1. Introduction
Male sexual debility is caused by the degenerative diseases in testicles. Male sexual debility is diagnosed when the general level of blood testosterone is below 3 µg L
−1 [
1]. With the quickening pace of modern life, the fierce competition of society, drug abuse, speedy increase of the aged population, and a variety of other reasons, the morbidity of male sexual debility is increasing steadily. Although this disease does not endanger the patient’s life, it can seriously lower the quality of life of the patients, affect the patients’ confidence, and damage the relationship between couples and thus affect the stability of the family. Therefore, the research on male sexual debility is of great significance to promote personal health and social harmony and has become a hot research topic.
Currently, the treatment of male sexual debility is focused on drug therapy, including testosterone replacement therapy [
2,
3,
4,
5] and phosphodiesterase-5 inhibitor therapy [
6,
7,
8,
9,
10]. However, these treatments have some limitations and have varying degrees of side effects [
11,
12,
13]. In recent years, nutritional interventions have received increasing attention, and there have been numerous studies showing that natural food ingredients are safe and effective in improving sexual function.
Ginseng (
Panax ginseng C.A. Meyer), a traditional Chinese medicinal herb, has been used as a restorative medicine for thousands of years [
14]. In 2012, Ministry of Public Health of China approved ginseng as new resource food [
15]. The ginseng root contains numerous bioactive components, such as ginsenoside, polysaccharides, amino acids, and peptides, that give it extensive biological activities including antioxidation [
16,
17,
18], anti-hypoxia effect [
19], immunoregulatory activity [
20,
21], hypolipidemic capacity [
22,
23], and the capacity to improve male sexual function [
24,
25,
26,
27]. Most studies attribute the effects of ginseng on male sexual function to ginsenoside [
28,
29].
Ostrea gigas Thunberg, also known as oyster, is rich in nutritional value and medicinal value. In 1758, the
Compendium of Materia Medica recorded its medical applications in improving male sexual function [
30]. Modern studies show that oysters can improve male sexual function by inhibiting the activity of phosphodiesterase-5 (PDE5) [
31]. However, the absorption rate of whole ginseng roots and oysters in human intestines is very low after oral administration, while oligopeptides, with low molecular weight, absorbable features, and high bioavailability, exist in whole ginseng roots and oysters as an important bioactive component. However, the effects of oligopeptides isolated from ginseng and oyster (GOPs and OOPs) on sexual function in male mice are rarely reported. Thus, the present study was designed to investigate the effects of GOPs, OOPs, and their combination on sexual function in male mice. This study may provide an important theoretical basis for exploring novel natural agents that improve male sexual function and may contribute to the further development and utilization of traditional ingredients such as ginseng and oysters.
2. Materials and Methods
2.1. Preparation and Identification of GOPs and OOPs
The GOP and OOP samples were provided by SinoMed Peptide Valley Bioengineering Co., Ltd. and International Bioactive Peptides Institute. GOPs and OOPs were derived from the Jilin ginseng root and oyster meat by enzymatic hydrolysis, respectively (Patent: CN105154509A, CN107997184A). In brief, fresh ginseng roots and oyster meat were cleansed, minced, homogenized in distilled water, and treated by complex protease (3000 U/g protein) at 40 °C for 3 h after adjusting the pH to 8.0 by sodium hydroxide. Next, nanofiltration, cryoconcentration, decolorization, purification, and spray drying were performed to obtain GOP and OOP powders. The powders were stored in a refrigerator at 4 °C under vacuum before being further processed.
The oligopeptide samples were purified by high-performance liquid chromatography (Waters Corporation, Milford, MA, USA) using a Phenomenex C18 column (10 mm × 250 mm). The mobile phase was acetonitrile 0.05 mol/L phosphate buffer (pH 3.2) (10:90) with a flow rate of 2.0 mL/min monitored by a Water 486 tunable UV detector at 208 nm. The molecular weight distribution of the GOP and OOP samples was measured by LDI-1700 matrix-assisted laser desorption ionization–time-of-flight mass spectrometry (MALDI-TOF-MS, Liner Scientific Inc., Reno, NV, USA).
2.2. Chemicals and Reagents
The whey protein sample was provided by China National Research Institute of Food and Fermen, and whey protein content was 80%. Assay kits used for the determination of nitric oxide (NO), testosterone, cyclic guanosine monophosphate (cGMP), and PDE5 were purchased from Andy Huatai Technology Co. LTD (Beijing, China). All the reagents used in this study were of analytical grade.
2.3. Animals
A total of 320 adult male ICR mice (18–22 g) and 160 adult female ICR mice (14–19 g) were obtained from the Animal Service of Health Science Center, Peking University. The environment was maintained at 25 ± 1 °C with a relative humidity of 50–60% and a 12 h/12 h light–dark cycle. All the mice had free access to an AIN-93G diet and water. Mice were allowed 7 days to adapt to the new environment. All animals were treated according to the Principle of Laboratory Animal Care (National Institutes of Health Publication No. 85-23, revised 1985) and the guidelines of the Peking University Animal Research Committee.
2.4. Groups and Treatment
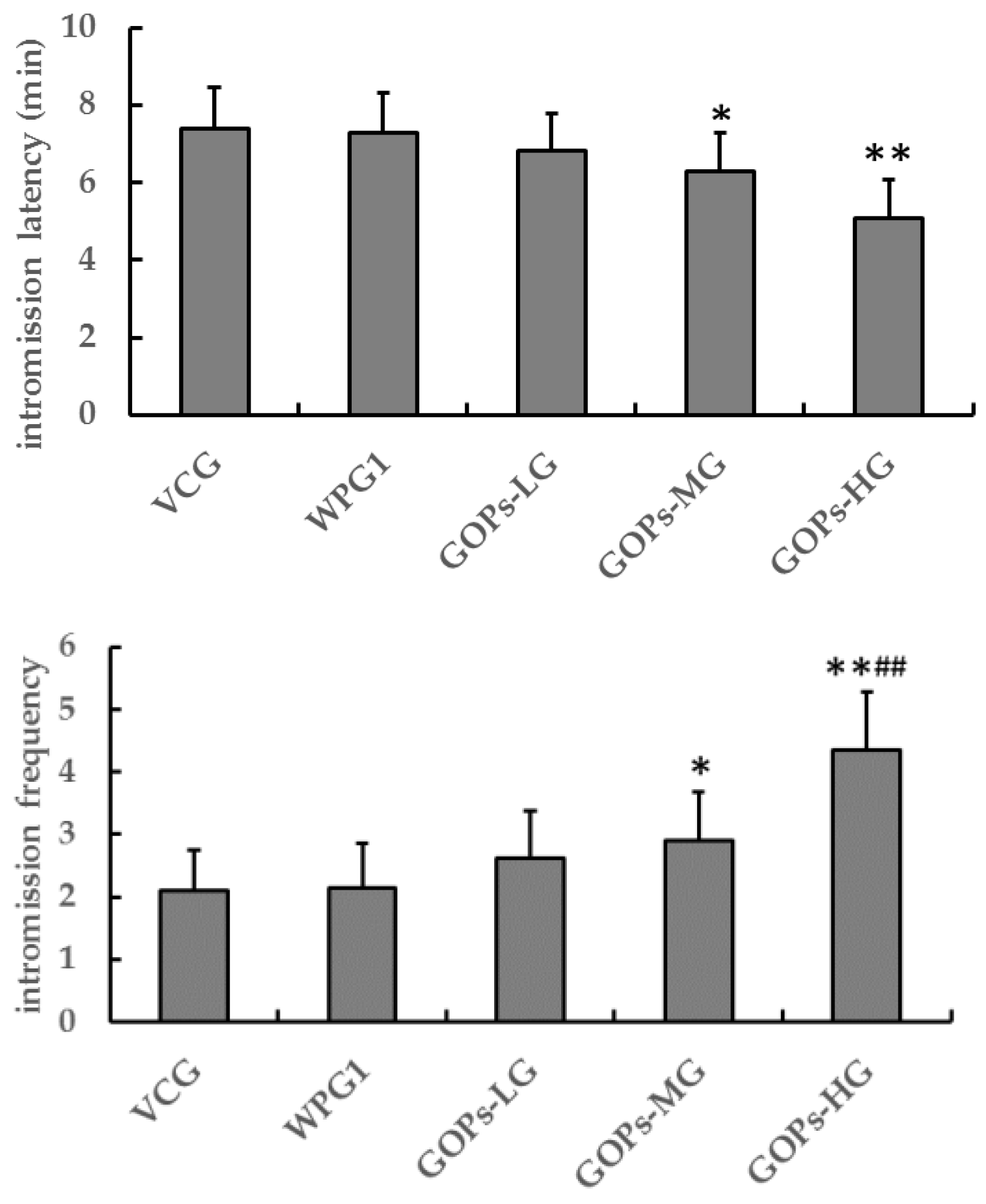
The first experiment was to explore the effect of GOPs on sexual function in male mice: Male mice were randomly divided into two subgroups (n = 50), namely subgroup A and subgroup B. Each subgroup was randomly divided into five groups (n = 10): vehicle control group (VCG), whey protein group 1 (125.0 mg kg−1, WPG1), and three groups of GOPs at different doses (62.5, 125.0, and 250.0 mg kg−1; GOPs-LG, GOPs-MG, and GOPs-HG, respectively). The VCG, WPG1, and three GOP groups were intragastrically administered vehicle, whey protein, and GOPs (0.1 mL/10 g), respectively. The animals were continuously treated for 30 days and then were used for further experiments.
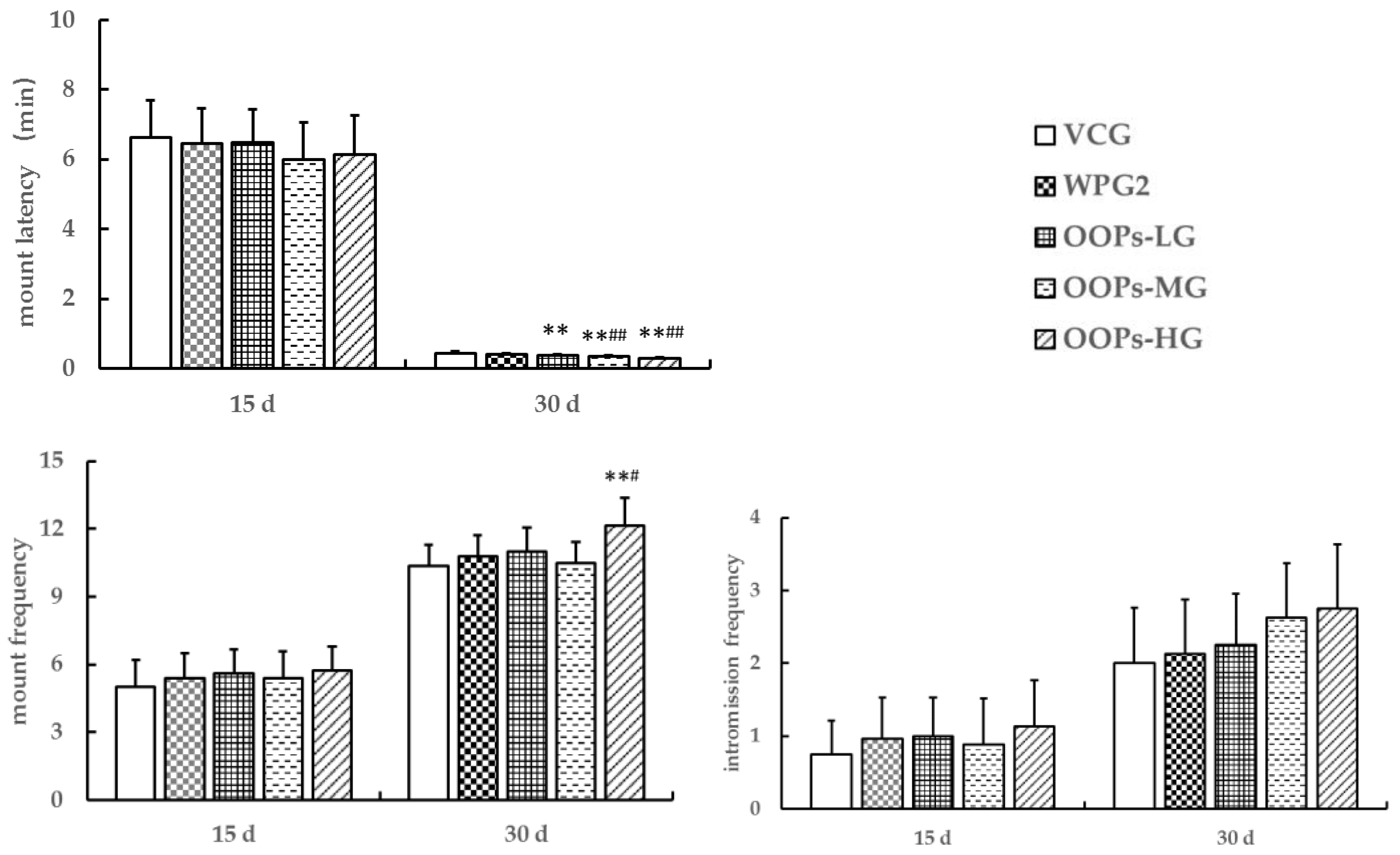
The second experiment was to explore the effect of OOPs on sexual function in male mice: Male mice were randomly divided into two subgroups (n = 50), namely subgroup A and subgroup B. Each subgroup was randomly divided into five groups (n = 10): vehicle control group (VCG), whey protein group 2 (160.0 mg kg−1, WPG2), and three groups of OOPs at different doses (80.0, 160.0, and 320.0 mg kg−1; OOPs-LG, OOPs-MG, and OOPs-HG, respectively). The VCG, WPG2, and three OOP groups were intragastrically administered vehicle, whey protein, and OOPs (0.1 mL/10 g), respectively. The animals were continuously treated for 30 days and then were used for further experiments.
The third experiment was to explore the combined effects of GOPs and OOPs on sexual function in male mice: Male mice were randomly divided into two subgroups (n = 60), namely subgroup A and subgroup B. Each subgroup was randomly divided into six groups (n = 10): vehicle control group (VCG), whey protein group 3 (222.5 mg kg−1, WPG3), and four groups of GOPs + OOPs at different doses (62.5 + 160.0, 62.5 + 320.0, 125.0 + 160.0, and 125.0 + 320.0 mg kg−1; GOPs + OOPs 1, GOPs + OOPs 2, GOPs + OOPs 3, and GOPs + OOPs 4, respectively). The VCG, WPG2, and four GOP + OOP groups were intragastrically administered vehicle, whey protein, and GOPs + OOPs (0.1 mL/10 g), respectively. The animals were continuously treated for 30 days and then were used for further experiments.
2.5. Sexual Behavior Study
In the first experiment, sexual behavior of male mice in subgroup A was observed on 30th day of dosing, and intromission latency and intromission frequency were recorded. In the second and third experiments, sexual behavior of male mice in subgroup A was observed on 15th and 30th day of dosing, and mount latency, mount frequency, and intromission frequency were recorded.
The detailed experimental method is as follows: Female mice were brought to estrus phase by administration of estradiol benzoate (250 μg/kg, i.h., administered 48 h prior to study; Sigma, St. Louis, MO, USA) and progesterone (250 μg/kg, i.h., administered 4 h prior to the study; Sigma) dissolved in corn oil. The sexual receptivity of female mice was evaluated before the study using a nonexperimental sexually vigorous male mouse [
32]. One hour after dosing, male mice were introduced to the observation chamber (30 cm × 15 cm × 15 cm) and allowed to adapt to the environment for 5 min (one mouse per cage). Then by slowly lifting upper glass lid, a female mouse in estrus was randomly selected and introduced into the cage, and the behavior parameters were observed. Mount latency refers to the time taken by the male mouse up to the first mount on female mouse, intromission latency refers to the time taken by the male mouse up to the first intromission, mount frequency refers to the number of mounts by a male mouse on a female mouse in 20 min, intromission frequency refers to the number of intromissions by a male mouse in 20 min. The experiment was conducted under weak light and quiet condition between 20:00 and 23:00.
2.6. Determination of Serum NO and Testosterone
Thirty minutes after the final oral administration, the retro-orbital blood collection was used to collect blood sample from the mice in subgroup B. The serum was obtained by centrifugation at 3500 rpm at 4 °C for 10 min. The NO and testosterone content in serum were measured by detection kits according to the instructions by nitrate reductase and ELISA method, respectively.
2.7. Determination of Sex Organ Indexes
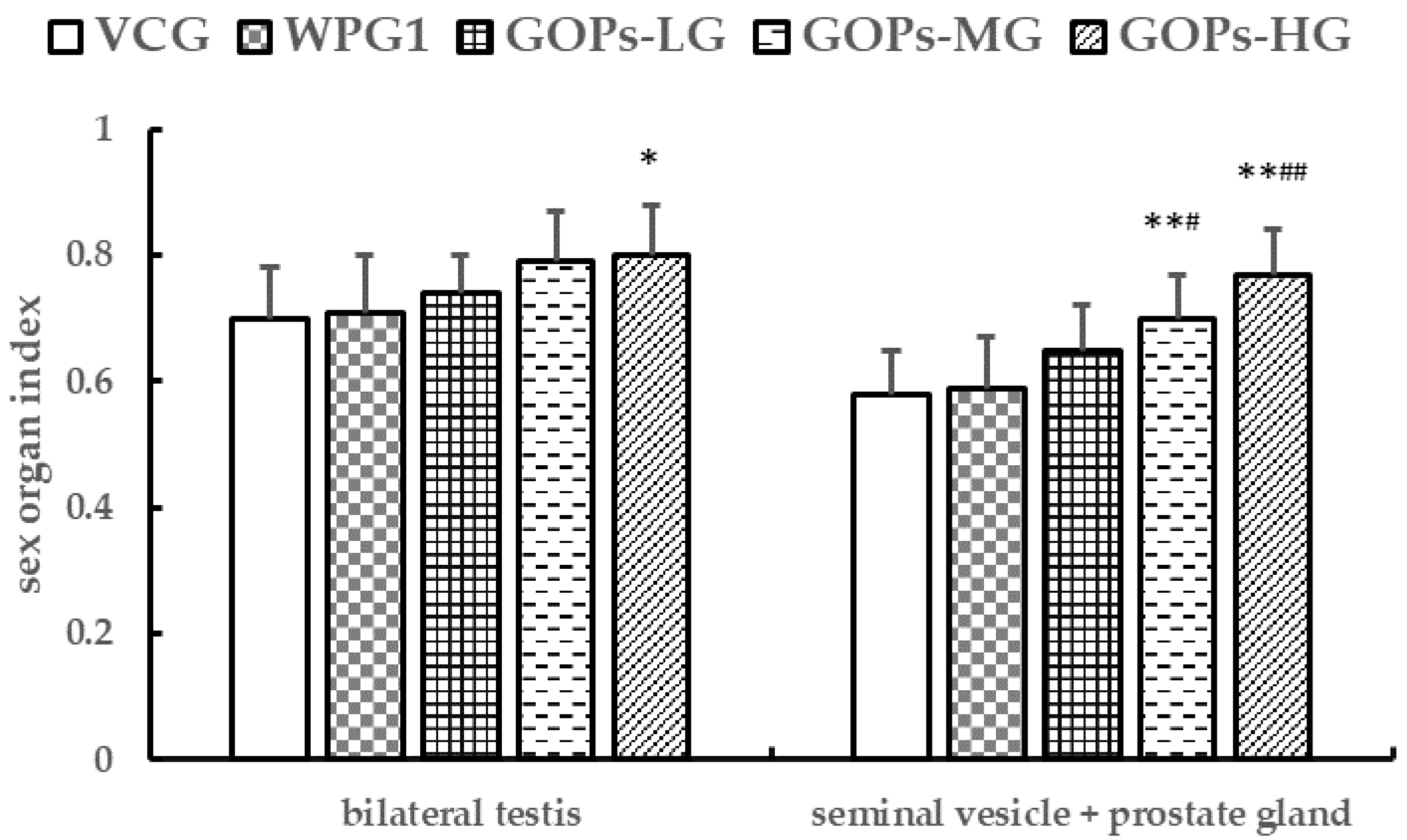
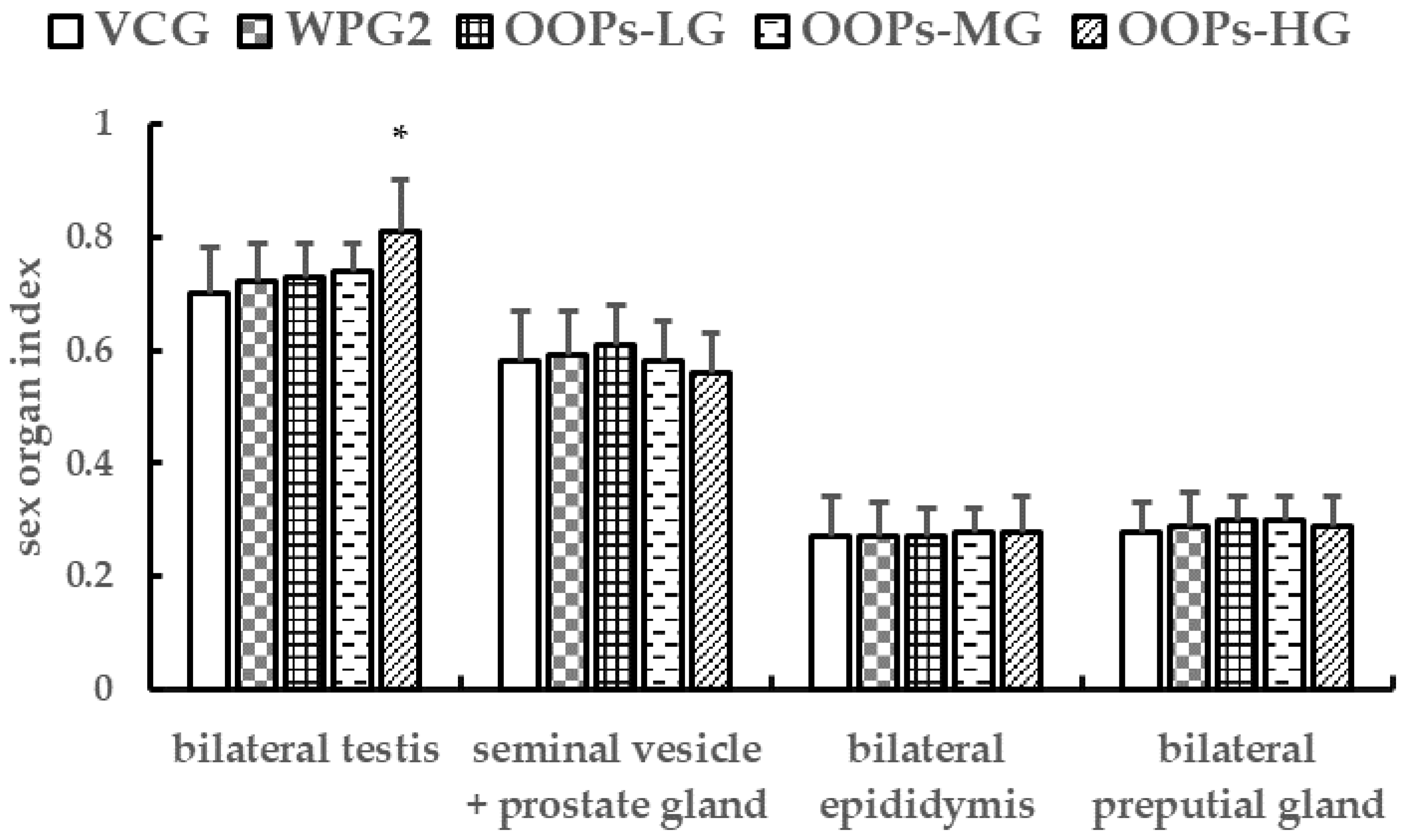
After the blood sample was obtained, the mice in subgroup B were sacrificed and the testis, epididymis, preputial glands, seminal vesicle and prostate glands were immediately isolated in an ice bath. Then the sex organs were weighed, and the organ indexes were calculated as weight of organ (g)/100 g body weight.
2.8. Examination of NO, cGMP, and PDE5 in Corpus Cavernosum Tissue
After the blood sample was obtained, the mice in subgroup B were sacrificed and the corpora cavernosa were immediately isolated in an ice bath. The concentrations of NO, cGMP and PDE5 were determined using available kits by Elisa method.
2.9. Statistical Analysis
The sexual behavior parameters are presented as mean ± standard error, and the other data are presented as mean ± standard deviation. Statistical significance with respect to vehicle was evaluated using one-way analysis of variance followed by Dunnett’s t-test, while the simple effects and interaction effects of GOPs and OOPs were evaluated by General Linear Model using SPSS software version 20 (SPSS Inc., Chicago, IL, USA). p < 0.05 was considered significant.
4. Discussion
With the fast development of society and economy, humans’ demands for better quality of life are becoming increasingly urgent. However, mental and physical exhaustion, tobacco use, and excessive alcohol consumption caused by the quickening pace of modern lives and fierce competition of society are driving an increase in the morbidity of male sexual debility [
33]. In the present study, we evaluated the effects of GOPs and OOPs on sexual function in male mice for the first time. Moreover, the combined effects of GOPs and OOPs were also further explored.
Whey protein is a catch-all term that includes several protein fractions such as β-lactoglobulin, α-lactalbumin, and immunoglobulin. Because it is easy to digest and absorb, whey protein has high bioavailability and various biological activities including immune support; fatigue resistance; and antioxidant, antibacterial, and antiviral activities [
15]. To exclude false-positive results caused by protein intake, we used whey protein as protein control by comparing oligopeptides to whey protein. According to the results we obtained from the current study, the effects of whey protein on sex function were not observed under our conditions, suggesting that the effects were due to oligopeptides, not protein intake.
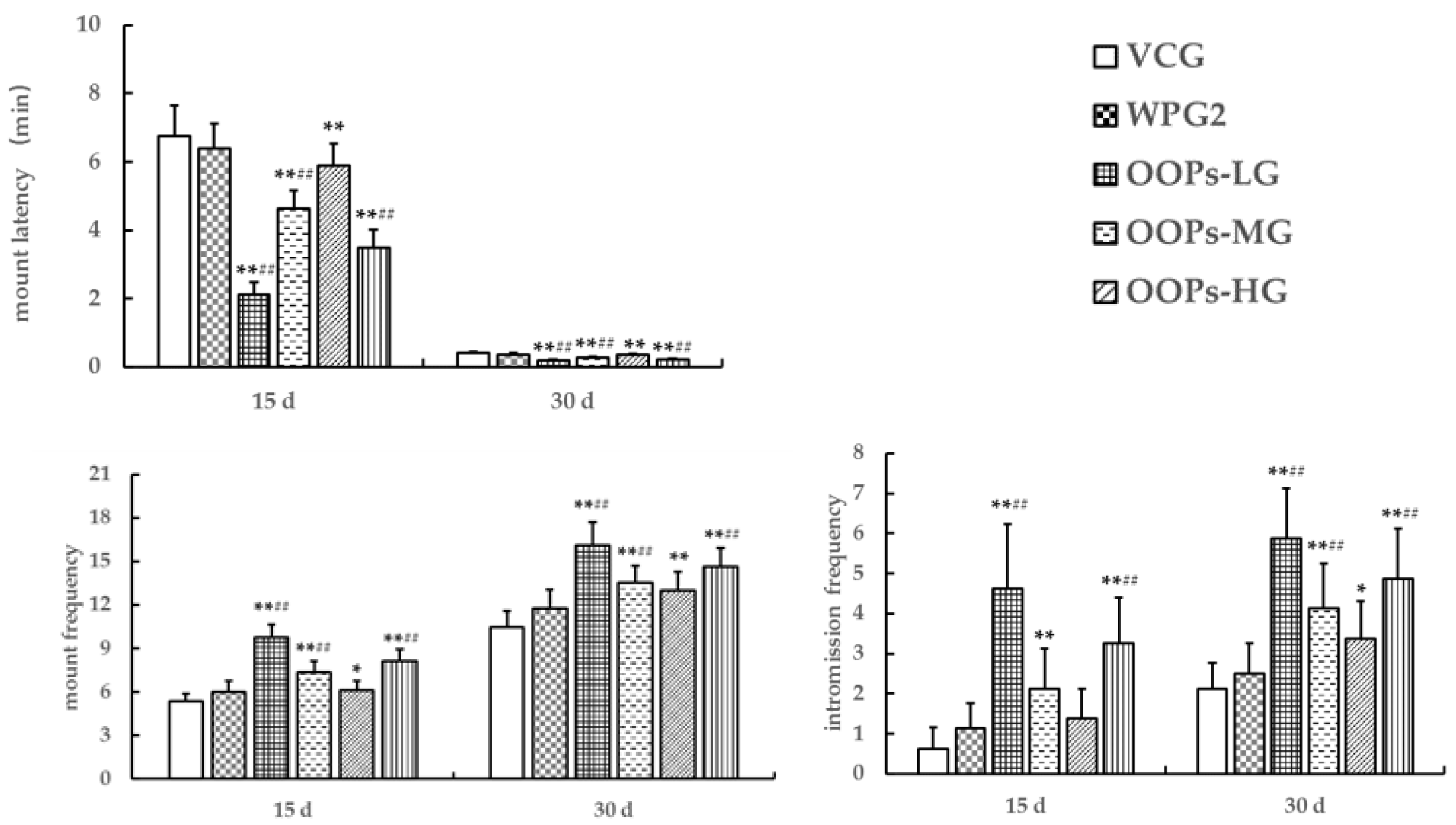
Male sexual function includes sexual desire, penile erection, sexual intercourse, orgasm, and ejaculation [
34]. Mounting is a measure of sexual motivation, whereas intromission is a reflection of the facilitation of sexual motivation [
35]. In the present study, GOPs at 250.0 mg kg
−1 showed the effects of decreasing intromission latency and increasing in intromission frequency, whereas GOPs at 62.5 or 125.0 mg kg
−1 had no significant effects on intromission latency and intromission frequency. Besides, we observed a decrease in mount latency in mice treated with OOPs at 160 or 320 mg kg
−1 and an increase in mount frequency in mice treated with OOPs at 320 mg kg
−1 but no increase in intromission frequency in mice treated with OOPs at 80.0, 160, or 320 mg kg
−1. These findings suggest that GOPs at 250.0 mg kg
−1 but not GOPs at 62.5 or 125.0 mg kg
−1 might improve male sexual function, and OOPs at 160 or 320 mg kg
−1 might only improve sexual motivation. In addition, mice treated with GOPs + OOPs at 62.5 + 160.0, 62.5 + 320.0, or 125.0 + 320.0 mg kg
−1 showed shorter mount latency and higher mount frequency, meanwhile, mice treated with GOPs + OOPs at 62.5 + 160.0 or 125.0 + 320.0 mg kg
−1 showed higher intromission frequency compared with the whey protein group on day 15 and day 30. It thus appeared that GOPs + OOPs at 62.5 + 160.0 or 125.0 + 320.0 mg kg
−1 might improve male sexual function, and GOPs + OOPs at 62.5 + 320.0 mg kg
−1 might only improve sexual motivation.
Studies on reproductive physiology and reproductive endocrinology have shown that the sexual activity is regulated by the neuroendocrine system, especially sexual hormone levels [
36]. Testosterone is an important sex hormone in the growth and development of the body, affecting the growth and development of germ cells as well as the development of sexual organs [
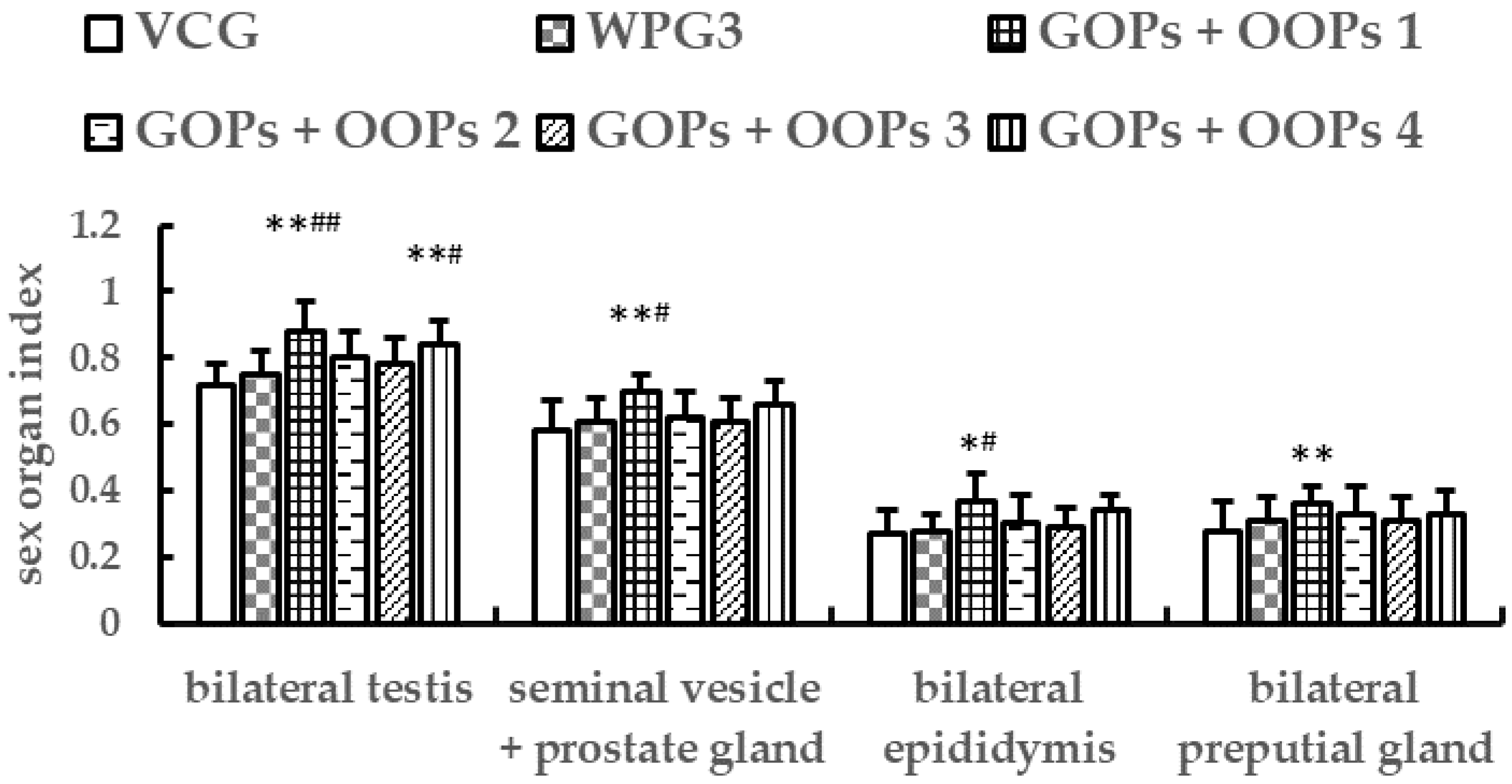
37]. The testicle is the most important organ for generating testosterone in mice, and testicular interstitial cells are the main synthetic cells. In the present study, GOPs at 250.0 mg kg
−1 resulted in an increase in the concentrations of serum testosterone, whereas GOPs at 62.5 or 125.0 mg kg
−1 and OOPs at 80.0, 160, or 320 mg kg
−1 had no significant increase. Besides, we observed an increase in bilateral testes index in mice treated with GOPs at 250.0 mg kg
−1 and OOPs at 320 mg kg
−1. These findings suggest that GOPs at 250.0 mg kg
−1 could elevate testosterone level, and OOPs at 320 mg kg
−1 might have the potential to elevate testosterone level. Furthermore, mice treated with GOPs + OOPs at 62.5 + 160.0, 62.5 + 320.0, or 125.0 + 320.0 mg kg
−1 showed an increase in the concentration of serum testosterone; meanwhile, mice treated with GOPs + OOPs at 62.5 + 160.0 or 125.0 + 320.0 mg kg
−1 showed an increase in bilateral testes index.
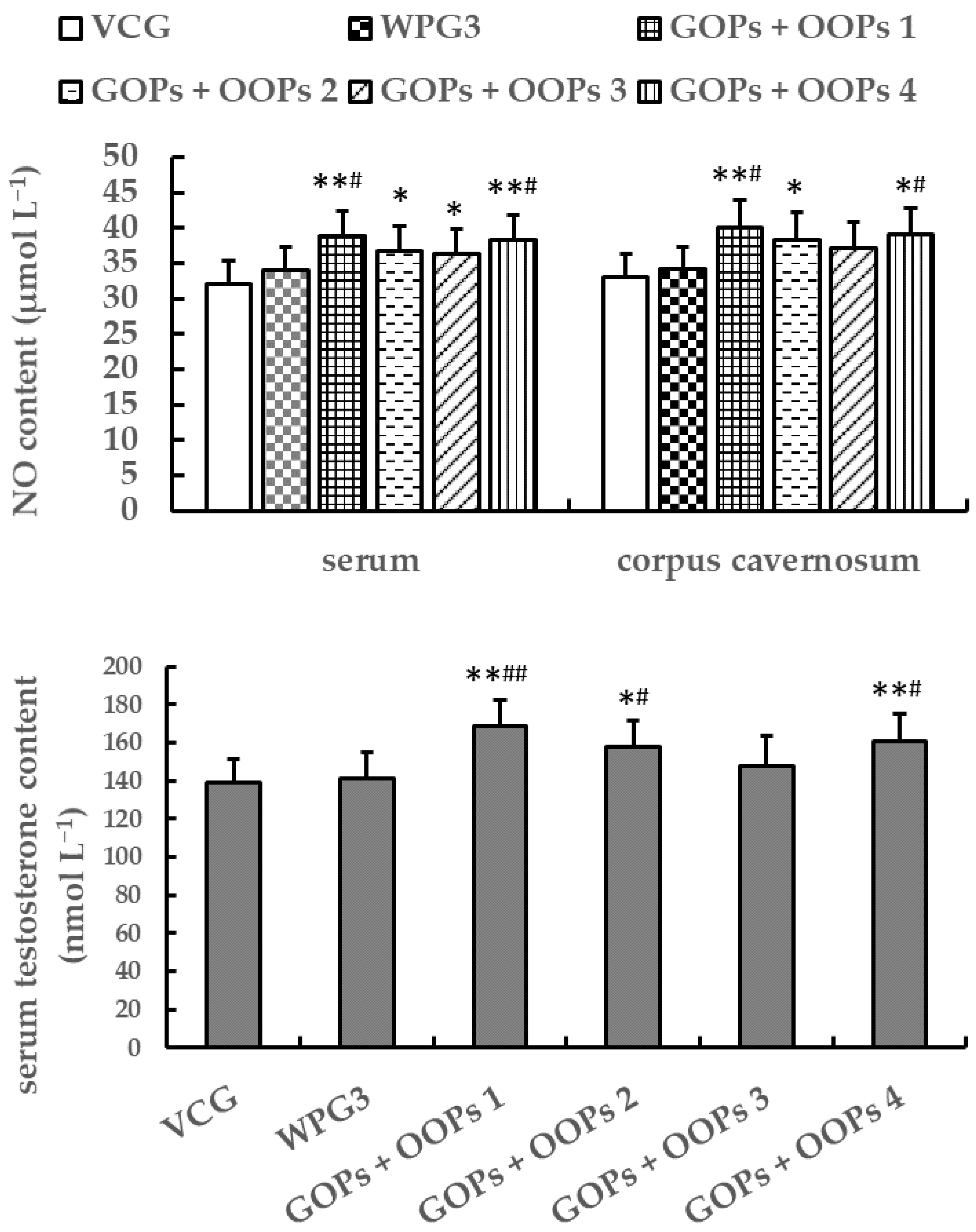
Penile erection is an important component of male sexual function, the mechanism of which is related to many neurotransmitters, ions, and enzymes [
38]. After diffusing into corpus cavernosum smooth muscle (CCSM), NO can stimulate soluble guanylyl cyclase (sGC) to generate cGMP from guanosine triphosphate (GTP). cGMP relaxes the CCSM by opening the potassium ion channel, thus increasing penis blood flow and erecting the penis [
7,
39]. Therefore, NO and cGMP are beneficial for penile erection. In the present study, mice treated with GOPs at 250.0 mg kg
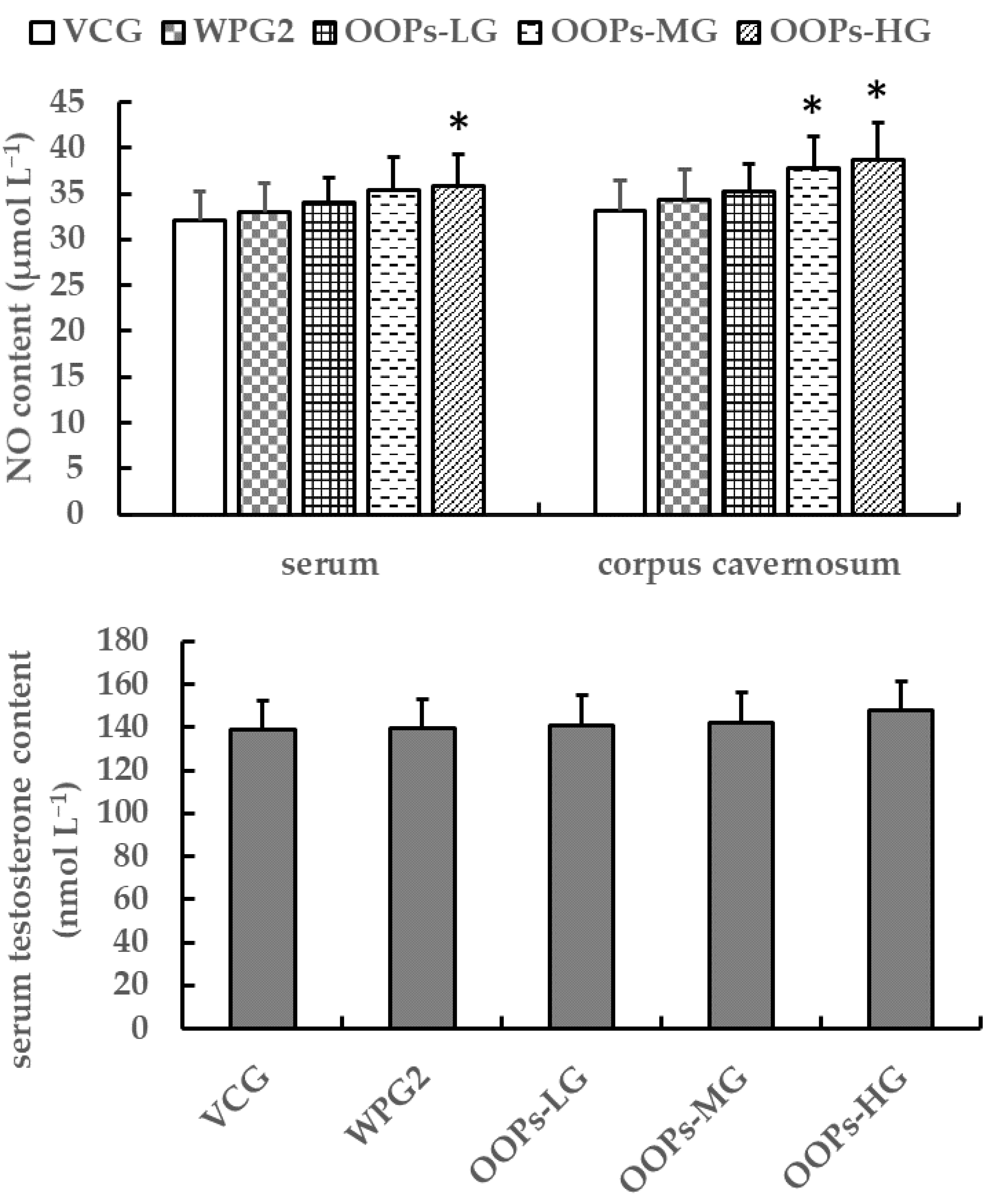
−1 showed an increase in the concentrations of serum NO, whereas those treated with GOPs at 62.5 or 125.0 mg kg
−1 and OOPs at 80.0, 160, or 320 mg kg
−1 had no significant increase. Besides, mice treated with GOPs + OOPs at 62.5 + 160.0 or 125.0 + 320.0 mg kg
−1 showed an increase in the concentrations of NO both in serum and corpus cavernosum. In short, GOPs at 62.5 or 125.0 mg kg
−1 and OOPs at 80.0, 160, or 320 mg kg
−1 had no effects on NO content, but GOPs + OOPs at 62.5 + 160.0 or 125.0 + 320.0 mg kg
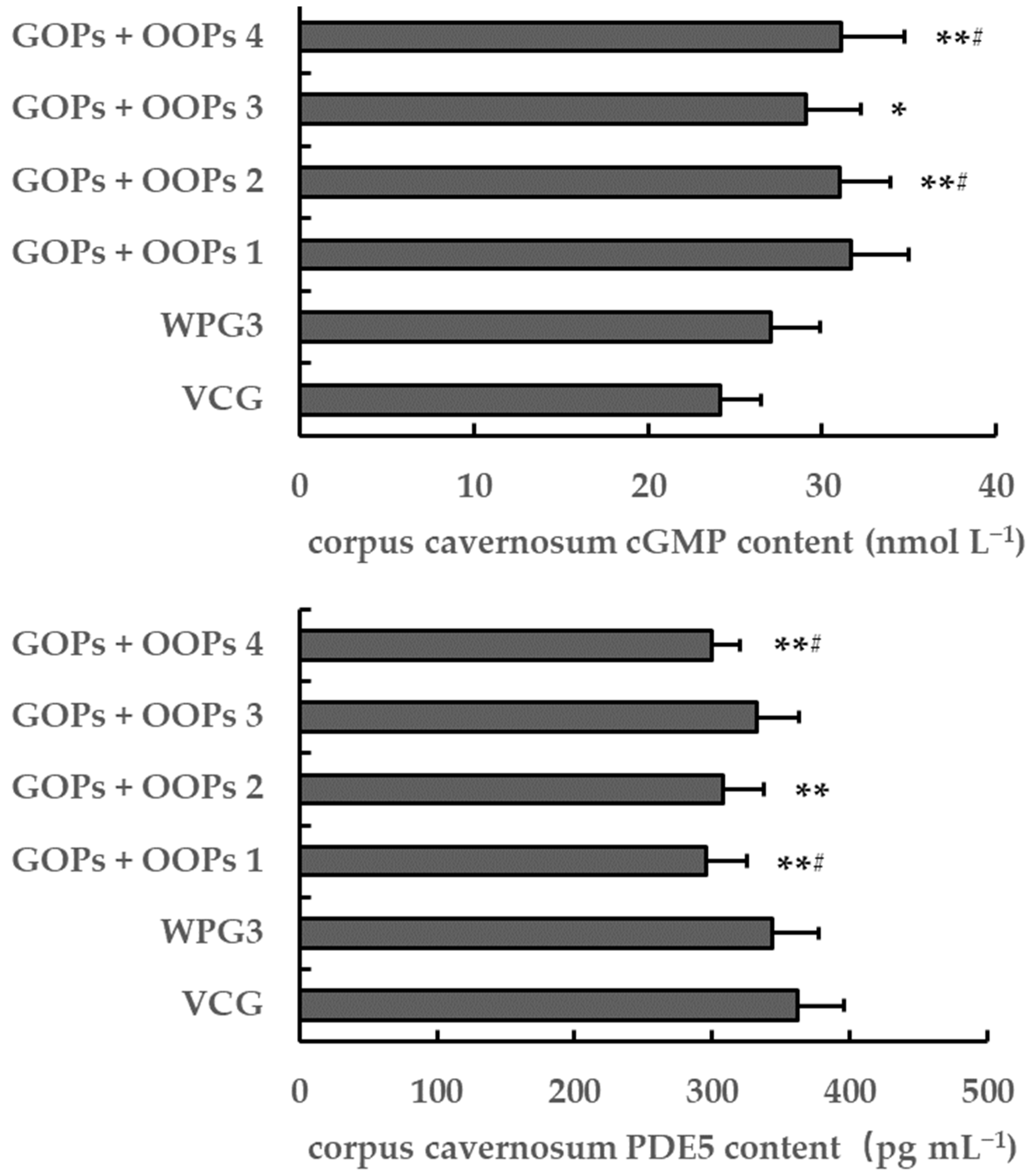
−1 did have effects on NO content. Moreover, we observed an increase in corpus cavernosum cGMP content in mice treated with GOPs + OOPs at 62.5 + 160.0, 62.5 + 320.0, or 125.0 + 320.0 mg kg
−1, which might explain the enhancement to penile erection.
Phosphodiesterases (PDEs) are a multigene family, among which PDE5 is closely related to the reproductive system and is mainly distributed in the corpus cavernosum and platelets. PDE5 specially hydrolyses cGMP [
7,
40,
41]. Therefore, PDE5 reduces the function of cGMP to relax CCSM, thus affecting penile erection. In the present study, mice treated with GOPs + OOPs at 62.5 + 160.0 or 125.0 + 320.0 mg kg
−1 showed a decrease in the corpus cavernosum PDE5 content, suggesting that these two dosages might be beneficial for penile erection.
Moreover, we analyzed the interaction effects of GOPs and OOPs. The results showed that GOPs and OOPs had interaction effects on bilateral testes index, seminal vesicle + prostate gland index, mount latency, mount frequency, intromission frequency, and serum testosterone content. The subsequent simple effects analysis results showed that there was significant difference between GOPs at 62.5 mg kg−1 and 125.0 mg kg−1 on the above variables when the dosage of OOPs was 160 mg kg−1. Interestingly, among the four combined groups, the best treatment for enhancing male sexual function was GOPs + OOPs at 62.5 + 160.0 mg kg−1, which obviously was the lowest dosage match.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}