Blaschitz, A.; Wintersteiger, R.; Semlitsch, M.; Hammer, A.; MacKenzie, C.R.; Walcher, W.; Reich, O.; Takikawa, O.; Dohr, G. Localization of indoleamine 2,3-dioxygenase in human female reproductive organs and the placenta. Mol. Hum. Reprod. 2002, 8, 385–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaschitz, A.; Gauster, M.; Fuchs, D.; Lang, I.; Maschke, P.; Ulrich, D.; Karpf, E.; Takikawa, O.; Schimek, M.G.; Dohr, G.; et al. Vascular Endothelial Expression of Indoleamine 2,3-Dioxygenase 1 Forms a Positive Gradient towards the Feto-Maternal Interface. PLoS ONE 2011, 6, e21774. [Google Scholar] [CrossRef] [Green Version]
- Kudo, Y.; Boyd, C.A.R.; Spyropoulou, I.; Redman, C.W.G.; Takikawa, O.; Katsuki, T.; Hara, T.; Ohama, K.; Sargent, I.L. Indoleamine 2,3-dioxygenase: Distribution and function in the developing human placenta. J. Reprod. Immunol. 2004, 61, 87–98. [Google Scholar] [CrossRef]
- Ligam, P.; Manuelpillai, U.; Wallace, E.M.; Walker, D. Localisation of indoleamine 2,3-dioxygenase and kynurenine hydroxylase in the human placenta and decidua: Implications for role of the kynurenine pathway in pregnancy. Placenta 2005, 26, 498–504. [Google Scholar] [CrossRef]
- Hönig, A.; Rieger, L.; Kapp, M.; Sütterlin, M.; Dietl, J.; Kämmerer, U. Indoleamine 2,3-dioxygenase (IDO) expression in invasive extravillous trophoblast supports role of the enzyme for materno-fetal tolerance. J. Reprod. Immunol. 2004, 61, 79–86. [Google Scholar] [CrossRef]
- Iwahashi, N.; Yamamoto, M.; Nanjo, S.; Toujima, S.; Minami, S.; Ino, K. Downregulation of indoleamine 2, 3-dioxygenase expression in the villous stromal endothelial cells of placentas with preeclampsia. J. Reprod. Immunol. 2017, 119, 54–60. [Google Scholar] [CrossRef]
- Manuelpillai, U.; Ligam, P.; Smythe, G.; Wallace, E.M.; Hirst, J.; Walker, D.W. Identification of kynurenine pathway enzyme mRNAs and metabolites in human placenta: Up-regulation by inflammatory stimuli and with clinical infection. Am. J. Obstet. Gynecol. 2005, 192, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Vento-Tormo, R.; Efremova, M.; Botting, R.A.; Turco, M.Y.; Vento-Tormo, M.; Meyer, K.B.; Park, J.E.; Stephenson, E.; Polański, K.; Goncalves, A.; et al. Single-cell reconstruction of the early maternal-fetal interface in humans. Nature 2018, 563, 347–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pique-Regi, R.; Romero, R.; Tarca, A.L.; Sendler, E.D.; Xu, Y.; Garcia-Flores, V.; Leng, Y.; Luca, F.; Hassan, S.S.; Gomez-Lopez, N. Single cell transcriptional signatures of the human placenta in term and preterm parturition. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.C.H.; Vong, J.S.L.; Ji, L.; Poon, L.C.Y.; Jiang, P.; Lui, K.O.; Ni, Y.-B.; To, K.F.; Cheng, Y.K.Y.; Chiu, R.W.K.; et al. Integrative single-cell and cell-free plasma RNA transcriptomics elucidates placental cellular dynamics. Proc. Natl. Acad. Sci. USA 2017, 114, E7786–E7795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolahi, K.S.; Valent, A.M.; Thornburg, K.L. Cytotrophoblast, Not Syncytiotrophoblast, Dominates Glycolysis and Oxidative Phosphorylation in Human Term Placenta. Sci. Rep. 2017, 7, 42941. [Google Scholar] [CrossRef] [Green Version]
- Abbassi-Ghanavati, M.; Greer, L.G.; Cunningham, F.G. Pregnancy and laboratory studies: A reference table for clinicians. Obstet. Gynecol. 2009, 114, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.B. Tryptophan metabolism, disposition and utilization in pregnancy. Biosci. Rep. 2015, 35, e00261. [Google Scholar] [CrossRef]
- Fairweather, D.V.I. Changes in levels of serum non-esterified fatty acid and blood glucose in pregnancy. BJOG Int. J. Obstet. Gynaecol. 1971, 78, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Plumptre, L.; Brosnan, M.E.; Pongnopparat, T.; Masih, S.P.; Visentin, C.E.; Berger, H.; Lamers, Y.; Caudill, M.A.; Malysheva, O.V.; et al. Formate concentrations in maternal plasma during pregnancy and in cord blood in a cohort of pregnant Canadian women: Relations to genetic polymorphisms and plasma metabolites. Am. J. Clin. Nutr. 2019, 110, 1131–1137. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2020, 49, D480–D489. [Google Scholar] [CrossRef]
- Abad, C.; Karahoda, R.; Kastner, P.; Portillo, R.; Horackova, H.; Kucera, R.; Nachtigal, P.; Staud, F. Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit During Gestation. Int. J. Mol. Sci. 2020, 21, 7578. [Google Scholar] [CrossRef]
- Vumma, R.; Johansson, J.; Lewander, T.; Venizelos, N. Tryptophan transport in human fibroblast cells-a functional characterization. Int. J. Tryptophan Res. 2011, 4, 19–27. [Google Scholar] [CrossRef]
- Kudo, Y.; Boyd, C.A. Characterisation of L-tryptophan transporters in human placenta: A comparison of brush border and basal membrane vesicles. J. Physiol. 2001, 531, 405–416. [Google Scholar] [CrossRef]
- Verrey, F. System L: Heteromeric exchangers of large, neutral amino acids involved in directional transport. Pflug. Arch. 2003, 445, 529–533. [Google Scholar] [CrossRef]
- Seymour, R.L.; Ganapathy, V.; Mellor, A.L.; Munn, D.H. A high-affinity, tryptophan-selective amino acid transport system in human macrophages. J. Leukoc. Biol. 2006, 80, 1320–1327. [Google Scholar] [CrossRef] [Green Version]
- Bhutia, Y.D.; Babu, E.; Ganapathy, V. Interferon-γ induces a tryptophan-selective amino acid transporter in human colonic epithelial cells and mouse dendritic cells. Biochim. Biophys. Acta (BBA)-Biomembr. 2015, 1848, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudo, Y.; Boyd, C.A. The role of L-tryptophan transport in L-tryptophan degradation by indoleamine 2,3-dioxygenase in human placental explants. J. Physiol. 2001, 531, 417–423. [Google Scholar] [CrossRef]
- Sinclair, L.V.; Neyens, D.; Ramsay, G.; Taylor, P.M.; Cantrell, D.A. Single cell analysis of kynurenine and System L amino acid transport in T cells. Nat. Commun. 2018, 9, 1981. [Google Scholar] [CrossRef] [PubMed]
- Scalise, M.; Pochini, L.; Console, L.; Losso, M.A.; Indiveri, C. The Human SLC1A5 (ASCT2) Amino Acid Transporter: From Function to Structure and Role in Cell Biology. Front. Cell Dev. Biol. 2018, 6, 96. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, H.; Hasegawa, K.; Suzuki, M.; Achiwa, Y.; Kato, T.; Saito, K.; Kurahashi, H.; Udagawa, Y. Mouse model for allogeneic immune reaction against fetus recapitulates human pre-eclampsia. J. Obstet. Gynaecol. Res. 2008, 34, 1–6. [Google Scholar] [CrossRef]
- Santillan, M.K.; Pelham, C.J.; Ketsawatsomkron, P.; Santillan, D.A.; Davis, D.R.; Devor, E.J.; Gibson-Corley, K.N.; Scroggins, S.M.; Grobe, J.L.; Yang, B.; et al. Pregnant mice lacking indoleamine 2,3-dioxygenase exhibit preeclampsia phenotypes. Physiol. Rep. 2015, 3, e12257. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A.B. The tryptophan utilization concept in pregnancy. Obstet. Gynecol. Sci. 2014, 57, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Mezrich, J.D.; Fechner, J.H.; Zhang, X.; Johnson, B.P.; Burlingham, W.J.; Bradfield, C.A. An Interaction between Kynurenine and the Aryl Hydrocarbon Receptor Can Generate Regulatory T Cells. J. Immunol. 2010, 185, 3190–3198. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.T.; Kimura, A.; Nakahama, T.; Chinen, I.; Masuda, K.; Nohara, K.; Fujii-Kuriyama, Y.; Kishimoto, T. Aryl hydrocarbon receptor negatively regulates dendritic cell immunogenicity via a kynurenine-dependent mechanism. Proc. Natl. Acad. Sci. USA 2010, 107, 19961–19966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, C.F.A.; Goth, S.R.; Dong, B.; Pessah, I.N.; Matsumura, F. Aryl hydrocarbon receptor signaling mediates expression of indoleamine 2,3-dioxygenase. Biochem. Biophys. Res. Commun. 2008, 375, 331–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Bianchi, R.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T cell apoptosis by tryptophan catabolism. Cell Death Differ. 2002, 9, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhang, G.-X.; Gran, B.; Fallarino, F.; Yu, S.; Li, H.; Cullimore, M.L.; Rostami, A.; Xu, H. IDO Upregulates Regulatory T Cells via Tryptophan Catabolite and Suppresses Encephalitogenic T Cell Responses in Experimental Autoimmune Encephalomyelitis. J. Immunol. 2010, 185, 5953–5961. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-S.; Lee, S.-M.; Kim, M.-K.; Park, S.-G.; Choi, I.-W.; Choi, I.; Joo, Y.-D.; Park, S.-J.; Kang, S.-W.; Seo, S.-K. The tryptophan metabolite 3-hydroxyanthranilic acid suppresses T cell responses by inhibiting dendritic cell activation. Int. Immunopharmacol. 2013, 17, 721–726. [Google Scholar] [CrossRef]
- Wirthgen, E.; Leonard, A.K.; Scharf, C.; Domanska, G. The Immunomodulator 1-Methyltryptophan Drives Tryptophan Catabolism Toward the Kynurenic Acid Branch. Front. Immunol. 2020, 11, 313. [Google Scholar] [CrossRef] [PubMed]
- Lewis, H.C.; Chinnadurai, R.; Bosinger, S.E.; Galipeau, J. The IDO inhibitor 1-methyl tryptophan activates the aryl hydrocarbon receptor response in mesenchymal stromal cells. Oncotarget 2017, 8, 91914–91927. [Google Scholar] [CrossRef] [Green Version]
- Agaugué, S.; Perrin-Cocon, L.; Coutant, F.; André, P.; Lotteau, V. 1-Methyl-Tryptophan Can Interfere with TLR Signaling in Dendritic Cells Independently of IDO Activity. J. Immunol. 2006, 177, 2061–2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallotta, M.T.; Rossini, S.; Suvieri, C.; Coletti, A.; Orabona, C.; Macchiarulo, A.; Volpi, C.; Grohmann, U. Indoleamine 2,3-dioxygenase 1 (IDO1): An up-to-date overview of an eclectic immunoregulatory enzyme. FEBS J. 2021. [Google Scholar] [CrossRef]
- Pallotta, M.T.; Orabona, C.; Volpi, C.; Vacca, C.; Belladonna, M.L.; Bianchi, R.; Servillo, G.; Brunacci, C.; Calvitti, M.; Bicciato, S.; et al. Indoleamine 2,3-dioxygenase is a signaling protein in long-term tolerance by dendritic cells. Nat. Immunol. 2011, 12, 870–878. [Google Scholar] [CrossRef] [Green Version]
- Iacono, A.; Pompa, A.; De Marchis, F.; Panfili, E.; Greco, F.A.; Coletti, A.; Orabona, C.; Volpi, C.; Belladonna, M.L.; Mondanelli, G.; et al. Class IA PI3Ks regulate subcellular and functional dynamics of IDO1. EMBO Rep. 2020, 21, e49756. [Google Scholar] [CrossRef] [PubMed]
- Hitzerd, E.; Broekhuizen, M.; Neuman, R.I.; Colafella, K.M.M.; Merkus, D.; Schoenmakers, S.; Simons, S.H.P.; Reiss, I.K.M.; Danser, A.H.J. Human Placental Vascular Reactivity in Health and Disease: Implications for the Treatment of Pre-eclampsia. Curr. Pharm. Des. 2019, 25, 505–527. [Google Scholar] [CrossRef]
- Zardoya-Laguardia, P.; Blaschitz, A.; Hirschmugl, B.; Lang, I.; Herzog, S.A.; Nikitina, L.; Gauster, M.; Häusler, M.; Cervar-Zivkovic, M.; Karpf, E.; et al. Endothelial indoleamine 2,3-dioxygenase-1 regulates the placental vascular tone and is deficient in intrauterine growth restriction and pre-eclampsia. Sci. Rep. 2018, 8, 5488. [Google Scholar] [CrossRef]
- Worton, S.A.; Pritchard, H.A.; Greenwood, S.L.; Alakrawi, M.; Heazell, A.E.; Wareing, M.; Greenstein, A.S.; Myers, J.E. Kynurenine Relaxes Arteries of Normotensive Women and Those with Preeclampsia. Circ. Res. 2021, 128, 1679–1693. [Google Scholar] [CrossRef]
- Werner, E.R.; Werner-Felmayer, G. Substrate and cofactor requirements of indoleamine 2,3-dioxygenase in interferon-gamma-treated cells: Utilization of oxygen rather than superoxide. Curr. Drug Metab. 2007, 8, 201–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horgan, R.P.; Broadhurst, D.I.; Dunn, W.B.; Brown, M.; Heazell, A.E.; Kell, D.B.; Baker, P.N.; Kenny, L.C. Changes in the metabolic footprint of placental explant-conditioned medium cultured in different oxygen tensions from placentas of small for gestational age and normal pregnancies. Placenta 2010, 31, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Hirata, F.; Hayaishi, O. Studies on indoleamine 2,3-dioxygenase. I. Superoxide anion as substrate. J. Biol. Chem. 1975, 250, 5960–5966. [Google Scholar] [CrossRef]
- Weiss, G.; Diez-Ruiz, A.; Murr, C.; Theur, I.; Fuchs, D. Tryptophan Metabolites as Scavengers of Reactive Oxygen and Chlorine Species. Pteridines 2002, 13, 140–143. [Google Scholar] [CrossRef]
- Lima, V.L.; Dias, F.; Nunes, R.D.; Pereira, L.O.; Santos, T.S.; Chiarini, L.B.; Ramos, T.D.; Silva-Mendes, B.J.; Perales, J.; Valente, R.H.; et al. The antioxidant role of xanthurenic acid in the Aedes aegypti midgut during digestion of a blood meal. PLoS ONE 2012, 7, e38349. [Google Scholar] [CrossRef] [Green Version]
- Lugo-Huitrón, R.; Blanco-Ayala, T.; Ugalde-Muñiz, P.; Carrillo-Mora, P.; Pedraza-Chaverrí, J.; Silva-Adaya, D.; Maldonado, P.D.; Torres, I.; Pinzón, E.; Ortiz-Islas, E.; et al. On the antioxidant properties of kynurenic acid: Free radical scavenging activity and inhibition of oxidative stress. Neurotoxicol. Teratol. 2011, 33, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Goda, K.; Hamane, Y.; Kishimoto, R.; Ogishi, Y. Radical scavenging properties of tryptophan metabolites. Estimation of their radical reactivity. Adv. Exp. Med. Biol. 1999, 467, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.Y.-H.; Oldham, W.M.; He, H.; Wang, R.; Mulhern, R.; Handy, D.E.; Loscalzo, J. Interferon-gamma Impairs Human Coronary Artery Endothelial Glucose Metabolism via Tryptophan Catabolism and Activates Fatty Acid Oxidation. Circulation 2021. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Liu, S.; Lian, R.; Huang, C.; Li, Y.; Chen, L.; Zeng, Y. Abnormal Expression of Indoleamine 2, 3-Dioxygenase in Human Recurrent Miscarriage. Reprod. Sci. 2020, 27, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Ban, Y.; Chang, Y.; Dong, B.; Kong, B.; Qu, X. Indoleamine 2,3-dioxygenase levels at the normal and recurrent spontaneous abortion fetal-maternal interface. J. Int. Med. Res. 2013, 41, 1135–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miwa, N.; Hayakawa, S.; Miyazaki, S.; Myojo, S.; Sasaki, Y.; Sakai, M.; Takikawa, O.; Saito, S. IDO expression on decidual and peripheral blood dendritic cells and monocytes/macrophages after treatment with CTLA-4 or interferon-γ increase in normal pregnancy but decrease in spontaneous abortion. Mol. Hum. Reprod. 2006, 11, 865–870. [Google Scholar] [CrossRef] [Green Version]
- Karahoda, R.; Robles, M.; Marushka, J.; Stranik, J.; Abad, C.; Horackova, H.; Tebbens, J.D.; Vaillancourt, C.; Kacerovsky, M.; Staud, F. Prenatal inflammation as a link between placental expression signature of tryptophan metabolism and preterm birth. Hum. Mol. Genet. 2021. [Google Scholar] [CrossRef]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. Hypertensive Disorders of Pregnancy. Hypertension 2018, 72, 24–43. [Google Scholar] [CrossRef] [Green Version]
- Steegers, E.A.P.; von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Pre-eclampsia. Lancet 2010, 376, 631–644. [Google Scholar] [CrossRef]
- Rana, S.; Lemoine, E.; Granger, J.P.; Karumanchi, S.A. Preeclampsia. Circ. Res. 2019, 124, 1094–1112. [Google Scholar] [CrossRef]
- Kudo, Y.; Boyd, C.A.; Sargent, I.L.; Redman, C.W. Decreased tryptophan catabolism by placental indoleamine 2,3-dioxygenase in preeclampsia. Am. J. Obstet. Gynecol. 2003, 188, 719–726. [Google Scholar] [CrossRef]
- Dunn, W.B.; Brown, M.; Worton, S.A.; Crocker, I.P.; Broadhurst, D.; Horgan, R.; Kenny, L.C.; Baker, P.N.; Kell, D.B.; Heazell, A.E. Changes in the metabolic footprint of placental explant-conditioned culture medium identifies metabolic disturbances related to hypoxia and pre-eclampsia. Placenta 2009, 30, 974–980. [Google Scholar] [CrossRef]
- Keaton, S.A.; Heilman, P.; Bryleva, E.Y.; Madaj, Z.; Krzyzanowski, S.; Grit, J.; Miller, E.S.; Jälmby, M.; Kalapotharakos, G.; Racicot, K.; et al. Altered Tryptophan Catabolism in Placentas from Women With Pre-eclampsia. Int. J. Tryptophan Res. 2019, 12, 1178646919840321. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Okatani, Y.; Sagara, Y. Serotonin Metabolism in the Fetus in Preeclampsia†. Asia-Ocean. J. Obstet. Gynaecol. 1994, 20, 77–86. [Google Scholar] [CrossRef]
- Nilsen, R.M.; Bjørke-Monsen, A.-L.; Midttun, Ø.; Nygård, O.; Pedersen, E.R.; Ulvik, A.; Magnus, P.; Gjessing, H.K.; Vollset, S.E.; Ueland, P.M. Maternal Tryptophan and Kynurenine Pathway Metabolites and Risk of Preeclampsia. Obstet. Gynecol. 2012, 119, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Sano, M.; Ferchaud-Roucher, V.; Kaeffer, B.; Poupeau, G.; Castellano, B.; Darmaun, D. Maternal and fetal tryptophan metabolism in gestating rats: Effects of intrauterine growth restriction. Amino Acids 2016, 48, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Murtaza, G.; Metwally, E.; Yang, H.; Kalhoro, M.S.; Kalhoro, D.H.; Chughtai, M.I.; Yin, Y. Role of Dietary Amino Acids and Nutrient Sensing System in Pregnancy Associated Disorders. Front. Pharmacol. 2020, 11, 586979. [Google Scholar] [CrossRef]
- Qiu, S.; Fang, Z.; Wu, D.; Lin, Y.; Che, L. Tryptophan Supplements Promote Pregnancy Success in Mice Challenged with Pseudorabies Virus (PRV) by Regulating the Expression of Systemic Cytokines, Immunoglobulins, PRV-Specific Protein Profiles, and Toll-Like Receptors. J. Med. Food 2011, 14, 857–865. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lin, I.C.; Yu, H.R.; Huang, L.T.; Tiao, M.M.; Tain, Y.L. Maternal Tryptophan Supplementation Protects Adult Rat Offspring against Hypertension Programmed by Maternal Chronic Kidney Disease: Implication of Tryptophan-Metabolizing Microbiome and Aryl Hydrocarbon Receptor. Int. J. Mol. Sci. 2020, 21, 4552. [Google Scholar] [CrossRef]
- Jo, J.H.; Lee, J.S.; Nejad, J.G.; Kim, W.S.; Moon, J.O.; Lee, H.G. Effects of Dietary Supplementation of Acetate and L-Tryptophan Conjugated Bypass Amino Acid on Productivity of Pre- and Post-Partum Dairy Cows and Their Offspring. Animals 2021, 11, 1726. [Google Scholar] [CrossRef]
- Ma, H.; Zhang, W.; Zhu, X.; Song, W.; Liu, J.; Jia, Z. Effects of rumen-protected tryptophan on growth performance, fibre characteristics, nutrient utilization and plasma essential amino acids in Cashmere goats during the cashmere slow growth period. Livest. Sci. 2010, 131, 227–233. [Google Scholar] [CrossRef]
- Forrest, C.M.; Mackay, G.M.; Stoy, N.; Egerton, M.; Christofides, J.; Stone, T.W.; Darlington, L.G. Tryptophan loading induces oxidative stress. Free Radic. Res. 2004, 38, 1167–1171. [Google Scholar] [CrossRef]
- Xu, K.; Fu, Y.; Gao, H.; Bai, M.; Liu, H.; Duan, Y. L-Tryptophan activates the aryl hydrocarbon receptor and induces cell cycle arrest in porcine trophectoderm cells. Theriogenology 2021, 171, 137–146. [Google Scholar] [CrossRef]
- Tchirikov, M.; Zhumadilov, Z.S.; Bapayeva, G.; Bergner, M.; Entezami, M. The effect of intraumbilical fetal nutrition via a subcutaneously implanted port system on amino acid concentration by severe IUGR human fetuses. J. Perinat. Med. 2017, 45, 227–236. [Google Scholar] [CrossRef]
- Jonas, A.J.; Butler, I.J. Circumvention of defective neutral amino acid transport in Hartnup disease using tryptophan ethyl ester. J. Clin. Investig. 1989, 84, 200–204. [Google Scholar] [CrossRef]
- Komrokji, R.S.; Wei, S.; Mailloux, A.W.; Zhang, L.; Padron, E.; Sallman, D.; Lancet, J.E.; Tinsley, S.; Nardelli, L.A.; Pinilla-Ibarz, J.; et al. A Phase II Study to Determine the Safety and Efficacy of the Oral Inhibitor of Indoleamine 2,3-Dioxygenase (IDO) Enzyme INCB024360 in Patients with Myelodysplastic Syndromes. Clin. Lymphoma Myeloma Leuk. 2019, 19, 157–161. [Google Scholar] [CrossRef]
- Maddison, D.C.; Giorgini, F. The kynurenine pathway and neurodegenerative disease. Semin. Cell Dev. Biol. 2015, 40, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Ayala, T.; Sathyasaikumar, K.V.; Uys, J.D.; Pérez-de-la-Cruz, V.; Pidugu, L.S.; Schwarcz, R. N-Acetylcysteine Inhibits Kynurenine Aminotransferase II. Neuroscience 2020, 444, 160–169. [Google Scholar] [CrossRef]
- Ayala, T.B.; Ortega, D.R.; Rodríguez, P.O.; Pineda, B.; de la Cruz, G.P.; Esquivel, D.G.; Schwarcz, R.; Sathyasaikumar, K.V.; Anguiano, A.J.; de la Cruz, V.P. Subchronic N-acetylcysteine Treatment Decreases Brain Kynurenic Acid Levels and Improves Cognitive Performance in Mice. Antioxidants 2021, 10, 147. [Google Scholar] [CrossRef]
- Walker, A.K.; Wing, E.E.; Banks, W.A.; Dantzer, R. Leucine competes with kynurenine for blood-to-brain transport and prevents lipopolysaccharide-induced depression-like behavior in mice. Mol. Psychiatry 2019, 24, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Zwilling, D.; Huang, S.Y.; Sathyasaikumar, K.V.; Notarangelo, F.M.; Guidetti, P.; Wu, H.Q.; Lee, J.; Truong, J.; Andrews-Zwilling, Y.; Hsieh, E.W.; et al. Kynurenine 3-monooxygenase inhibition in blood ameliorates neurodegeneration. Cell 2011, 145, 863–874. [Google Scholar] [CrossRef] [Green Version]
- Rojewska, E.; Piotrowska, A.; Makuch, W.; Przewlocka, B.; Mika, J. Pharmacological kynurenine 3-monooxygenase enzyme inhibition significantly reduces neuropathic pain in a rat model. Neuropharmacology 2016, 102, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Ramakrishnan, N.; Vo-Le, B.; Vo-Le, B.; Smith, M.A.; Iqbal, T.; Swann, A.C.; Mathew, S.J.; Lijffijt, M. A randomized cross-over trial to define neurophysiological correlates of AV-101 N-methyl-d-aspartate receptor blockade in healthy veterans. Neuropsychopharmacology 2021, 46, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Park, L.T.; Kadriu, B.; Gould, T.D.; Zanos, P.; Greenstein, D.; Evans, J.W.; Yuan, P.; Farmer, C.A.; Oppenheimer, M.; George, J.M.; et al. A Randomized Trial of the N-Methyl-d-Aspartate Receptor Glycine Site Antagonist Prodrug 4-Chlorokynurenine in Treatment-Resistant Depression. Int. J. Neuropsychopharmacol. 2020, 23, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Meyers, C.D.; Kamanna, V.S.; Kashyap, M.L. Niacin therapy in atherosclerosis. Curr. Opin. Lipidol. 2004, 15, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.J.; Yan, L.; Charlton, F.; Witting, P.; Barter, P.J.; Rye, K.-A. Evidence That Niacin Inhibits Acute Vascular Inflammation and Improves Endothelial Dysfunction Independent of Changes in Plasma Lipids. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 968–975. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Toné, S.; Takikawa, O.; Kubo, T.; Kohno, I.; Minatogawa, Y. Expression of indoleamine 2,3-dioxygenase and tryptophan 2,3-dioxygenase in early concepti. Biochem. J. 2001, 355, 425–429. [Google Scholar] [CrossRef]





| Measured Metabolite | Reference | Maternal Vein | Umbilical Vein | Umbilical Artery |
|---|---|---|---|---|
| Tryptophan (μmol/L) | Zhao (2021) [29] | 34.5 | 64.1 | ∙ |
| Holm (2017) [30] | 41.1 | 88.3 | 83 | |
| Murthi (2017) [31] | 28.5 | 60.2 | 58.9 | |
| Schulpis (2008) [32] | 28 | 26 | ∙ | |
| Camelo (2004) [33] | 29 | 40 | ∙ | |
| Morita (1992) [34] | 35.1 | 75.9 | 68.9 | |
| Kamimura (1991) [35] | 39.6 | 82.6 | 83.4 | |
| Zanardo (1985) [36] | 7.32 | 10.14 | ∙ | |
| Tricklebank (1979) [37] | 53.3 | 101.47 | ∙ | |
| Free tryptophan (μmol/L) | Kamimura (1991) [35] | 7.5 | 17.5 | 16.4 |
| Tricklebank (1979) [37] | 12.18 | ∙ | 20.46 | |
| N-Formyl-kynurenine (μmol/L) | Zhao (2021) [29] | 0.18 | 0.40 | ∙ |
| Kynurenine (μmol/L) | Zhao (2021) [29] | 0.85 | 1.78 | ∙ |
| Murthi (2017) [31] | 0.9 | 4 | 4 | |
| Morita (1992) [34] | 0.91 | 4.96 | 4.59 | |
| Kamimura (1991) [35] | 1.17 | 4.12 | 4.25 | |
| Kynurenic acid (nmol/L) | Zhao (2021) [29] | 23 | 22 | ∙ |
| Kamimura (1991) [35] | 20.5 | 38.1 | 39.8 | |
| Murthi (2017) [31] | 100 | 300 | 360 | |
| 3-Hydroxykynurenine (nmol/L) | Zhao (2021) [29] | 196 | 272 | ∙ |
| Xanthurenic acid (nmol/L) | Zhao (2021) [29] | 118 | 315 | ∙ |
| Kamimura (1991) [35] | 188 | 295 | 258 | |
| Anthranilic acid (nmol/L) | Kamimura (1991) [35] | 413 | 387 | 432 |
| 3-Hydroxyanthranilic acid (nmol/L) | Zhao (2021) [29] | 287 | 448 | ∙ |
| Murthi (2017) [31] | 300 | 300 | 340 | |
| Morita (1992) [34] | N.D. | 270 | 260 | |
| Kamimura (1991) [35] | 12 | 527 | 702 | |
| Quinolinic acid (nmol/L) | Zhao (2021) [29] | 1333 | 2343 | ∙ |
| Murthi (2017) [31] | 600 | 1500 | 1700 | |
| Picolinic acid (nmol/L) | Zhao (2021) [29] | 223 | 287 | ∙ |
| Murthi (2017) [31] | 500 | 1500 | 1400 |
| First Trimester | Kamimura [28] (1991) | Santoso [41] (2002) | Sedlmayr [42] (2002) | Hönig [46] (2004) | Kudoku [26] (2004) | Ligam [45] (2005) | Blaschitz [43] (2011) | Iwahashi [47] (2017) | Kudo [39] (2020) | Murthy [40] (2021) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Antibody against | IDO 1 | IDO 1 | IDO 1 | IDO 2 | IDO 1 | IDO 1 | IDO 1 | IDO 1 | IDO1 3 | IDO2 4 | IDO1 5 |
| Placenta | |||||||||||
| Endothelial cells | nm | + | + | + | nm | ||||||
| Macrophages | nm | + | nm | ||||||||
| Cytotrophoblasts | nm | + | nm | ||||||||
| Syncythiotrophoblasts | ++ | nm | ++ | ++ | nm | ++ | |||||
| Stromal cells | nm | + | + | nm | |||||||
| Decidua | |||||||||||
| Endothelial cells | nm | + | + | + | nm | ||||||
| Epithelial glandular cells | nm | ++ | + | ++ | + | + | nm | ++ | |||
| Extravillous trophoblasts | nm | ++ | ++ | nm | |||||||
| Stromal cells | nm | + | + | nm | |||||||
| Term | |||||||||||
| Placenta | |||||||||||
| Endothelial cells | ++ | + | + | ++ | ++ | ++ | ++ | ++ | ++ | ||
| Macrophages | ++ | ++ | + | ||||||||
| Cytotrophoblasts | + | ||||||||||
| Syncythiotrophoblasts | ++ | + | ++ | + | |||||||
| Stromal cells | |||||||||||
| Decidua | |||||||||||
| Endothelial cells | + | ++ | + | ||||||||
| Epithelial glandular cells | + | ||||||||||
| Extravillous trophoblasts | ++ | + | + | ||||||||
| Stromal cells | |||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broekhuizen, M.; Danser, A.H.J.; Reiss, I.K.M.; Merkus, D. The Function of the Kynurenine Pathway in the Placenta: A Novel Pharmacotherapeutic Target? Int. J. Environ. Res. Public Health 2021, 18, 11545. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182111545
Broekhuizen M, Danser AHJ, Reiss IKM, Merkus D. The Function of the Kynurenine Pathway in the Placenta: A Novel Pharmacotherapeutic Target? International Journal of Environmental Research and Public Health. 2021; 18(21):11545. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182111545
Chicago/Turabian StyleBroekhuizen, Michelle, A. H. Jan Danser, Irwin K. M. Reiss, and Daphne Merkus. 2021. "The Function of the Kynurenine Pathway in the Placenta: A Novel Pharmacotherapeutic Target?" International Journal of Environmental Research and Public Health 18, no. 21: 11545. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182111545