
Polydatin Beneficial Effects in Zebrafish Larvae Undergoing Multiple Stress Types

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Rearing
2.2. Experimental Design
- Control-CTRL- control group reared in E3 medium from T0 to T3
- CuSO4- larvae exposed to 10 µM CuSO4 from T1 to T2
- PD-T0- larvae exposed to 400 µM PD from T0 to T1
- PD-T2- larvae exposed to 400 µM PD from T2 to T3
- PD + CuSO4- larvae exposed to 400 µM PD from T0 to T1 and to 10 µM CuSO4·5H2O from T1 to T2
- CuSO4 + PD- larvae exposed to 10 µM CuSO4·5H2O from T1 to T2 and to 400 µM PD from T2 to T3
- CTRL- reared in E3 medium
- PD-T0- Larvae exposed to PD from T0 to T1 and then reared in E3 m3dium till T3
- PD-T2 Larvae exposed to PD from T2 to T3 and sampled
- TAIL CUT- caudal fin amputation at T1 and reared in E3
- PD + TAIL CUT- larvae were treated with 400 µM PD from T0 to T1 before caudal fin amputation at T1 and then reared in E3 medium
- TAIL CUT + PD- caudal fin amputation at T1, transferred in E3 medium till T2 and treated with PD (400 µM) till T3.
2.3. Myeloperoxidase (MPO) Staining
2.4. RNA Extraction and cDNA Synthesis
2.5. Real Time PCR
2.6. Catalase Activity
2.7. Statistical Analysis
3. Results
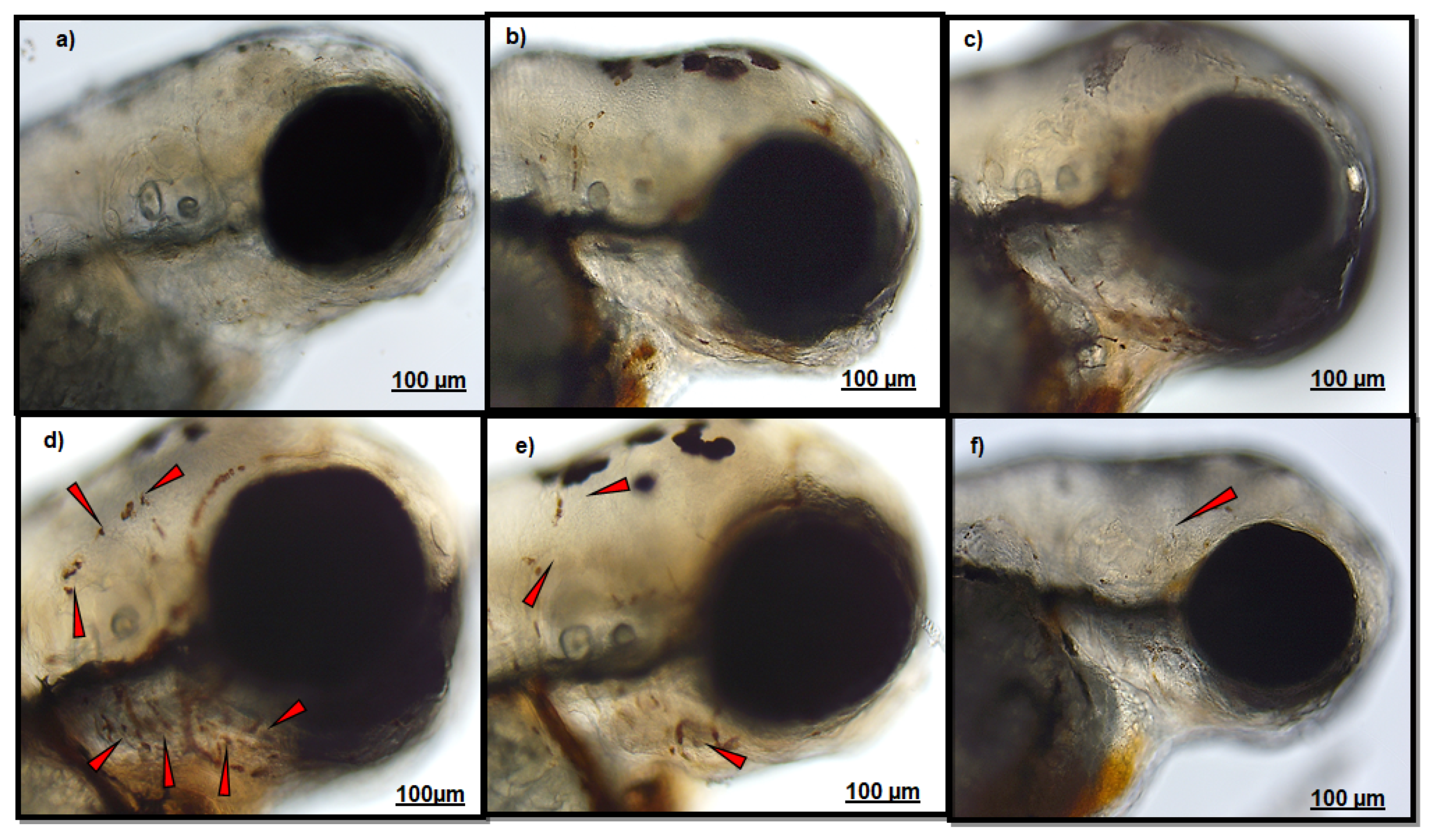
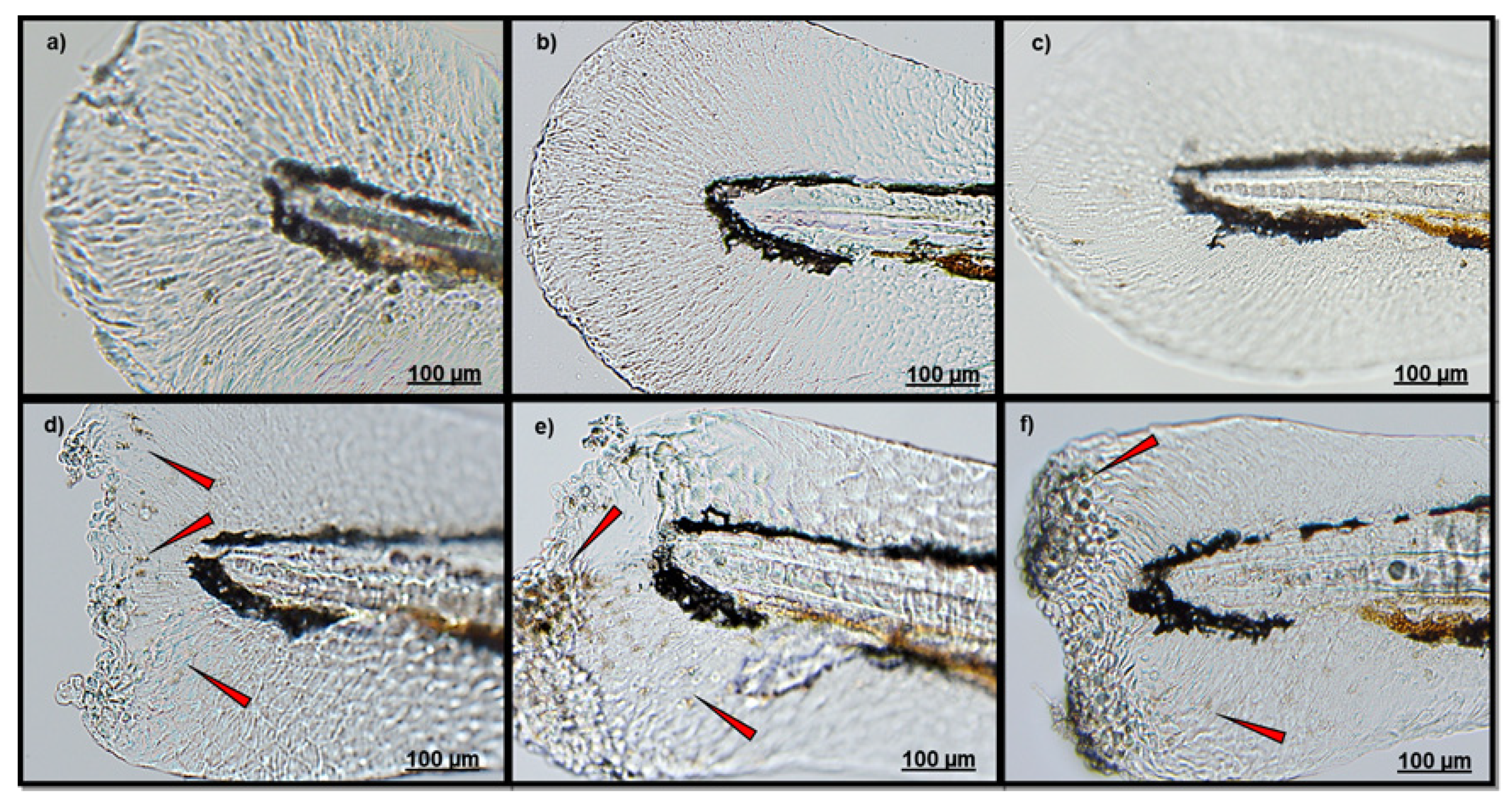
3.1. Effect of Polydatin on Neutrophils Migration
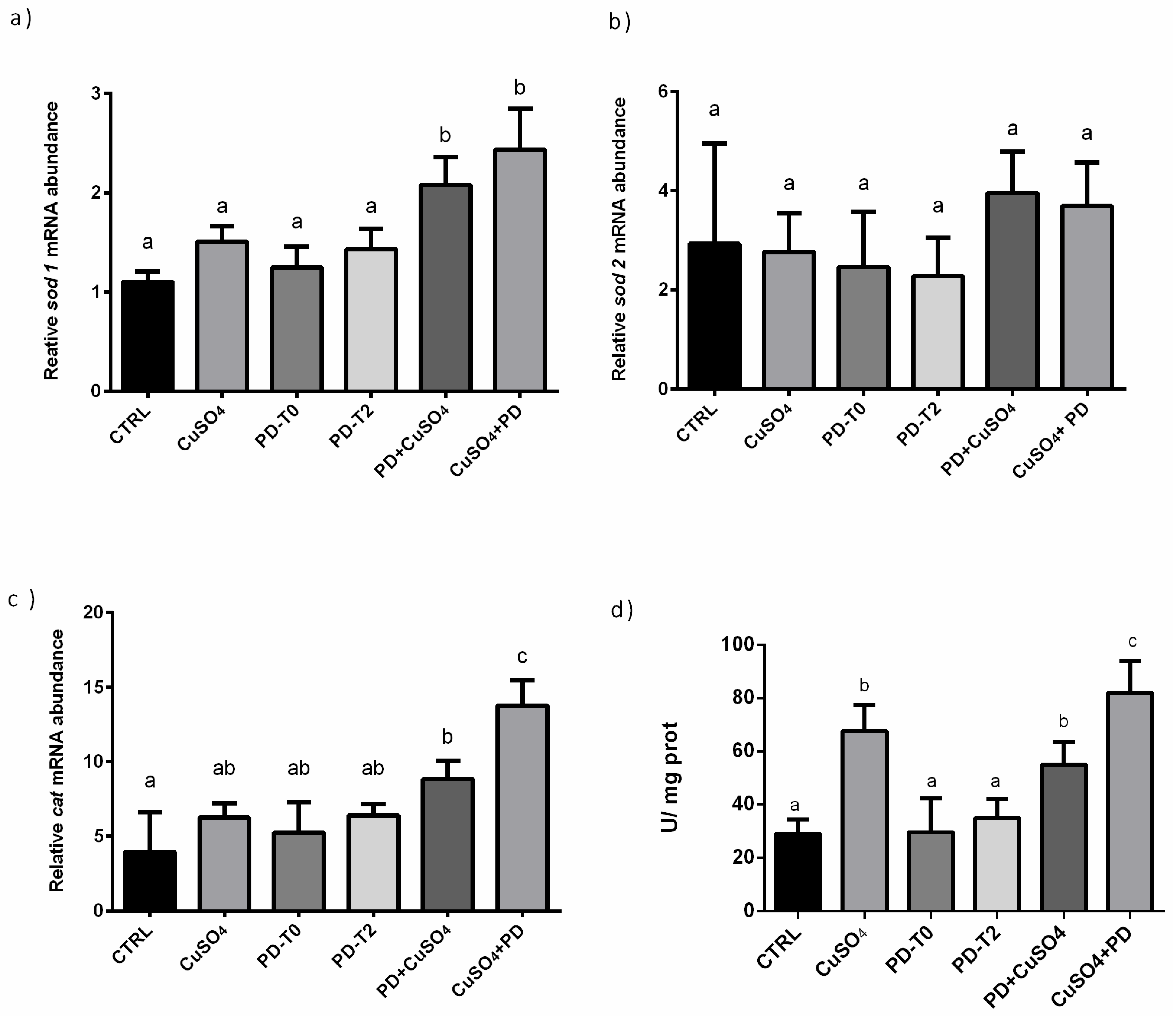
3.2. Effect of Polydatin on Oxidative Stress Pathway
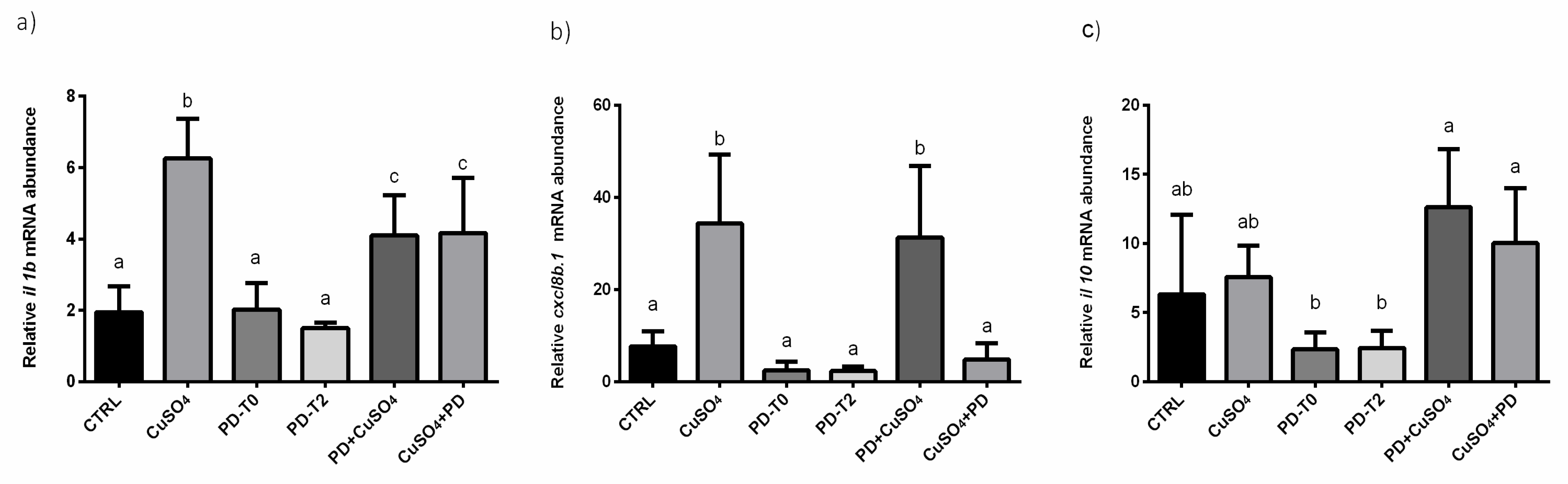
3.3. Effect of Polydatin on Inflammation Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Platella, C.; Raucci, U.; Rega, N.; D’Atri, S.; Levati, L.; Roviello, G.N.; Fuggetta, M.P.; Musumeci, D.; Montesarchio, D. Shedding light on the interaction of polydatin and resveratrol with G-quadruplex and duplex DNA: A biophysical, computational and biological approach. Int. J. Biol. Macromol. 2020, 151, 1163–1172. [Google Scholar] [CrossRef]
- Gu, L.; Liu, J.; Xu, D.; Lu, Y. Polydatin prevents LPS-induced acute kidney injury through inhibiting inflammatory and oxidative responses. Microb. Pathog. 2019, 137, 103688. [Google Scholar] [CrossRef]
- Liao, P.; He, Y.; Yang, F.; Luo, G.; Zhuang, J.; Zhai, Z.; Zhuang, L.; Lin, Z.; Zheng, J.; Sun, E. Polydatin effectively attenuates disease activity in lupus-prone mouse models by blocking ROS-mediated NET formation. Arthritis Res. Ther. 2018, 20, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ince, S.; Arslan Acaroz, D.; Neuwirth, O.; Demirel, H.H.; Denk, B.; Kucukkurt, I.; Turkmen, R. Protective effect of polydatin, a natural precursor of resveratrol, against cisplatin-induced toxicity in rats. Food Chem. Toxicol. 2014, 72, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Lv, R.; Du, L.; Zhang, L.; Zhang, Z. Polydatin attenuates spinal cord injury in rats by inhibiting oxidative stress and microglia apoptosis via Nrf2/HO-1 pathway. Life Sci. 2019, 217, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Feng, Q.; Shao, M.; Yuan, F.; Liu, F. Polydatin attenuates cadmium-induced oxidative stress via stimulating SOD activity and regulating mitochondrial function in Musca domestica larvae. Chemosphere 2020, 248, 126009. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Zhou, C.; Huang, P.; Dong, Z.; Mo, C.; Xie, L.; Lin, H.; Zhou, Z.; Deng, G.; Liu, Y.; et al. Polydatin alleviated alcoholic liver injury in zebrafish larvae through ameliorating lipid metabolism and oxidative stress. J. Pharmacol. Sci. 2018, 138, 46–53. [Google Scholar] [CrossRef]
- Gurău, F.; Baldoni, S.; Prattichizzo, F.; Espinosa, E.; Amenta, F.; Procopio, A.D.; Albertini, M.C.; Bonafè, M.; Olivieri, F. Anti-senescence compounds: A potential nutraceutical approach to healthy aging. Ageing Res. Rev. 2018, 46, 14–31. [Google Scholar] [CrossRef]
- Matacchione, G.; Gurău, F.; Baldoni, S.; Prattichizzo, F.; Silvestrini, A.; Giuliani, A.; Pugnaloni, A.; Espinosa, E.; Amenta, F.; Bonafè, M.; et al. Pleiotropic effects of polyphenols on glucose and lipid metabolism: Focus on clinical trials. Ageing Res. Rev. 2020, 61, 101074. [Google Scholar] [CrossRef]
- Cadena, P.G.; Sales Cadena, M.R.; Sarmah, S.; Marrs, J.A. Protective effects of quercetin, polydatin, and folic acid and their mixtures in a zebrafish (Danio rerio) fetal alcohol spectrum disorder model. Neurotoxicol. Teratol. 2020, 82, 106928. [Google Scholar] [CrossRef]
- Lam, S.H.; Chua, H.L.; Gong, Z.; Lam, T.J.; Sin, Y.M. Development and maturation of the immune system in zebrafish, Danio rerio: A gene expression profiling, in situ hybridization and immunological study. Dev. Comp. Immunol. 2004, 28, 9–28. [Google Scholar] [CrossRef]
- Murdoch, C.C.; Rawls, J.F. Commensal Microbiota Regulate Vertebrate Innate Immunity-Insights From the Zebrafish. Front. Immunol. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, M.A.; Sousa, L.P.; Pinho, V.; Perretti, M.; Teixeira, M.M. Resolution of inflammation: What controls its onset? Front. Immunol. 2016, 7, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mocchegiani, E.; Costarelli, L.; Giacconi, R.; Malavolta, M.; Basso, A.; Piacenza, F.; Ostan, R.; Cevenini, E.; Gonos, E.S.; Monti, D. Micronutrient-gene interactions related to inflammatory/immune response and antioxidant activity in ageing and inflammation. A systematic review. Mech. Ageing Dev. 2014, 136–137, 29–49. [Google Scholar] [CrossRef]
- Malavolta, M.; Piacenza, F.; Basso, A.; Giacconi, R.; Costarelli, L.; Mocchegiani, E. Serum copper to zinc ratio: Relationship with aging and health status. Mech. Ageing Dev. 2015, 151, 93–100. [Google Scholar] [CrossRef]
- Sozzani, S.; Bosisio, D.; Mantovani, A.; Ghezzi, P. Linking stress, oxidation and the chemokine system. Eur. J. Immunol. 2005, 35, 3095–3098. [Google Scholar] [CrossRef]
- Novoa, B.; Figueras, A. Zebrafish: Model for the study of inflammation and the innate immune response to infectious diseases. Curr. Top. Innate Immun. II Adv. Exp. Med. Biol. 2012, 946, 253–275. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Yan, B.; Shi, Y.Q.; Zhang, W.Q.; Wen, Z.L. Live imaging reveals differing roles of macrophages and neutrophils during zebrafish tail fin regeneration. J. Biol. Chem. 2012, 287, 25353–25360. [Google Scholar] [CrossRef] [Green Version]
- Pohanka, M. Copper and copper nanoparticles toxicity and their impact on basic functions in the body. Bratisl. Med. J. 2019, 1120, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Hordyjewska, A.; Popiołek, Ł.; Kocot, J. The many “faces” of copper in medicine and treatment. BioMetals 2014, 27, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Pei, R.; Zhang, Z.; Liao, J.; Yu, W.; Qiao, N.; Han, Q.; Li, Y.; Hu, L.; Guo, J.; et al. Copper induces oxidative stress and apoptosis through mitochondria-mediated pathway in chicken hepatocytes. Toxicol. In Vitro 2019, 54, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.; Chaturvedi, M.; Aggarwal, B. Oxidative stress, inflammation, and cancer. Free Radic. Biol. Med. 2011, 49, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.; Shekhar, H. Free Radicals in Human Health and Disease; Rani, V., Singh Yadav, U.C., Eds.; Springer: New Delhi, India, 2015; ISBN 9788132220350. [Google Scholar]
- Mocchegiani, E.; Costarelli, L.; Giacconi, R.; Piacenza, F.; Basso, A.; Malavolta, M. Micronutrient (Zn, Cu, Fe)-gene interactions in ageing and inflammatory age-related diseases: Implications for treatments. Ageing Res. Rev. 2012, 11, 297–319. [Google Scholar] [CrossRef] [PubMed]
- Wazir, S.M.; Ghobrial, I. Copper deficiency, a new triad: Anemia, leucopenia, and myeloneuropathy. J. Community Hosp. Intern. Med. Perspect. 2017, 7, 265–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Ji, X.; Cui, N.; Cao, S.; Liu, C.; Liu, J. Association between serum copper status and working memory in schoolchildren. Nutrients 2015, 7, 7185–7196. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, X.; Long, X.; Liu, Z.; Yan, S. Copper nanoparticles and copper sulphate induced cytotoxicity in hepatocyte primary cultures of Epinephelus coioides. PLoS ONE 2016, 11, e0149484. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Long, X.; Liu, Z.; Cheng, Y.; Yan, S. Effect of copper nanoparticles and copper sulphate on oxidation stress, cell apoptosis and immune responses in the intestines ofjuvenile Epinephelus coioides. Fish Shellfish Immunol. 2015, 44, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xu, L.; Wu, J.J.; Wang, W.M.; Mei, J.; Ma, X.F.; Liu, J.X. Transcriptional responses and mechanisms of copper-induced dysfunctional locomotor behavior in zebrafish embryos. Toxicol. Sci. 2015, 148, 299–310. [Google Scholar] [CrossRef] [Green Version]
- d’Alençon, C.A.; Peña, O.A.; Wittmann, C.; Gallardo, V.E.; Jones, R.A.; Loosli, F.; Liebel, U.; Grabher, C.; Allende, M.L. A high-throughput chemically induced inflammation assay in zebrafish. BMC Biol. 2010, 8, 151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Liu, X.; Li, Y.; Wang, M.; Chen, L.; Hu, B. Suppression of inflammation delays hair cell regeneration and functional recovery following lateral line damage in zebrafish larvae. Biomolecules 2020, 10, 1451. [Google Scholar] [CrossRef]
- Qin, L.; Liu, F.; Liu, H.; Wei, Z.; Sun, P.; Wang, Z. Evaluation of HODE-15, FDE-15, CDE-15, and BDE-15 toxicity on adult and embryonic zebrafish (Danio rerio). Environ. Sci. Pollut. Res. 2014, 21, 14047–14057. [Google Scholar] [CrossRef] [PubMed]
- Pompermaier, A.; Varela, A.C.C.; Fortuna, M.; Mendonça-Soares, S.; Koakoski, G.; Aguirre, R.; Oliveira, T.A.; Sordi, E.; Moterle, D.F.; Pohl, A.R.; et al. Water and suspended sediment runoff from vineyard watersheds affecting the behavior and physiology of zebrafish. Sci. Total Environ. 2021, 757, 143794. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, O.; Santobuono, M.; Forner-Piquer, I.; Randazzo, B.; Mylonas, C.C.; Ancillai, D.; Giorgini, E.; Maradonna, F. Dietary diisononylphthalate contamination induces hepatic stress: A multidisciplinary investigation in gilthead seabream (Sparus aurata) liver. Arch. Toxicol. 2019, 93, 2361–2373. [Google Scholar] [CrossRef] [PubMed]
- Santangeli, S.; Maradonna, F.; Zanardini, M.; Notarstefano, V.; Gioacchini, G.; Forner-Piquer, I.; Habibi, H.; Carnevali, O. Effects of diisononyl phthalate on Danio rerio reproduction. Environ. Pollut. 2017, 231, 1051–1062. [Google Scholar] [CrossRef]
- Pardal, D.; Caro, M.; Tueros, I.; Barranco, A.; Navarro, V. Resveratrol and piceid metabolites and their fat-reduction effects in zebrafish larvae. Zebrafish 2014, 11, 32–40. [Google Scholar] [CrossRef]
- Hernandez, P.P.; Undurraga, C.; Gallardo, V.E.; Mackenzie, N.; Allende, M.L.; Reyes, A.E. Sublethal concentrations of waterborne copper induce cellular stress and cell death in zebrafish embryos and larvae. Biol. Res. 2011, 44, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Kessels, M.Y.; Huitema, L.A.F.; Boeren, S.; Kranenbarg, S.; Schulte-Merker, S.; Van Leeuwen, J.L.; De Vries, S.C. Proteomics analysis of the Zebrafish Skeletal extracellular matrix. PLoS ONE 2014, 9, e90568. [Google Scholar] [CrossRef] [Green Version]
- Bates, J.M.; Akerlund, J.; Mittge, E.; Guillemin, K. Intestinal Alkaline Phosphatase Detoxifies Lipopolysaccharide and Prevents Inflammation in Zebrafish in Response to the Gut Microbiota. Cell Host. Microbe 2007, 2, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.J.; Chen, Y.H. Developmental nephrotoxicity of aristolochic acid in a zebrafish model. Toxicol. Appl. Pharmacol. 2012, 261, 59–65. [Google Scholar] [CrossRef]
- Piccinetti, C.C.; Montis, C.; Bonini, M.; Laurà, R.; Guerrera, M.C.; Radaelli, G.; Vianello, F.; Santinelli, V.; Maradonna, F.; Nozzi, V.; et al. Transfer of Silica-Coated Magnetic (Fe3O4) Nanoparticles Through Food: A Molecular and Morphological Study in Zebrafish. Zebrafish 2014, 11, 567–579. [Google Scholar] [CrossRef]
- Velki, M.; Meyer-Alert, H.; Seiler, T.B.; Hollert, H. Enzymatic activity and gene expression changes in zebrafish embryos and larvae exposed to pesticides diazinon and diuron. Aquat. Toxicol. 2017, 193, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Lv, T.; Shen, L.; Yang, L.; Diao, W.; Yang, Z.; Zhang, Y.; Yu, S.; Li, Y. Polydatin ameliorates dextran sulfate sodium-induced colitis by decreasing oxidative stress and apoptosis partially via Sonic hedgehog signaling pathway. Int. Immunopharmacol. 2018, 64, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Leibovich, S.J. Inflammatory cells during wound repair: The good, the bad and the ugly. Trends Cell Biol. 2005, 15, 599–607. [Google Scholar] [CrossRef] [PubMed]
- von Gersdorff Jørgensen, L.; Korbut, R.; Jeberg, S.; Kania, P.W.; Buchmann, K. Association between adaptive immunity and neutrophil dynamics in zebrafish (Danio rerio) infected by a parasitic ciliate. PLoS ONE 2018, 13, e0203297. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Van Doan, H.; Ashouri, G.; Gioacchini, G.; Maradonna, F.; Carnevali, O. Oxidative Stress and Antioxidant Defense in Fish: The Implications of Probiotic, Prebiotic, and Synbiotics. Rev. Fish. Sci. Aquac. 2020, 1–20. [Google Scholar] [CrossRef]
- Mao, L.; Jia, W.; Zhang, L.; Zhang, Y.; Zhu, L.; Sial, M.U.; Jiang, H. Embryonic development and oxidative stress effects in the larvae and adult fish livers of zebrafish (Danio rerio) exposed to the strobilurin fungicides, kresoxim-methyl and pyraclostrobin. Sci. Total Environ. 2020, 729, 139031. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Niethammer, P.; Grabher, C.; Look, A.T.; Mitchison, T.J. A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in zebrafish. Nature 2009, 459, 996–999. [Google Scholar] [CrossRef]
- Lou, Z.; Li, X.; Zhao, X.; Du, K.; Li, X.; Wang, B. Resveratrol attenuates hydrogen peroxide-induced apoptosis, reactive oxygen species generation, and PSGL-1 and VWF activation in human umbilical vein endothelial cells, potentially via MAPK signalling pathways. Mol. Med. Rep. 2018, 17, 2479–2487. [Google Scholar] [CrossRef]
- Bosutti, A.; Degens, H. The impact of resveratrol and hydrogen peroxide on muscle cell plasticity shows a dose-dependent interaction. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Sun, Y.; Li, Z. Resveratrol protects Leydig cells from nicotine-induced oxidative damage through enhanced autophagy. Clin. Exp. Pharmacol. Physiol. 2018, 45, 573–580. [Google Scholar] [CrossRef]
- Fukui, M.; Ting Zhu, B. Mitochondrial Superoxide Dismutase SOD2, but not Cytosolic SOD1, Plays a Critical Role in Protection against Glutamate-Induced Oxidative Stress and Cell Death in HT22 Neuronal Cells. Free Radic. Biol. Med. 2010, 48, 821–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torraca, V.; Otto, N.A.; Tavakoli-Tameh, A.; Meijer, A.H. The inflammatory chemokine Cxcl18b exerts neutrophil-specific chemotaxis via the promiscuous chemokine receptor Cxcr2 in zebrafish. Dev. Comp. Immunol. 2017, 67, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roebuck, K.A. Oxidant stress regulation of IL-8 and ICAM-1 gene expression: Differential activation and binding of the transcription factors AP-1 and NF-kappaB (Review). Int. J. Mol. Med. 1999, 4, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Jeong, M.; Park, J.S.; Kim, M.H.; Lee, D.B.; Shin, B.A.; Mukaida, N.; Ellis, L.M.; Kim, H.R.; Ahn, B.W.; et al. Interleukin-1β stimulates IL-8 expression through MAP kinase and ROS signaling in human gastric carcinoma cells. Oncogene 2004, 23, 6603–6611. [Google Scholar] [CrossRef] [Green Version]
- Ravagnan, G.; De Filippis, A.; Cartenì, M.; De Maria, S.; Cozza, V.; Petrazzuolo, M.; Tufano, M.A.; Donnarumma, G. Polydatin, a natural precursor of resveratrol, induces β-defensin production and reduces inflammatory response. Inflammation 2013, 36, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Austin, B. Cytokine expression in leucocytes and gut cells of rainbow trout, Oncorhynchus mykiss Walbaum, induced by probiotics. Vet. Immunol. Immunopathol. 2006, 114, 297–304. [Google Scholar] [CrossRef]
- Raida, M.K.; Buchmann, K. Development of adaptive immunity in rainbow trout, Oncorhynchus mykiss (Walbaum) surviving an infection with Yersinia ruckeri. Fish Shellfish Immunol. 2008, 25, 533–541. [Google Scholar] [CrossRef]
- Pérez-Sánchez, T.; Balcázar, J.L.; Merrifield, D.L.; Carnevali, O.; Gioacchini, G.; de Blas, I.; Ruiz-Zarzuela, I. Expression of immune-related genes in rainbow trout (Oncorhynchus mykiss) induced by probiotic bacteria during Lactococcus garvieae infection. Fish Shellfish Immunol. 2011, 31, 196–201. [Google Scholar] [CrossRef]
- Gioacchini, G.; Giorgini, E.; Olivotto, I.; Maradonna, F.; Merrifield, D.L.; Carnevali, O. The Influence of Probiotics on Zebrafish Danio Rerio Innate Immunity and Hepatic Stress. Zebrafish 2014, 11, 98–106. [Google Scholar] [CrossRef]
- Zhao, X.J.; Yu, H.W.; Yang, Y.Z.; Wu, W.Y.; Chen, T.Y.; Jia, K.K.; Kang, L.L.; Jiao, R.Q.; Kong, L.D. Polydatin prevents fructose-induced liver inflammation and lipid deposition through increasing miR-200a to regulate Keap1/Nrf2 pathway. Redox Biol. 2018, 18, 124–137. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | ||||
|---|---|---|---|---|
| PD-CuSO4 | CuSO4-PD | |||
| Endpoints | MPO Test | Head Localization | ↓ | ↓↓ |
| Lateral Line | - | ↓ | ||
| RT-PCR | sod1 | ↑ | ↑ | |
| sod2 | - | - | ||
| cat | - | ↑ | ||
| il 1b | ↓ | ↓ | ||
| cxcl8b.1 | ↑ | - | ||
| il 10 | - | - | ||
| Enzyme Activity | CAT | ↑ | ↑ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pessina, A.; Di Vincenzo, M.; Maradonna, F.; Marchegiani, F.; Olivieri, F.; Randazzo, B.; Gioacchini, G.; Carnevali, O. Polydatin Beneficial Effects in Zebrafish Larvae Undergoing Multiple Stress Types. Int. J. Environ. Res. Public Health 2021, 18, 1116. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18031116
Pessina A, Di Vincenzo M, Maradonna F, Marchegiani F, Olivieri F, Randazzo B, Gioacchini G, Carnevali O. Polydatin Beneficial Effects in Zebrafish Larvae Undergoing Multiple Stress Types. International Journal of Environmental Research and Public Health. 2021; 18(3):1116. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18031116
Chicago/Turabian StylePessina, Andrea, Mariangela Di Vincenzo, Francesca Maradonna, Francesca Marchegiani, Fabiola Olivieri, Basilio Randazzo, Giorgia Gioacchini, and Oliana Carnevali. 2021. "Polydatin Beneficial Effects in Zebrafish Larvae Undergoing Multiple Stress Types" International Journal of Environmental Research and Public Health 18, no. 3: 1116. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18031116