
Structure of Bacterial Community with Resistance to Antibiotics in Aquatic Environments. A Systematic Review


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy and Selection Criteria
2.2. Eligibility Criteria
2.3. Search for Articles
2.4. Data Extraction
3. Results and Discussion
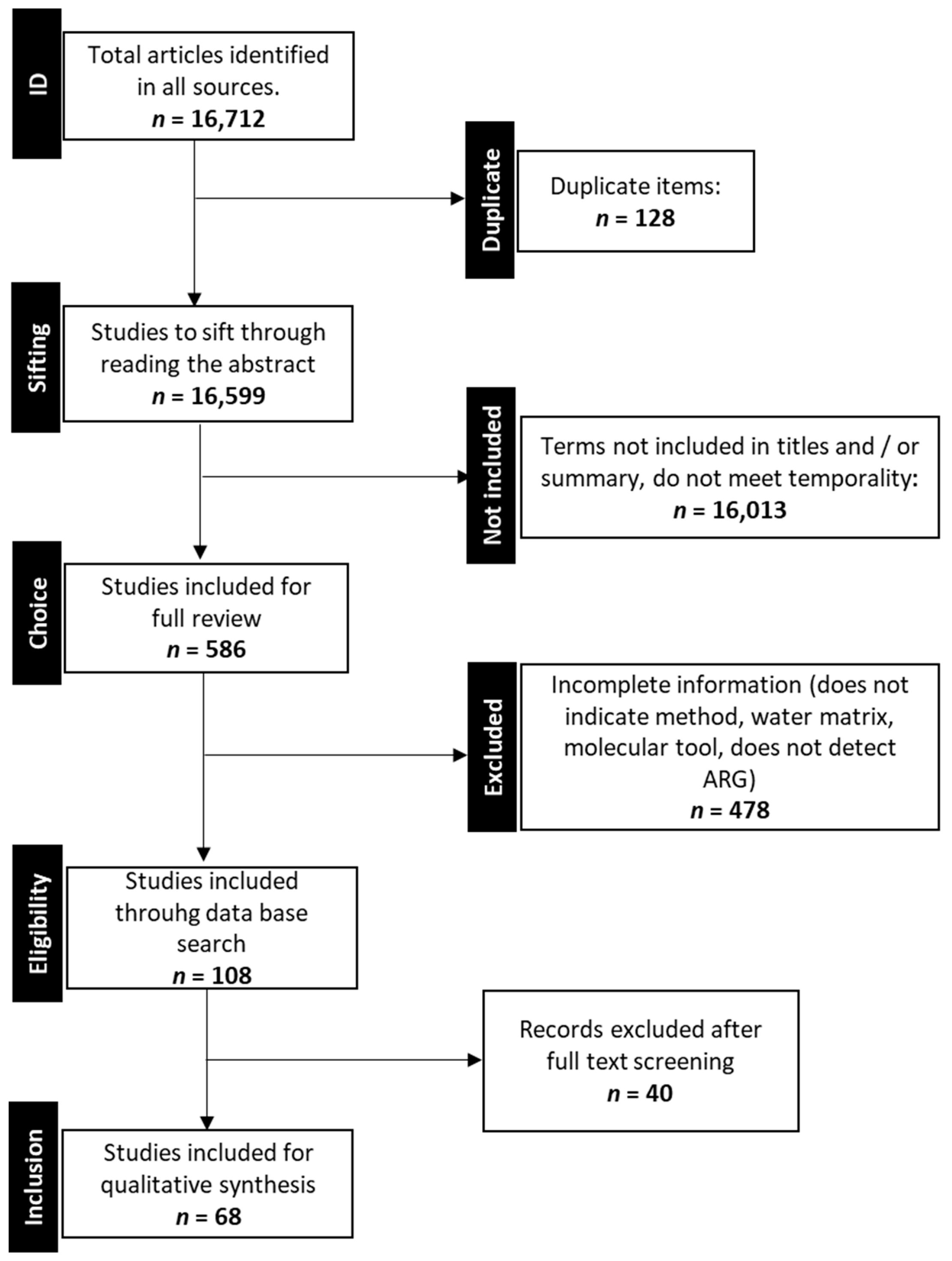
3.1. Search Results
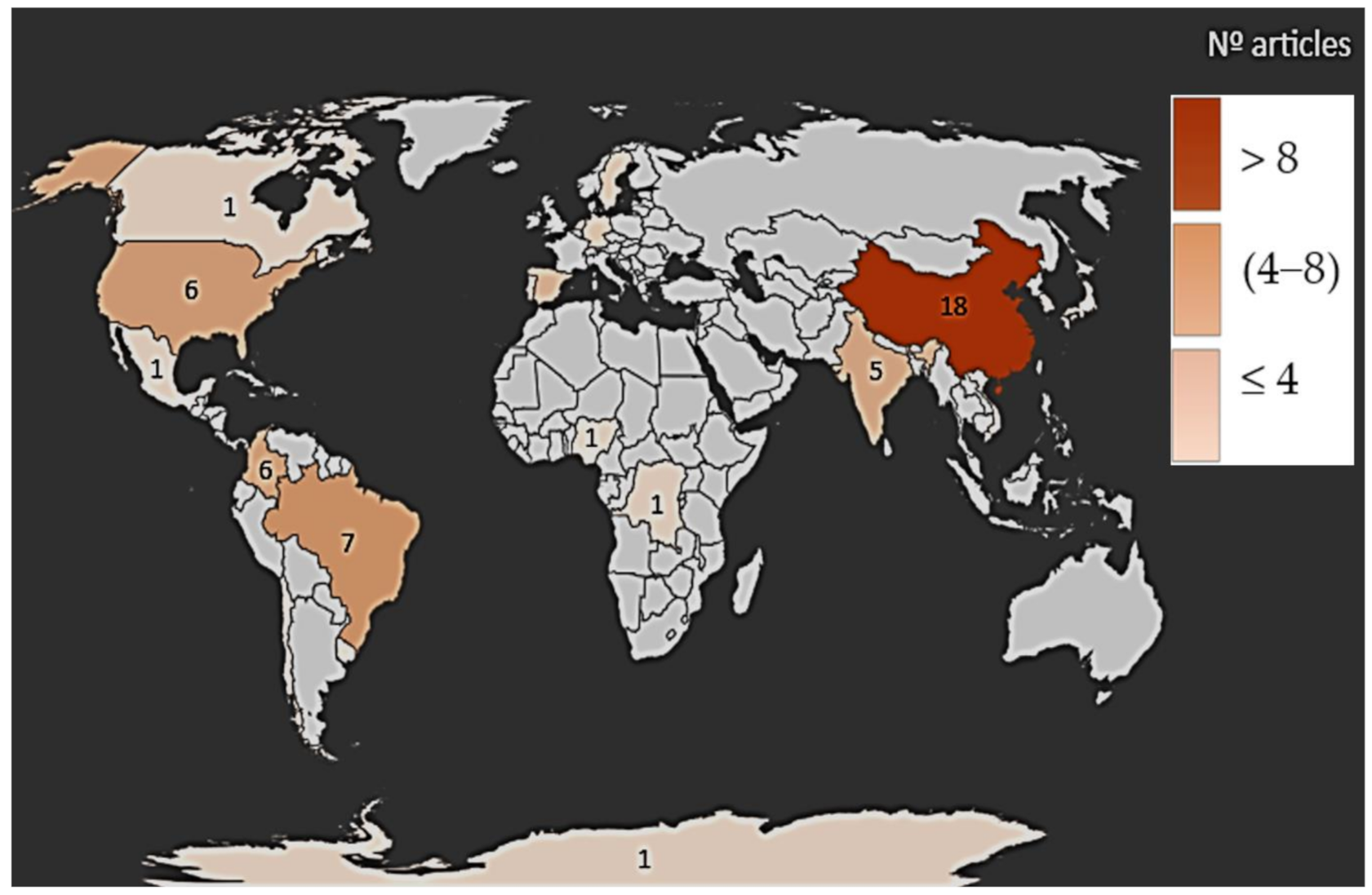
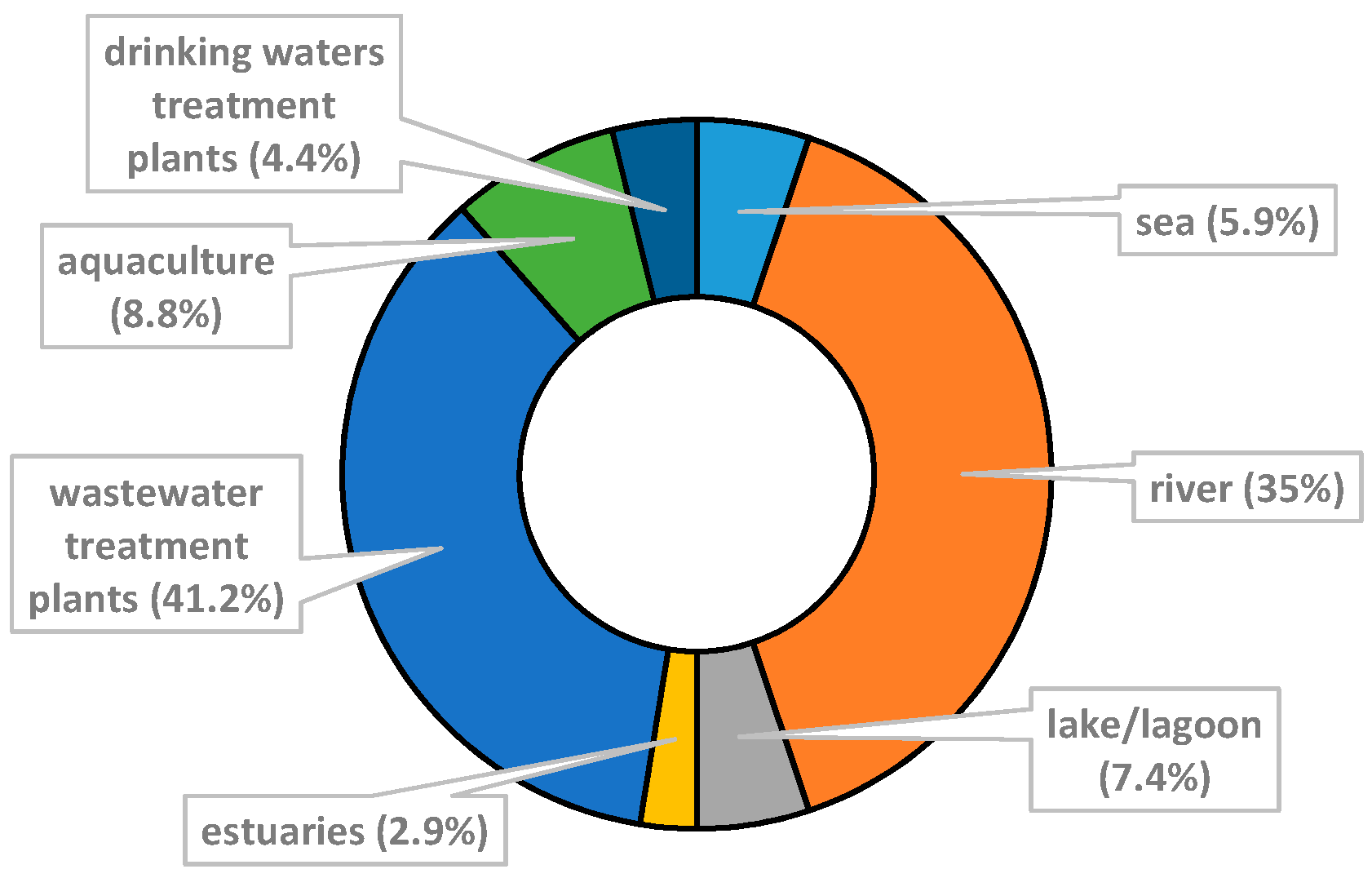
3.2. Characteristics of Included Studies
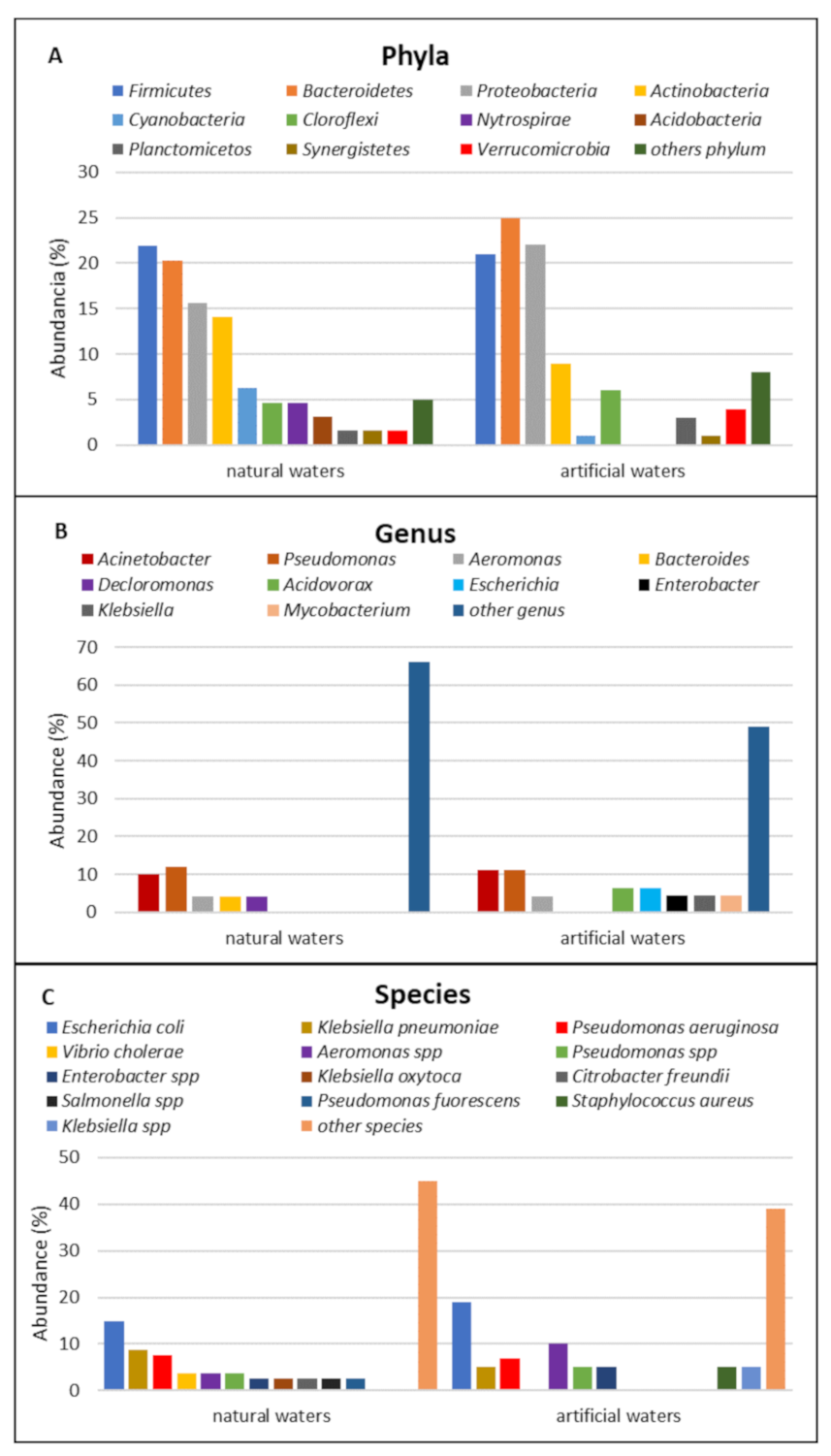
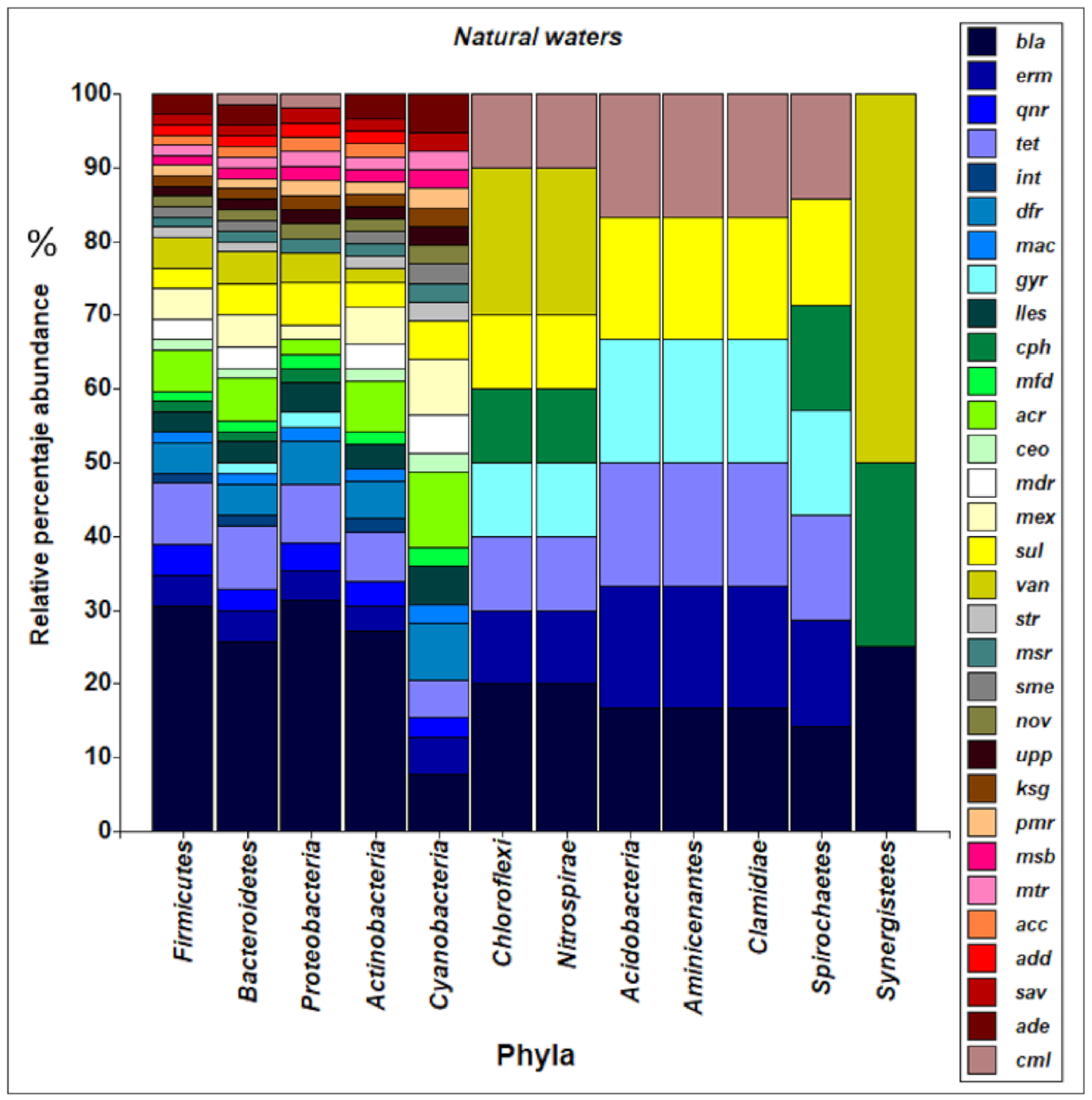
3.3. Structure of the Bacterial Community of the Aquatic Environment
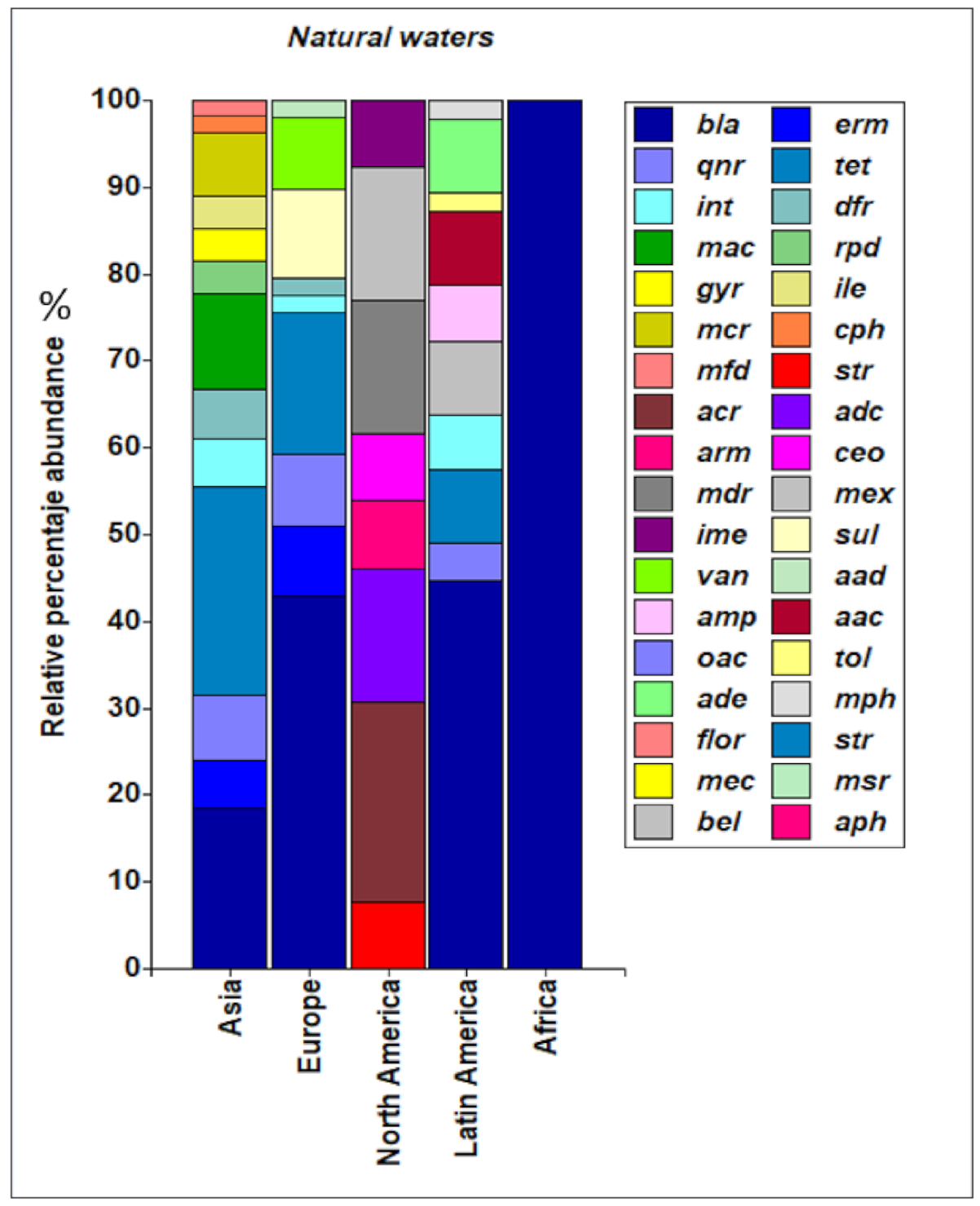
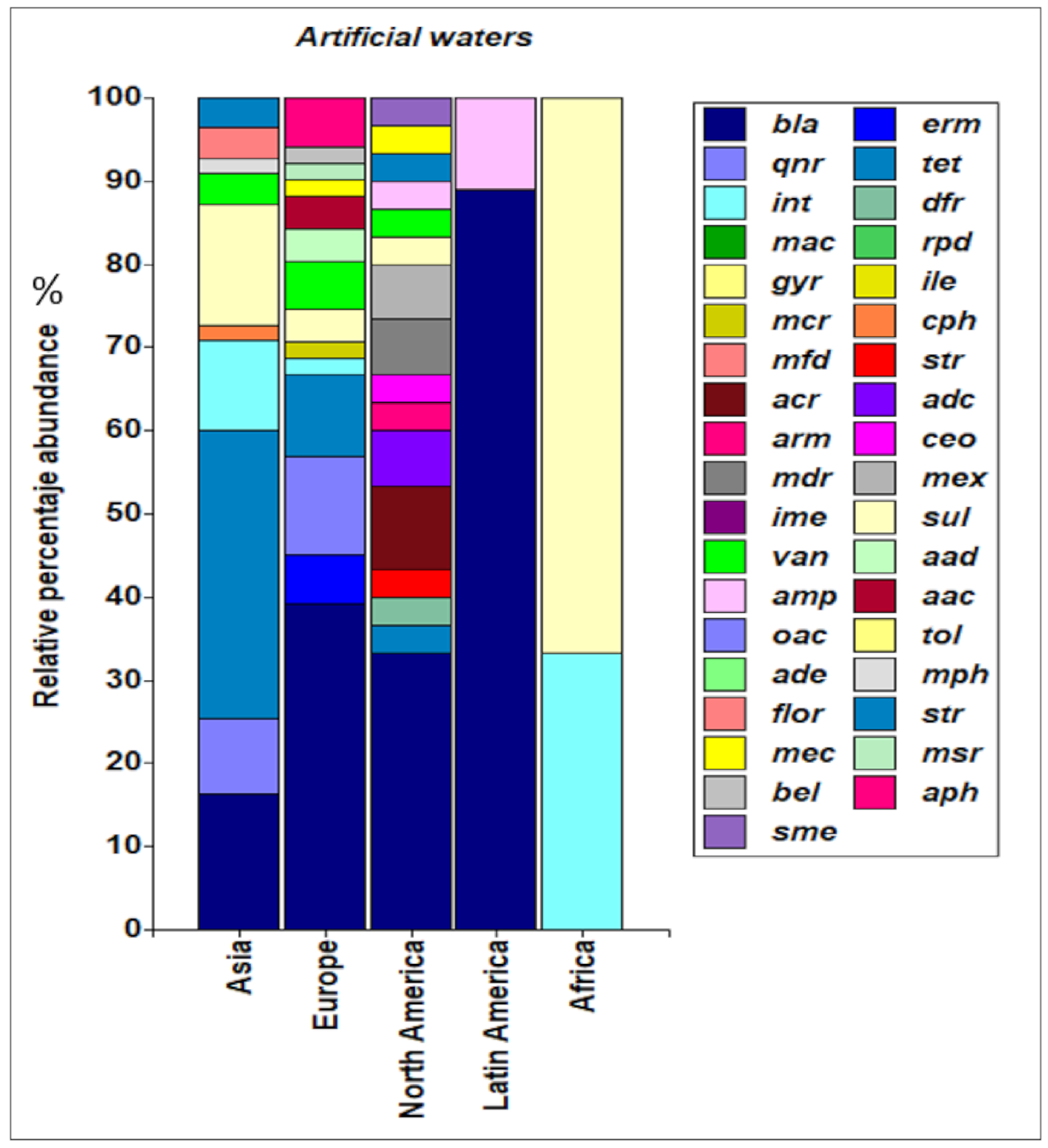
3.4. Abundance of ARGs in Aquatic Environments
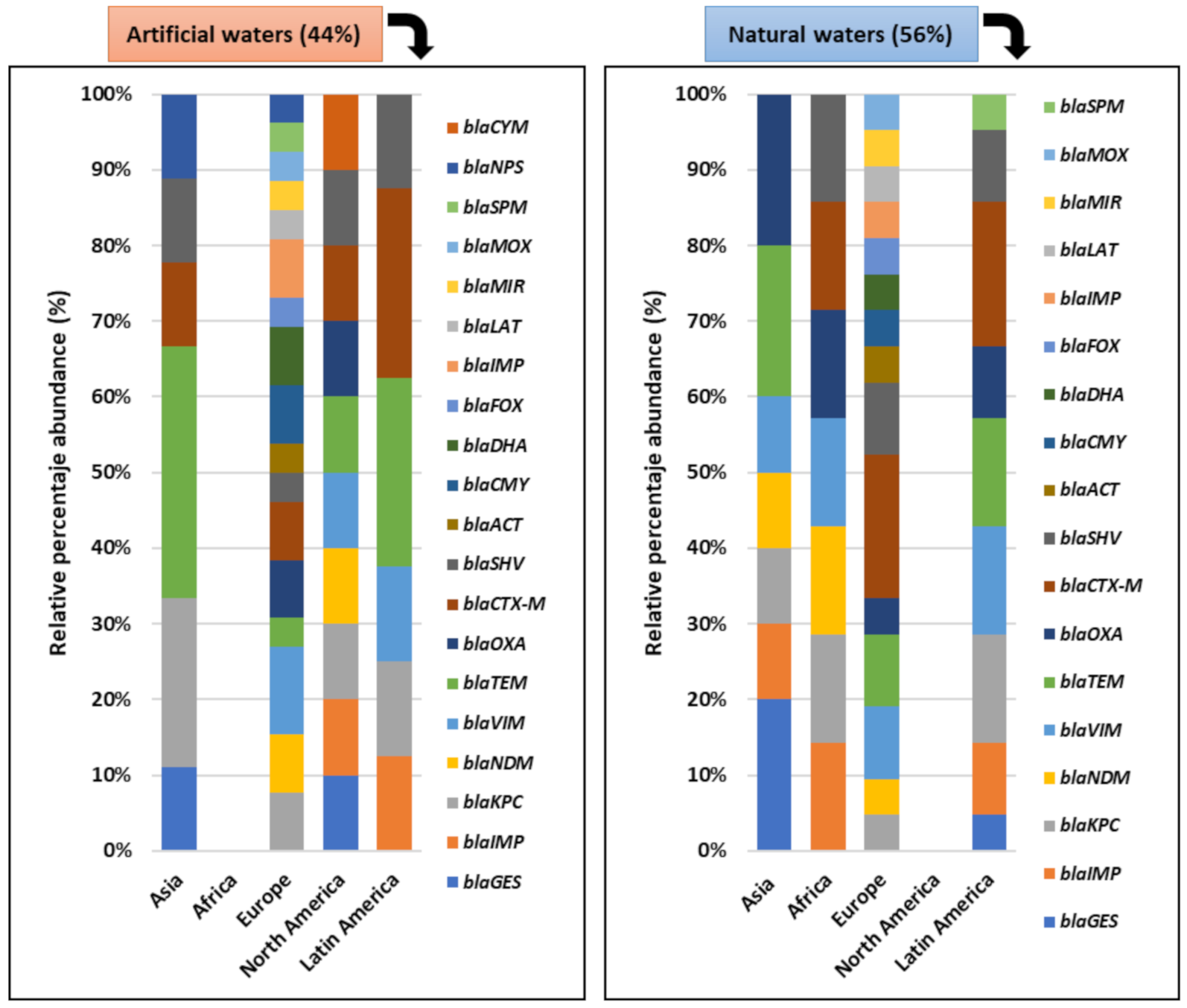
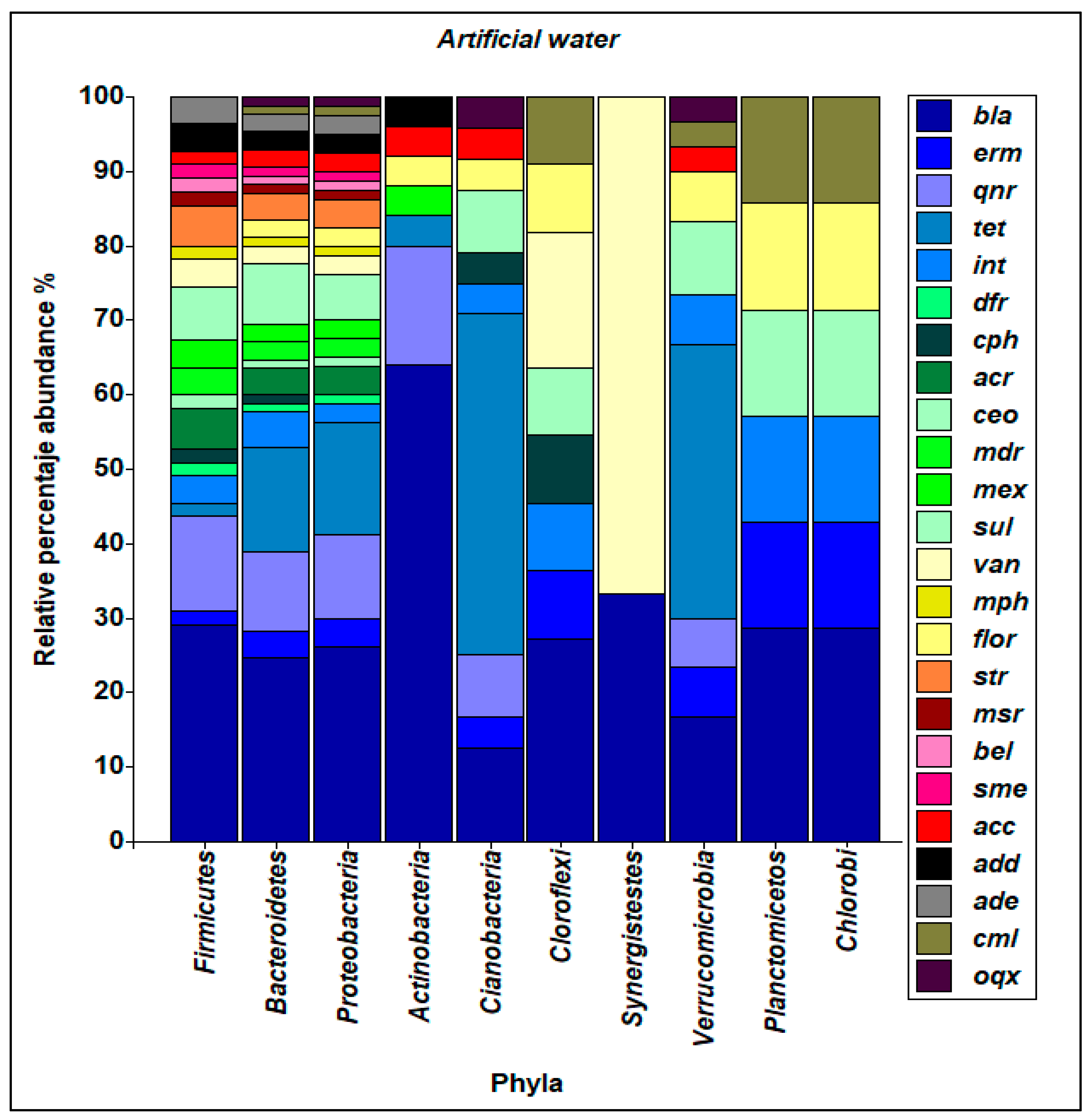
3.5. Composition of the Antibiotic-Resistant Bacterial Community and ARGs
3.6. Mobile Genetic Element
3.7. Effect of Exposure to WWTP Effluent in Aquatic Ecosystems
3.8. Limitations of the Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Decker, B.K.; Palmore, T.N. The role of water in healthcare-associated infections. Curr. Opin. Infect. Dis. 2013, 26, 345–351. [Google Scholar] [CrossRef]
- Manzetti, S.; Ghisi, R. The environmental release and fate of antibiotics. Mar. Pollut. Bull. 2014, 79, 7–15. [Google Scholar] [CrossRef]
- Ding, C.; He, J. Effect of antibiotics in the environment on microbial populations. Appl. Microbiol. Biotechnol. 2010, 87, 925–941. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Pruden, A.; Virta, M.; Zhang, T. Editorial: Antibiotic Resistance in Aquatic Systems. Front. Microbiol. 2017, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Elupo, A.; Ecoyne, S.; Berendonk, T.U. Origin and Evolution of Antibiotic Resistance: The Common Mechanisms of Emergence and Spread in Water Bodies. Front. Microbiol. 2012, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Yang, Y.; Liang, X.; Yu, K.; Zhang, T.; Li, X. Metagenomic Pro fi les of Antibiotic Resistance Genes (ARGs) between Human Impacted Estuary and Deep Ocean Sediments. Environ. Sci. Technol. 2013, 47, 12753–12760. [Google Scholar] [CrossRef] [PubMed]
- Czekalski, N.; Díez, E.G.; Bürgmann, H. Wastewater as a point source of antibiotic-resistance genes in the sediment of a freshwater lake. ISME J. 2014, 8, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Sivalingam, P.; Poté, J.; Prabakar, K. Environmental Prevalence of Carbapenem Resistance Enterobacteriaceae (CRE) in a Tropical Ecosystem in India: Human Health Perspectives and Future Directives. Pathogens 2019, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Knapp, C.W.; Beattie, T.K. Antibiotic Resistant Bacteria Found in Municipal Drinking Water. Environ. Process. 2016, 3, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. J. Clin. Epidemiol. 2009, 62, e1–e34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, F.A.; Söderquist, B.; Jass, J. Prevalence and Diversity of Antibiotic Resistance Genes in Swedish Aquatic Environments Impacted by Household and Hospital Wastewater. Front. Microbiol. 2019, 10, 688. [Google Scholar] [CrossRef] [Green Version]
- Chu, B.T.T.; Petrovich, M.L.; Chaudhary, A.; Wright, D.; Murphy, B.; Wells, G.; Poretsky, R. Metagenomics Reveals the Impact of Wastewater Treatment Plants on the Dispersal of Microorganisms and Genes in Aquatic Sediments. Appl. Environ. Microbiol. 2017, 84, e02168-17. [Google Scholar] [CrossRef] [Green Version]
- Girijan, S.K.; Paul, R.; Rejish Kumar, V.J.; Pillai, D. Investigating the impact of hospital antibiotic usage on aquatic environment and aquaculture systems: A molecular study of quinolone resistance in Escherichia coli. Sci. Total Environ. 2020, 748, 141538. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Yu, T.; Zhang, Y.; Yang, M.; Li, Z.; Liu, M.; Qi, R. Antibiotic Resistance Characteristics of Environmental Bacteria from an Oxytetracycline Production Wastewater Treatment Plant and the Receiving River. Appl. Environ. Microbiol. 2010, 76, 3444–3451. [Google Scholar] [CrossRef] [Green Version]
- Vivas, M.C.; Caicedo, L.D.; Castillo, J.E. Occurrence ofβ-Lactamase-Producing Gram-Negative Bacterial Isolates in Water Sources in Cali City, Colombia. Int. J. Microbiol. 2019, 2019, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Zhao, W.; Xu, T.; Zheng, B.; Yin, D. Occurrence and distribution of antibiotic resistance genes in the water and sediments of Qingcaosha Reservoir, Shanghai, China. Environ. Sci. Eur. 2019, 31, 1–9. [Google Scholar] [CrossRef]
- Oliveira, M.; Nunes, M.; Crespo, M.T.B.; Silva, A.F. The environmental contribution to the dissemination of carbapenem and (fluoro)quinolone resistance genes by discharged and reused wastewater effluents: The role of cellular and extracellular DNA. Water Res. 2020, 182, 116011. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Jia, J.; Wu, L.; Xue, X.; Zhang, G.; Wang, Z. Analysis of Bacterial Community Characteristics, Abundance of Antibiotics and Antibiotic Resistance Genes Along a Pollution Gradient of Ba River in Xi’an, China. Front. Microbiol. 2018, 9, 3191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-Y.; An, X.-L.; Huang, F.-Y.; Su, J.-Q. Antibiotic resistome in a landfill leachate treatment plant and effluent-receiving river. Chemosphere 2020, 242, 125207. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bai, X.; Li, Y.; Jing, L.; Chen, R.; Teng, Y. Source identification of antibiotic resistance genes in a peri-urban river using novel crAssphage marker genes and metagenomic signatures. Water Res. 2019, 167, 115098. [Google Scholar] [CrossRef]
- Berglund, B.; Fick, J.; Lindgren, P.-E. Urban wastewater effluent increases antibiotic resistance gene concentrations in a receiving northern European river. Environ. Toxicol. Chem. 2015, 34, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Herrig, I.; Fleischmann, S.; Regnery, J.; Wesp, J.; Reifferscheid, G.; Manz, W. Prevalence and seasonal dynamics of blaCTX-M antibiotic resistance genes and fecal indicator organisms in the lower Lahn River, Germany. PLoS ONE 2020, 15, e0232289. [Google Scholar] [CrossRef] [PubMed]
- Collins-Fairclough, A.M.; Co, R.; Ellis, M.C.; Hug, L.A. Widespread Antibiotic, Biocide, and Metal Resistance in Microbial Communities Inhabiting a Municipal Waste Environment and Anthropogenically Impacted River. mSphere 2018, 3, e00346-18. [Google Scholar] [CrossRef] [Green Version]
- Subirats, J.; Triadó-Margarit, X.; Mandaric, L.; Acuña, V.; Balcázar, J.L.; Sabater, S.; Borrego, C.M. Wastewater pollution differently affects the antibiotic resistance gene pool and biofilm bacterial communities across streambed compartments. Mol. Ecol. 2017, 26, 5567–5581. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, V.D.; Meirelles-Pereira, F.; Cataldo, M.; Fonseca, B.D.O.; Nogueira, B.A.; Olivella, J.G.B.; Esteves, F.D.A.; Mattos-Guaraldi, A.L.; De Andrade, A.F.B.; Bello, A.R.; et al. Detection of multidrug-resistant Enterobacteriaceae isolated from river waters flowing to the Guanabara Bay and from clinical samples of hospitals in Rio de Janeiro, Brazil. Biomédica 2019, 39, 135–149. [Google Scholar] [CrossRef]
- Posada-Perlaza, C.E.; Ramírez-Rojas, A.; Porras, P.; Adu-Oppong, B.; Botero-Coy, A.-M.; Hernández, F.; Anzola, J.M.; Díaz, L.; Dantas, G.; Reyes, A.; et al. Bogotá River anthropogenic contamination alters microbial communities and promotes spread of antibiotic resistance genes. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Port, J.A.; Wallace, J.C.; Griffith, W.C.; Faustman, E.M. Metagenomic Profiling of Microbial Composition and Antibiotic Resistance Determinants in Puget Sound. PLoS ONE 2012, 7, e48000. [Google Scholar] [CrossRef] [Green Version]
- Kristiansson, E.; Fick, J.; Janzon, A.; Grabic, R.; Rutgersson, C.; Weijdegård, B.; Söderström, H.; Larsson, D.G.J. Pyrosequencing of Antibiotic-Contaminated River Sediments Reveals High Levels of Resistance and Gene Transfer Elements. PLoS ONE 2011, 6, e17038. [Google Scholar] [CrossRef]
- Marti, E.; Jofre, J.; Balcazar, J.L. Prevalence of Antibiotic Resistance Genes and Bacterial Community Composition in a River Influenced by a Wastewater Treatment Plant. PLoS ONE 2013, 8, e78906. [Google Scholar] [CrossRef] [PubMed]
- Sekizuka, T.; Inamine, Y.; Segawa, T.; Hashino, M.; Yatsu, K.; Kuroda, M. Potential KPC-2 carbapenemase reservoir of environmental Aeromonas hydrophila and Aeromonas caviae isolates from the effluent of an urban wastewater treatment plant in Japan. Environ. Microbiol. Rep. 2019, 11, 589–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xie, J.; Liu, M.; Tian, Z.; He, Z.; Van Nostrand, J.D.; Ren, L.; Zhou, J.; Yang, M. Microbial community functional structure in response to antibiotics in pharmaceutical wastewater treatment systems. Water Res. 2013, 47, 6298–6308. [Google Scholar] [CrossRef]
- Yuan, K.; Wang, X.; Chen, X.; Zhao, Z.; Fang, L.; Chen, B.; Jiang, J.; Luan, T.; Chen, B. Occurrence of antibiotic resistance genes in extracellular and intracellular DNA from sediments collected from two types of aquaculture farms. Chemosphere 2019, 234, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, B.; Jiang, X.-T.; Wang, Y.-L.; Xiao-Tao, J.; Li, A.-D.; Zhang, T. Catalogue of antibiotic resistome and host-tracking in drinking water deciphered by a large scale survey. Microbiome 2017, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Gardea, M.C.E.; Tamez-Guerra, P.; Gomez-Flores, R.; De La Serna, F.J.Z.-D.; La Vega, G.E.-D.; Nevárez-Moorillón, G.V.; Pérez-Recoder, M.C.; Sánchez-Ramírez, B.; González-Horta, M.D.C.; Infante-Ramírez, R. Multidrug-Resistant Bacteria Isolated from Surface Water in Bassaseachic Falls National Park, Mexico. Int. J. Environ. Res. Public Health 2016, 13, 597. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.M.; Kim, Y.B.; Choi, S.; Lee, Y.; Shin, S.G.; Unno, T. Prevalence of antibiotic resistance genes from effluent of coastal aquaculture, South Korea. Environ. Pollut. 2018, 233, 1049–1057. [Google Scholar] [CrossRef]
- Obayiuwana, A.; Ogunjobi, A.; Yang, M.; Ibekwe, M. Characterization of Bacterial Communities and Their Antibiotic Resistance Profiles in Wastewaters Obtained from Pharmaceutical Facilities in Lagos and Ogun States, Nigeria. Int. J. Environ. Res. Public Health 2018, 15, 1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendriksen, R.S.; Munk, P.; Njage, P.; Van Bunnik, B.; McNally, L.; Lukjancenko, O.; Röder, T.; Nieuwenhuijse, D.; Pedersen, S.K.; Kjeldgaard, J.; et al. Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 2019, 10, 1124. [Google Scholar] [CrossRef]
- Cacace, D.; Fatta-Kassinos, D.; Manaia, C.M.; Cytryn, E.; Kreuzinger, N.; Rizzo, L.; Karaolia, P.; Schwartz, T.; Alexander, J.; Merlin, C.; et al. Antibiotic resistance genes in treated wastewater and in the receiving water bodies: A pan-European survey of urban settings. Water Res. 2019, 162, 320–330. [Google Scholar] [CrossRef]
- Jara, D.; Bello-Toledo, H.; Domínguez, M.; Cigarroa, C.; Fernández, P.; Vergara, L.; Quezada-Aguiluz, M.; Opazo-Capurro, A.; Lima, C.A.; González-Rocha, G. Antibiotic resistance in bacterial isolates from freshwater samples in Fildes Peninsula, King George Island, Antarctica. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Buelow, E.; Bayjanov, J.R.; Majoor, E.; Willems, R.J.L.; Bonten, M.J.M.; Schmitt, H.; Van Schaik, W. Limited influence of hospital wastewater on the microbiome and resistome of wastewater in a community sewerage system. FEMS Microbiol. Ecol. 2018, 94, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lira, F.; Vaz-Moreira, I.; Tamames, J.; Manaia, C.M.; Martínez, J.L. Metagenomic analysis of an urban resistome before and after wastewater treatment. Sci. Rep. 2020, 10, 8174. [Google Scholar] [CrossRef]
- Quintela-Baluja, M.; Abouelnaga, M.; Romalde, J.; Su, J.-Q.; Yu, Y.; Gomez-Lopez, M.; Smets, B.; Zhu, Y.-G.; Graham, D.W. Spatial ecology of a wastewater network defines the antibiotic resistance genes in downstream receiving waters. Water Res. 2019, 162, 347–357. [Google Scholar] [CrossRef]
- Rodríguez, E.; Garzón, L.; Gómez, I.; Jiménez, J.N. Multidrug resistance and diversity of resistance profiles in carbapenem-resistant Gram-negative bacilli throughout a wastewater treatment plant in Colombia. J. Glob. Antimicrob. Resist. 2020, 22, 358–366. [Google Scholar] [CrossRef]
- Aristizábal-Hoyos, A.; Rodríguez, E.; Arias, L.; Jiménez, J. High clonal diversity of multidrug-resistant and extended spectrum beta-lactamase-producing Escherichia coli in a wastewater treatment plant. J. Environ. Manag. 2019, 245, 37–47. [Google Scholar] [CrossRef]
- Coutinho, F.H.; Silveira, C.B.; Pinto, L.H.; Salloto, G.R.B.; Cardoso, A.M.; Martins, O.B.; Vieira, R.P.; Clementino, M.M. Antibiotic Resistance is Widespread in Urban Aquatic Environments of Rio de Janeiro, Brazil. Microb. Ecol. 2014, 68, 441–452. [Google Scholar] [CrossRef]
- Chávez, M.; Cabezas, A.F.; Ferauds, M.; Castillo, J.E.; Caicedo, L.D. Antimicrobial resistance patterns and genotypic diversity between clinical and water systems isolates of Pseudomonas aeruginosa in Cali, Colombia. Trop. Biomed. 2020, 37, 650–662. [Google Scholar] [CrossRef]
- Alexander, J.; Bollmann, A.; Seitz, W.; Schwartz, T. Microbiological characterization of aquatic microbiomes targeting taxonomical marker genes and antibiotic resistance genes of opportunistic bacteria. Sci. Total. Environ. 2015, 512–513, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Han, I.; Yoo, K. Metagenomic Profiles of Antibiotic Resistance Genes in Activated Sludge, Dewatered Sludge and Bioaerosols. Water 2020, 12, 1516. [Google Scholar] [CrossRef]
- Bartley, P.S.; Domitrovic, T.N.; Moretto, V.T.; Santos, C.S.; Ponce-Terashima, R.; Reis, M.G.; Barbosa, L.M.; Blanton, R.E.; Bonomo, R.A.; Perez, F. Antibiotic Resistance in Enterobacteriaceae from Surface Waters in Urban Brazil Highlights the Risks of Poor Sanitation. Am. J. Trop. Med. Hyg. 2019, 100, 1369–1377. [Google Scholar] [PubMed]
- Leite, B.; De Chaves, M.A.; Nunes, A.A.T.; Jank, L.; Corção, G. Antibiotic resistance in surface waters from a coastal lagoon of Southern Brazil under the impact of anthropogenic activities. Ambient. Agua. Interdiscip. J. Appl. Sci. 2019, 14, 1–17. [Google Scholar] [CrossRef]
- Fang, H.; Huang, K.; Yu, J.; Ding, C.; Wang, Z.; Zhao, C.; Yuan, H.; Wang, Z.; Wang, S.; Hu, J.; et al. Metagenomic analysis of bacterial communities and antibiotic resistance genes in the Eriocheir sinensis freshwater aquaculture environment. Chemosphere 2019, 224, 202–211. [Google Scholar] [CrossRef]
- Ling, Z.; Yang, Y.; Huang, Y.; Zou, S.; Luan, T. A preliminary investigation on the occurrence and distribution of antibiotic resistance genes in the Beijiang River, South China. J. Environ. Sci. 2013, 25, 1656–1661. [Google Scholar] [CrossRef]
- Wang, M.; Shen, W.; Yan, L.; Wang, X.-H.; Xu, H. Stepwise impact of urban wastewater treatment on the bacterial community structure, antibiotic contents, and prevalence of antimicrobial resistance. Environ. Pollut. 2017, 231, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Lin, L.; Zhang, W.; Dong, L.; Yang, Y. Metagenome sequencing to unveil the resistome in a deep subtropical lake on the Yunnan-Guizhou Plateau, China. Environ. Pollut. 2020, 263, 114470. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yuan, K.; Chen, X.; Yang, Y.; Zhang, T.; Wang, Y.; Luan, T.; Zou, S.; Li, X. Metagenomic Analysis Revealing Antibiotic Resistance Genes (ARGs) and Their Genetic Compartments in the Tibetan Environment. Environ. Sci. Technol. 2016, 50, 6670–6679. [Google Scholar] [CrossRef]
- Fresia, P.; Antelo, V.; Salazar, C.; Giménez, M.; D’Alessandro, B.; Afshinnekoo, E.; Mason, C.; Gonnet, G.H.; Iraola, G. City-wide metagenomics uncover antibiotic resistance reservoirs in urban beach and sewage waters. bioRxiv 2018, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Eduardo-Correia, B.; Morales-Filloy, H.; Abad, J.P. Bacteria from the Multi-Contaminated Tinto River Estuary (SW, Spain) Show High Multi-Resistance to Antibiotics and Point to Paenibacillus spp. as Antibiotic-Resistance-Dissemination Players. Front. Microbiol. 2020, 10, 3071. [Google Scholar] [CrossRef]
- Lambirth, K.; Tsilimigras, M.; Lulla, A.; Johnson, J.; Al-Shaer, A.; Wynblatt, O.; Sypolt, S.; Brouwer, C.; Clinton, S.; Keen, O.; et al. Microbial Community Composition and Antibiotic Resistance Genes within a North Carolina Urban Water System. Water 2018, 10, 1539. [Google Scholar] [CrossRef] [Green Version]
- Picão, R.C.; Cardoso, J.P.; Campana, E.H.; Nicoletti, A.G.; Petrolini, F.V.; Assis, D.M.; Juliano, L.; Gales, A.C. The route of antimicrobial resistance from the hospital effluent to the environment: Focus on the occurrence of KPC-producing Aeromonas spp. and Enterobacteriaceae in sewage. Diagn. Microbiol. Infect. Dis. 2013, 76, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Fontes, L.C.; Neves, P.R.; Oliveira, S.; Silva, K.C.; Hachich, E.M.; Sato, M.I.Z.; Lincopan, N. Isolation of Pseudomonas aeruginosa Coproducing Metallo-β-Lactamase SPM-1 and 16S rRNA Methylase RmtD1 in an Urban River. Antimicrob. Agents Chemother. 2011, 55, 3063–3064. [Google Scholar] [CrossRef] [Green Version]
- Clarke, A.; Azulai, D.; Dueker, M.E.; Vos, M.; Perron, G.G. Triclosan Alters Microbial Communities in Freshwater Microcosms. Water 2019, 11, 961. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, A.J.F.C.; De França, P.T.R.; Pinto, A.B. Antimicrobial resistance of heterotrophic marine bacteria isolated from seawater and sands of recreational beaches with different organic pollution levels in southeastern Brazil: Evidences of resistance dissemination. Environ. Monit. Assess. 2009, 169, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Miranda, C.D.; Rojas, R.; Garrido, M.; Geisse, J.; Gonzalez, G. Role of shellfish hatchery as a reservoir of antimicrobial resistant bacteria. Mar. Pollut. Bull. 2013, 74, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, Y.; Wu, J. Continental-scale spatio-temporal distribution of antibiotic resistance genes in coastal waters along coastline of China. Chemosphere 2020, 247, 125908. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, R.; Zhang, M.; Cheng, Z.; Li, J.; Zhang, G.; Chen, B.; Zou, S.; Yang, Y. Exploring the differences of antibiotic resistance genes profiles between river surface water and sediments using metagenomic approach. Ecotoxicol. Environ. Saf. 2018, 161, 64–69. [Google Scholar] [CrossRef]
- Qu, J.; Jia, C.; Liu, Q.; Li, Z.; Liu, P.; Yang, M.; Zhao, M.; Li, W.; Zhu, H.; Zhang, Q. Dynamics of Bacterial Community Diversity and Structure in the Terminal Reservoir of the South-To-North Water Diversion Project in China. Water 2018, 10, 709. [Google Scholar] [CrossRef] [Green Version]
- Laffite, A.; Al Salah, D.M.M.; Slaveykova, V.I.; Otamonga, J.-P.; Poté, J. Impact of anthropogenic activities on the occurrence and distribution of toxic metals, extending-spectra β-lactamases and carbapenem resistance in sub-Saharan African urban rivers. Sci. Total. Environ. 2020, 727, 138129. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, Y.; An, W.; Lu, J.; Hu, J.; Yang, M. Antibiotic resistomes in drinking water sources across a large geographical scale: Multiple drivers and co-occurrence with opportunistic bacterial pathogens. Water Res. 2020, 183, 116088. [Google Scholar] [CrossRef]
- Chen, B.; Liang, X.; Nie, X.; Huang, X.; Zou, S.; Li, X. The role of class I integrons in the dissemination of sulfonamide resistance genes in the Pearl River and Pearl River Estuary, South China. J. Hazard. Mater. 2015, 282, 61–67. [Google Scholar] [CrossRef]
- Zhang, R.-Q.; Ying, G.-G.; Su, H.-C.; Zhou, L.-J.; Liu, Y.-S. Antibiotic resistance and genetic diversity ofEscherichia coliisolates from traditional and integrated aquaculture in South China. J. Environ. Sci. Health Part B 2013, 48, 999–1013. [Google Scholar] [CrossRef]
- Klase, G.; Lee, S.; Liang, S.; Kim, J.; Zo, Y.-G.; Lee, J. The microbiome and antibiotic resistance in integrated fishfarm water: Implications of environmental public health. Sci. Total. Environ. 2019, 649, 1491–1501. [Google Scholar] [CrossRef]
- Rodríguez, E.; Aristizábal-Hoyos, A.; Morales-Zapata, S.; Arias, L.; Jiménez, J. High frequency of gram-negative bacilli harboring blaKPC-2 in the different stages of wastewater treatment plant: A successful mechanism of resistance to carbapenems outside the hospital settings. J. Environ. Manag. 2020, 271, 111046. [Google Scholar] [CrossRef]
- Nakayama, T.; Hoa, T.T.T.; Harada, K.; Warisaya, M.; Asayama, M.; Hinenoya, A.; Lee, J.W.; Phu, T.M.; Ueda, S.; Sumimura, Y.; et al. Water metagenomic analysis reveals low bacterial diversity and the presence of antimicrobial residues and resistance genes in a river containing wastewater from backyard aquacultures in the Mekong Delta, Vietnam. Environ. Pollut. 2017, 222, 294–306. [Google Scholar] [CrossRef]
- Santoro, D.O.; Romão, C.M.; Clementino, M.M. Decreased aztreonam susceptibility amongPseudomonas aeruginosaisolates from hospital effluent treatment system and clinical samples. Int. J. Environ. Health Res. 2012, 22, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Serrato Díaz, A.; Cornejo Romero, A.; Amilpa Castro, O. La biodiversidad y evolución en ambientes acuáticos analizadas con herramientas moleculares. Hidrobiologica 2013, 23, 275–286. [Google Scholar]
- Mi, R.; Patidar, R.; Farenhorst, A.; Cai, Z.; Sepehri, S.; Khafipour, E.; Kumar, A. Detection of fecal bacteria and antibiotic resistance genes in drinking water collected from three First Nations communities in Manitoba, Canada. FEMS Microbiol. Lett. 2019, 366, 64–75. [Google Scholar] [CrossRef]
- Mittal, P.; Pk, V.P.; Dhakan, D.B.; Kumar, S.; Sharma, V.K. Metagenome of a polluted river reveals a reservoir of metabolic and antibiotic resistance genes. Environ. Microbiome 2019, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Marathe, N.P.; Pal, C.; Gaikwad, S.S.; Jonsson, V.; Kristiansson, E.; Larsson, D.J. Untreated urban waste contaminates Indian river sediments with resistance genes to last resort antibiotics. Water Res. 2017, 124, 388–397. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Baena, A.M.; Caicedo-Bejarano, L.D.; Chávez-Vivas, M. Structure of Bacterial Community with Resistance to Antibiotics in Aquatic Environments. A Systematic Review. Int. J. Environ. Res. Public Health 2021, 18, 2348. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18052348
Sánchez-Baena AM, Caicedo-Bejarano LD, Chávez-Vivas M. Structure of Bacterial Community with Resistance to Antibiotics in Aquatic Environments. A Systematic Review. International Journal of Environmental Research and Public Health. 2021; 18(5):2348. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18052348
Chicago/Turabian StyleSánchez-Baena, Ana María, Luz Dary Caicedo-Bejarano, and Mónica Chávez-Vivas. 2021. "Structure of Bacterial Community with Resistance to Antibiotics in Aquatic Environments. A Systematic Review" International Journal of Environmental Research and Public Health 18, no. 5: 2348. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18052348