
Muscle Oxygen Desaturation and Re-Saturation Capacity Limits in Repeated Sprint Ability Performance in Women Soccer Players: A New Physiological Interpretation

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
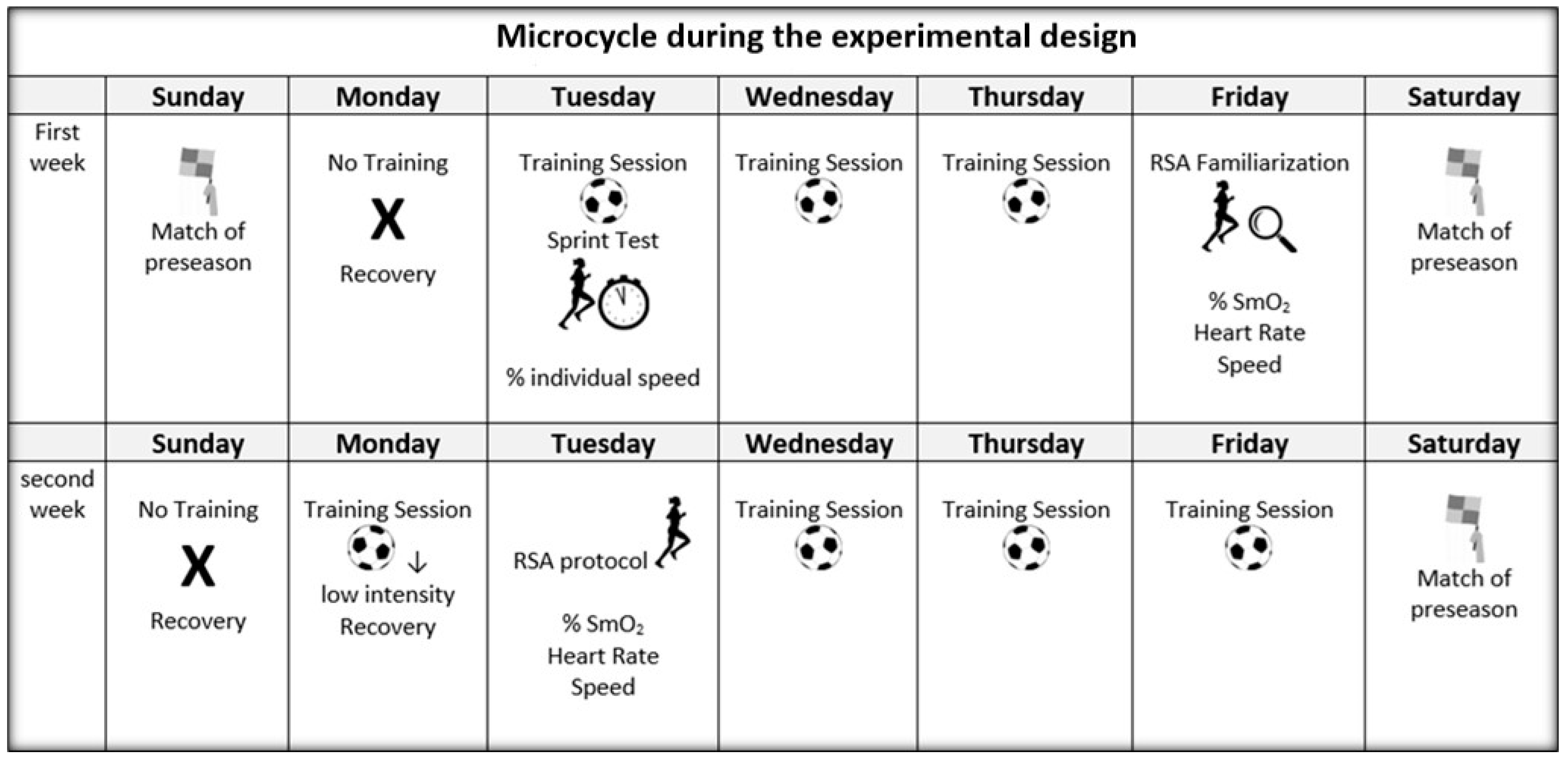
2.2. Experimental Design
2.3. Assessment
2.3.1. Repeated Sprint Ability Test
2.3.2. Individual Speed Zone
2.3.3. Heart Rate Zones
2.3.4. Relationship between External and Internal Load Measurements
2.4. Muscle Oxygenation Dynamics
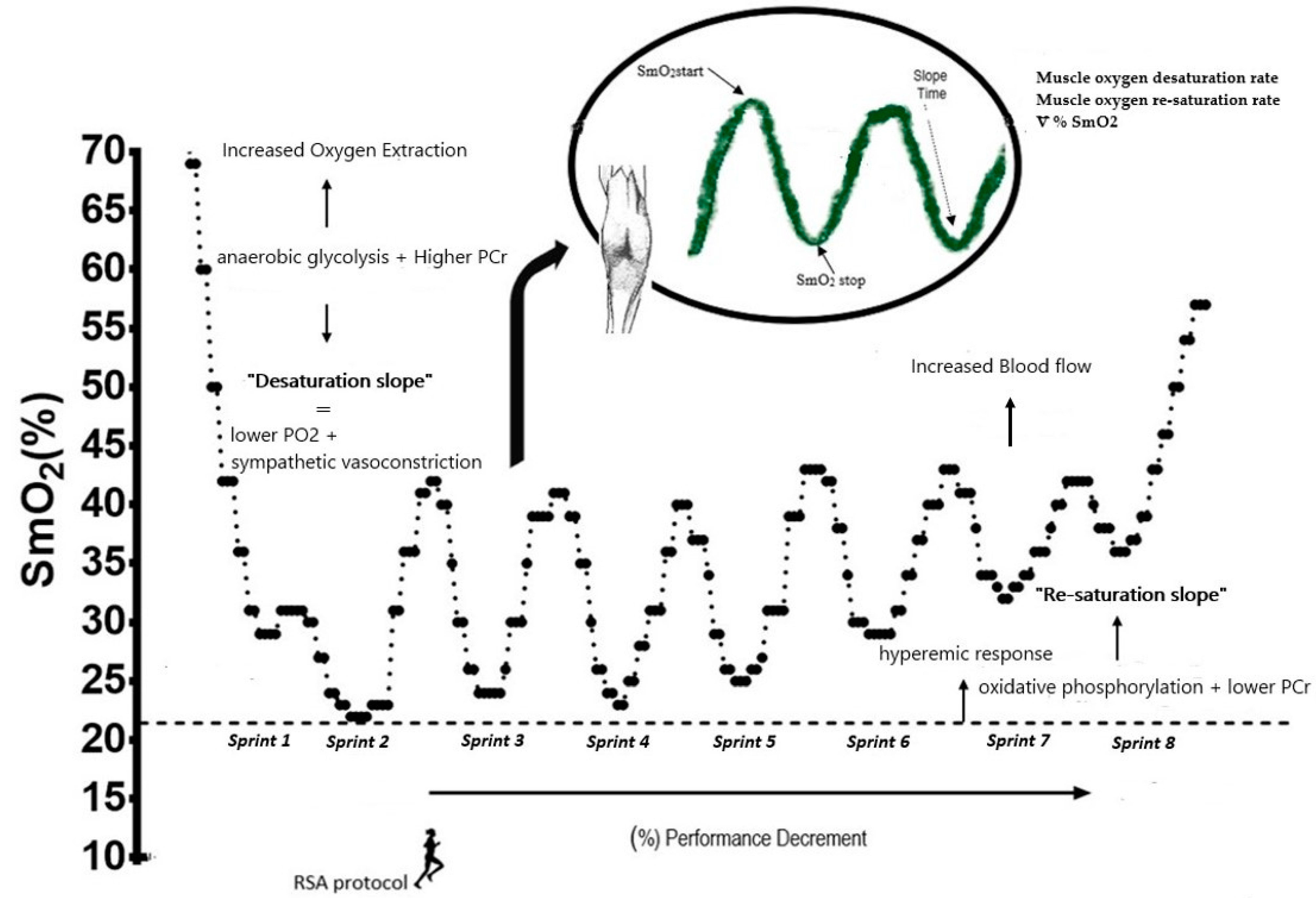
- Raw SmO2 values were calculated during desaturation and re-saturation of the last second of each of the eight sprints [10].
- The muscle oxygen desaturation rate was evaluated as the difference between the maximum (work interval) and minimum SmO2 values (rest interval) and divided by the duration of the work interval. Similarly, the muscle oxygen re-saturation rate was determined as the difference between the minimum (rest interval) and maximum SmO2 (working interval), divided by the rest duration of the interval (20 s) [10].
- The percentage of muscle oxygen extraction from the SmO2 desaturation and re- saturation values was obtained during each sprint using the difference between SmO2 at the start and end of the sprint. The SmO2 start value was considered 1 s before starting each series, while the SmO2 stop value was determined in the last second of the work interval of each sprint with the following formula: ∇%SmO2 = ((SmO2Stop × 100/SmO2start) – 100) x −1 [22].
2.5. Statical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lockie, R.G.; Moreno, M.R.; Orjalo, A.J.; Stage, A.A.; Liu, T.M.; Birmingham-Babauta, S.A.; Hurley, J.M.; Torne, I.A.; Beiley, M.D.; Risso, F.G.; et al. Repeated-Sprint Ability in Division i Collegiate Male Soccer Players: Positional Differences and Relationships with Performance Tests. J. Strength Cond. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.M.; Macpherson, T.W.; Spears, I.R.; Weston, M. Repeated sprints: An independent not dependent variable. Int. J. Sports Physiol. Perform. 2016, 11, 693–696. [Google Scholar] [CrossRef] [PubMed]
- Girard, O.; Mendez-Villanueva, A.; Bishop, D. Repeated-Sprint Ability—Part I. Sport. Med. 2011. [Google Scholar] [CrossRef]
- Nakamura, F.Y.; Pereira, L.A.; Loturco, I.; Rosseti, M.; Moura, F.A.; Bradley, P.S. Repeated-Sprint Sequences during Female Soccer Matches Using Fixed and Individual Speed Thresholds. J. Strength Cond. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bradley, P.S.; Vescovi, J.D. Velocity thresholds for women’s soccer matches: Sex specificity dictates high-speed-running and sprinting thresholds-female athletes in motion (FAiM). Int. J. Sports Physiol. Perform. 2015, 10, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Arrones, L.; Torreño, N.; Requena, B.; Sáez De Villarreal, E.; Casamichana, D.; Barbero-Alvarez, J.C.; Munguía-Izquierdo, D. Match-play activity proile in professional soccer players during oficial games and the relationship between external and internal load. J. Sports Med. Phys. Fitness 2015, 55, 1417–1422. [Google Scholar] [PubMed]
- McManus, C.J.; Collison, J.; Cooper, C.E. Performance comparison of the MOXY and PortaMon near-infrared spectroscopy muscle oximeters at rest and during exercise. J. Biomed. Opt. 2018, 23, 1. [Google Scholar] [CrossRef]
- Azevedo, R.D.A.; Béjar Saona, J.E.; Inglis, E.C.; Iannetta, D.; Murias, J.M. The effect of the fraction of inspired oxygen on the NIRS-derived deoxygenated hemoglobin “breakpoint” during ramp-incremental test. Am. J. Physiol. Integr. Comp. Physiol. 2020, 318, R399–R409. [Google Scholar] [CrossRef]
- McLean, S.; Kerhervé, H.; Lovell, G.P.; Gorman, A.D.; Solomon, C. The effect of recovery duration on vastus lateralis oxygenation, heart rate, perceived exertion and time motion descriptors during small sided football games. PLoS ONE 2016. [Google Scholar] [CrossRef] [Green Version]
- Buchheit, M.; Cormie, P.; Abbiss, C.R.; Ahmaidi, S.; Nosaka, K.K.; Laursen, P.B. Muscle deoxygenation during repeated sprint running: Effect of active vs. Passive recovery. Int. J. Sports Med. 2009. [Google Scholar] [CrossRef]
- Brocherie, F.; Millet, G.P.; Girard, O. Neuro-mechanical and metabolic adjustments to the repeated anaerobic sprint test in professional football players. Eur. J. Appl. Physiol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Haseler, L.J.; Hogan, M.C.; Richardson, R.S. Skeletal muscle phosphocreatine recovery in exercise-trained humans is dependent on O2 availability. J. Appl. Physiol. 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabbett, T.J. The development of a test of repeated-sprint ability for elite women’s soccer players. J. Strength Cond. Res. 2010. [Google Scholar] [CrossRef] [PubMed]
- Datson, N.; Weston, M.; Drust, B.; Gregson, W.; Lolli, L. High-intensity endurance capacity assessment as a tool for talent identification in elite youth female soccer. J. Sports Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zagatto, A.M.; Beck, W.R.; Gobatto, C.A. Validity of the running anaerobic sprint test for assessing anaerobic power and predicting short-distance performances. J. Strength Cond. Res. 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, A.R.; Mukherjee, S.; Chia, M.Y.H.; Teh, K.C. Validity of the running repeated sprint ability test among playing positions and level of competitiveness in trained soccer players. Int. J. Sports Med. 2008. [Google Scholar] [CrossRef]
- Karvonen, M.J.; Kentala, E.; Mustala, O. The effects of training on heart rate; a longitudinal study. Ann. Med. Exp. Biol. Fenn. 1957, 35, 307–315. [Google Scholar]
- Ihsan, M.; Watson, G.; Lipski, M.; Abbiss, C.R. Influence of postexercise cooling on muscle oxygenation and blood volume changes. Med. Sci. Sports Exerc. 2013. [Google Scholar] [CrossRef]
- Rodriguez, R.F.; Townsend, N.E.; Aughey, R.J.; Billaut, F. Influence of averaging method on muscle deoxygenation interpretation during repeated-sprint exercise. Scand. J. Med. Sci. Sport. 2018. [Google Scholar] [CrossRef]
- Bangde, W.; Guodong, X.; Qingping, T.; Jinyan, S.; Bailei, S.; Lei, Z.; Qingming, L.; Hui, G.; Wang, B.; Xu, G.; et al. Differences between the vastus lateralis and gastrocnemius lateralis in the assessment ability of breakpoints of muscle oxygenation for aerobic capacity indices during an incremental cycling exercise. J. Sport. Sci. Med. 2012, 11, 606–613. [Google Scholar]
- Feldmann, A.; Schmitz, R.W.; Erlacher, D. Near-infrared spectroscopy-derived muscle oxygen saturation on a 0% to 100% scale: Reliability and validity of the Moxy Monitor. J. Biomed. Opt. 2019, 24, 115001. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Carmona, C.D.; Bastida-Castillo, A.; Rojas-Valverde, D.; de la Cruz Sánchez, E.; García-Rubio, J.; Ibáñez, S.J.; Pino-Ortega, J. Lower-limb Dynamics of Muscle Oxygen Saturation During the Back-squat Exercise: Effects of Training Load and Effort Level. J. strength Cond. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Buchheit, M.; Abbiss, C.R.; Peiffer, J.J.; Laursen, P.B. Performance and physiological responses during a sprint interval training session: Relationships with muscle oxygenation and pulmonary oxygen uptake kinetics. Eur. J. Appl. Physiol. 2012. [Google Scholar] [CrossRef]
- Hecksteden, A.; Pitsch, W.; Rosenberger, F.; Meyer, T. Repeated testing for the assessment of individual response to exercise training. J. Appl. Physiol. 2018. [Google Scholar] [CrossRef]
- Ross, R.; Goodpaster, B.H.; Koch, L.G.; Sarzynski, M.A.; Kohrt, W.M.; Johannsen, N.M.; Skinner, J.S.; Castro, A.; Irving, B.A.; Noland, R.C.; et al. Precision exercise medicine: Understanding exercise response variability. Br. J. Sports Med. 2019. [Google Scholar] [CrossRef] [Green Version]
- Alexiou, H.; Coutts, A.J. A comparison of methods used for quantifying internal training load in women soccer players. Int. J. Sports Physiol. Perform. 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beato, M.; Drust, B. Acceleration intensity is an important contributor to the external and internal training load demands of repeated sprint exercises in soccer players. Res. Sport. Med. 2020. [Google Scholar] [CrossRef]
- McGawley, K.; Bishop, D.J. Oxygen uptake during repeated-sprint exercise. J. Sci. Med. Sport 2015. [Google Scholar] [CrossRef] [Green Version]
- Dawson, B.; Goodman, C.; Lawrence, S.; Preen, D.; Polglaze, T.; Fitzsimons, M.; Fournier, P. Muscle phosphocreatine repletion following single and repeated short sprint efforts. Scand. J. Med. Sci. Sports 2007. [Google Scholar] [CrossRef]
- Whipp, B.J.; Ward, S.A.; Rossiter, H.B. Pulmonary O2 uptake during exercise: Conflating muscular and cardiovascular responses. Med. Sci. Sports Exerc. 2005, 37, 1574–1585. [Google Scholar] [CrossRef]
- Billaut, F.; Bishop, D. Muscle fatigue in males and females during multiple-sprint exercise. Sport. Med. 2009, 39, 257–278. [Google Scholar] [CrossRef]
- Woorons, X.; Mucci, P.; Aucouturier, J.; Anthierens, A.; Millet, G.P. Acute effects of repeated cycling sprints in hypoxia induced by voluntary hypoventilation. Eur. J. Appl. Physiol. 2017. [Google Scholar] [CrossRef]
- Lapointe, J.; Paradis-Deschênes, P.; Woorons, X.; Lemaître, F.; Billaut, F. Impact of Hypoventilation Training on Muscle Oxygenation, Myoelectrical Changes, Systemic [K+], and Repeated-Sprint Ability in Basketball Players. Front. Sport. Act. Living 2020, 2, 29. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.F.; Townsend, N.E.; Aughey, R.J.; Billaut, F. Muscle oxygenation maintained during repeated-sprints despite inspiratory muscle loading. PLoS ONE 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murias, J.M.; Spencer, M.D.; Keir, D.A.; Paterson, D.H. Systemic and vastus lateralis muscle blood flow and O2 extraction during ramp incremental cycle exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.D.; Victor, R.G. Nitric oxide mediates contraction-induced attenuation of sympathetic vasoconstriction in rat skeletal muscle. J. Physiol. 1998. [Google Scholar] [CrossRef]
- Calaine Inglis, E.; Iannetta, D.; Murias, J.M. The plateau in the NIRS-derived [HHb] signal near the end of a ramp incremental test does not indicate the upper limit of O2 extraction in the vastus lateralis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017. [Google Scholar] [CrossRef]
- Rampinini, E.; Bishop, D.; Marcora, S.M.; Ferrari Bravo, D.; Sassi, R.; Impellizzeri, F.M. Validity of simple field tests as indicators of match-related physical performance in top-level professional soccer players. Int. J. Sports Med. 2007. [Google Scholar] [CrossRef] [PubMed]
- Bendiksen, M.; Pettersen, S.A.; Ingebrigtsen, J.; Randers, M.B.; Brito, J.; Mohr, M.; Bangsbo, J.; Krustrup, P. Application of the Copenhagen Soccer Test in high-level women players—Locomotor activities, physiological response and sprint performance. Hum. Mov. Sci. 2013. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Sprint 1 | Sprint 2 | Sprint 3 | Sprint 4 | Sprint 5 | Sprint 6 | Sprint 7 | Sprint 8 | N2 Partial/Factor |
|---|---|---|---|---|---|---|---|---|---|
| Time (s) | 3.98 ± 0.28 | 3.94 ± 0.17 | 4.01 ± 0.26 | 4.11 ± 0.30 | 4.07 ± 0.20 | 4.19 ± 0.30 | 4.14 ± 0.33 | 4.18 ± 0.35 | 0.484 |
| % Change | BL | −1.0 | 0.8 | 3.3 | 2.3 | 5.3 ↑↔ * | 4 ↑↔ * | 5 ↑↔ * | Linear |
| Power (w) | 396 ± 97 | 399 ± 70 | 388 ± 96 | 359 ± 94 | 366 ± 72 | 343 ± 93 | 356 ± 107 | 348 ± 101 | 0.835 |
| % Change | BL | 0.8 | −2 | −9.3 ↓ ** | −7.6 ↓ ** | −13.4 ↓ *** | −10.1 ↓ *** | −12.1 ↓ *** | Linear |
| Heart Rate (pmm) | 144 ± 13 | 159 ± 6 | 165 ± 6 | 170 ± 7 | 172 ± 4 | 174 ± 4 | 175 ± 5 | 173 ± 6 | 0.952 |
| % Change | BL | 10.4 ↑ *** | 14.6 ↑ *** | 18.1 ↑ *** | 19.4 ↑ *** | 21.2 ↑ *** | 21.9 ↑ *** | 20.6 ↑ *** | Linear |
| Heart Rate Zone (%) | 61 ± 11 | 73 ± 5 | 78 ± 5 | 82 ± 6 | 84 ± 4 | 85 ± 3 | 87 ± 4 | 85 ± 5 | 0.952 |
| % Change | BL | 19.7 ↑ *** | 27.9↑ *** | 34.4↑ *** | 37.7↑ *** | 41↑ *** | 42.6↑ *** | 39.3↑ *** | Linear |
| Speed (km/h) | 18.5 ± 1.2 | 18.7 ± 0.8 | 18.0 ± 1.2 | 17.6 ± 1.3 * | 17.7 ± 0.9 | 17.3 ± 1.2 ** | 17.5 ± 1.3 | 17.3 ± 1.4 | 0.672 |
| % Change | BL | 1.1 | −2.7 | −4.9 ↓ *** | −4.3 ↓ ** | −6.5 ↓ *** | −5.4 ↓ *** | −6.5 ↓ *** | Linear |
| Individual Speed (%) | 95 ± 4 | 95 ± 3 | 94 ± 4 | 92 ± 5 | 93 ± 5 | 90 ± 5 | 91 ± 5 | 90 ± 6 | 0.672 |
| % Change | BL | 0 | −1.1 | −3.2 ↓ ** | −2.1 ↓ ** | −5.3 ↓ *** | −4.2 ↓ *** | −5.3 ↓ *** | Linear |
| SmO2 Dynamics | Sprint 1 | Sprint 2 | Sprint 3 | Sprint 4 | Sprint 5 | Sprint 6 | Sprint 7 | Sprint 8 | N2 Partial Square |
|---|---|---|---|---|---|---|---|---|---|
| Time Total (Sec) | 3.98 ± 0.28 | 23.94 ± 0.17 | 47.95 ± 0.39 | 72.12 ± 0.65 | 96.14 ± 0.74 | 120.33 ± 0.98 | 144.47 ± 1.23 | 168.65 ± 1.52 | |
| SmO2 desaturation | 45 ± 22 | 34 ± 17 | 32 ± 16 | 32 ± 16 | 31 ± 16 | 32 ± 14 | 32 ± 14 | 33 ± 14 | 0.742 |
| % change | (BL) −24.4 ↓ *** | −5.9 | −5.5 | −8.8 | −5.4 | −5.9 | −2.9 | quadratic | |
| SmO2 re-saturation | 48 ± 20 | 38 ± 17 | 36 ± 16 | 37 ± 16 | 37 ± 16 | 38 ± 16 | 40 ± 17 | 41 ± 17 | 0.730 |
| % change | (BL) −20.8 ↓ *** | −5.3 | 2.8 | 0 | −0.0 | 5.3 | 7.9 | quadratic | |
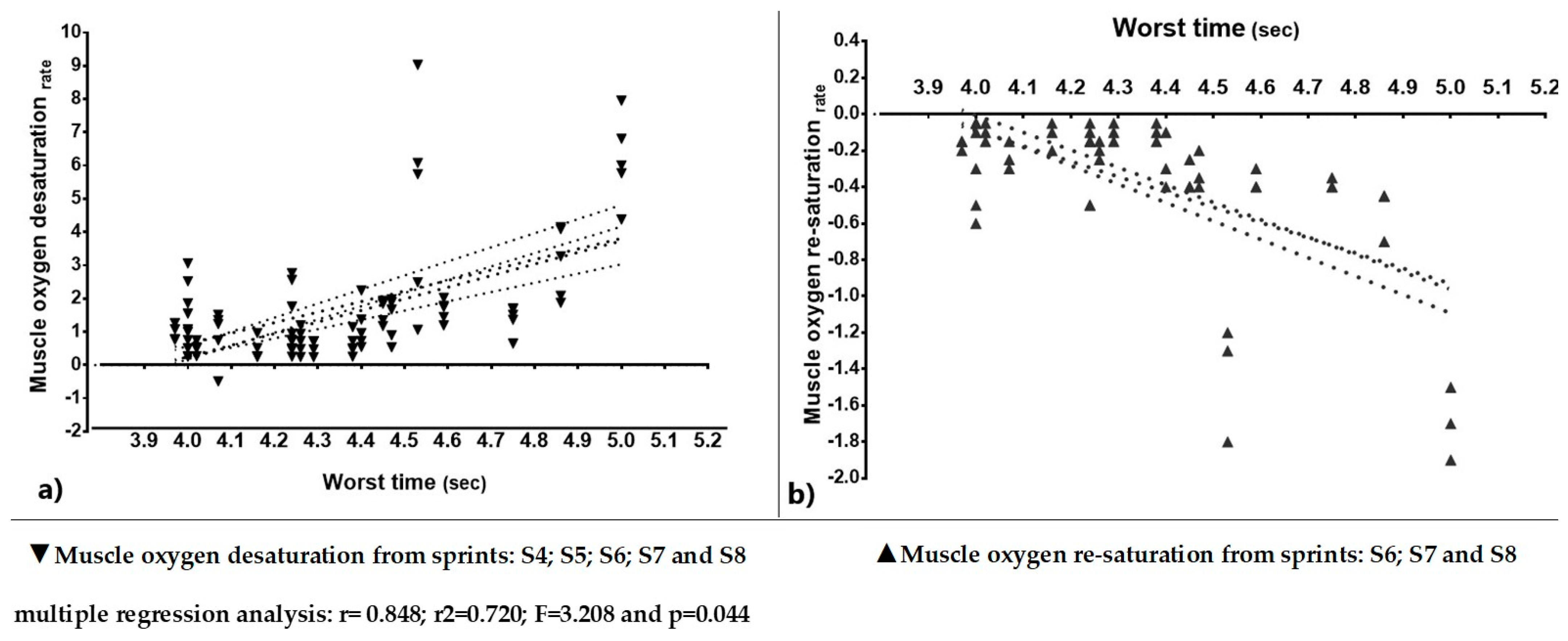
| Desaturation rate | −0.22 ± 1.88 | 0.93 ± 1.21 | 1.13 ± 0.87 | 1.23 ± 1.19 | 1.47 ± 1,40 | 1.58 ± 1.73 | 2.07 ± 2.41 | 1.76 ± 1.74 | 0.617 |
| % change | (BL) −523 ↑ *** | 21.5 | 32.3 ↑ * | 58.1 ↑ ** | 69.9 ↑ ** | 122.7 ↑ *** | 89.2 ↑ ** | quadratic | |
| Re-saturation rate | 0.04 ± 0.37 | −0.18 ± 0.34 | −0.23 ± 0.18 | −0.25 ± 0.26 | −0.30 ± 0.29 | −0.34 ± 0.39 | −0.43 ± 0.52 | −0.38 ± 0.42 | 0.588 |
| % change | −550 ↓↔ * | 27.8 | 38.9 | 66.7 | 88.9 ↓↔ * | 138.9 ↓↔ * | 111.8 ↓↔ * | quadratic | |
| ∇%SmO2 | 0.20 ± 17.1 | 14.3 ± 19.5 | 19.2 ± 20.5 | 24.7 ± 48.5 | 38.4 ± 85.7 | 26.1 ± 38.2 | 40.4 ± 84.4 | 28.1 ± 43.7 | 0.305 |
| % change | (BL) 6709 ↑ ** | 36.4 | 72.7 ↑ * | 168.5 ↑ ** | 82.5 ↑ ** | 182.5 ↑ *** | 96.5 ↑ *** | quadratic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasquez-Bonilla, A.A.; Camacho-Cardeñosa, A.; Timón, R.; Martínez-Guardado, I.; Camacho-Cardeñosa, M.; Olcina, G. Muscle Oxygen Desaturation and Re-Saturation Capacity Limits in Repeated Sprint Ability Performance in Women Soccer Players: A New Physiological Interpretation. Int. J. Environ. Res. Public Health 2021, 18, 3484. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18073484
Vasquez-Bonilla AA, Camacho-Cardeñosa A, Timón R, Martínez-Guardado I, Camacho-Cardeñosa M, Olcina G. Muscle Oxygen Desaturation and Re-Saturation Capacity Limits in Repeated Sprint Ability Performance in Women Soccer Players: A New Physiological Interpretation. International Journal of Environmental Research and Public Health. 2021; 18(7):3484. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18073484
Chicago/Turabian StyleVasquez-Bonilla, Aldo A., Alba Camacho-Cardeñosa, Rafael Timón, Ismael Martínez-Guardado, Marta Camacho-Cardeñosa, and Guillermo Olcina. 2021. "Muscle Oxygen Desaturation and Re-Saturation Capacity Limits in Repeated Sprint Ability Performance in Women Soccer Players: A New Physiological Interpretation" International Journal of Environmental Research and Public Health 18, no. 7: 3484. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18073484