
Impact of Olive Oil Supplement Intake on Dendritic Cell Maturation after Strenuous Physical Exercise: A Preliminary Study

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. Supplements
2.4. Protocol
2.5. Dietary Assessment
2.6. Analytical Procedures
2.6.1. Physical Test and Metabolic Analysis
2.6.2. Antibodies and Reagents
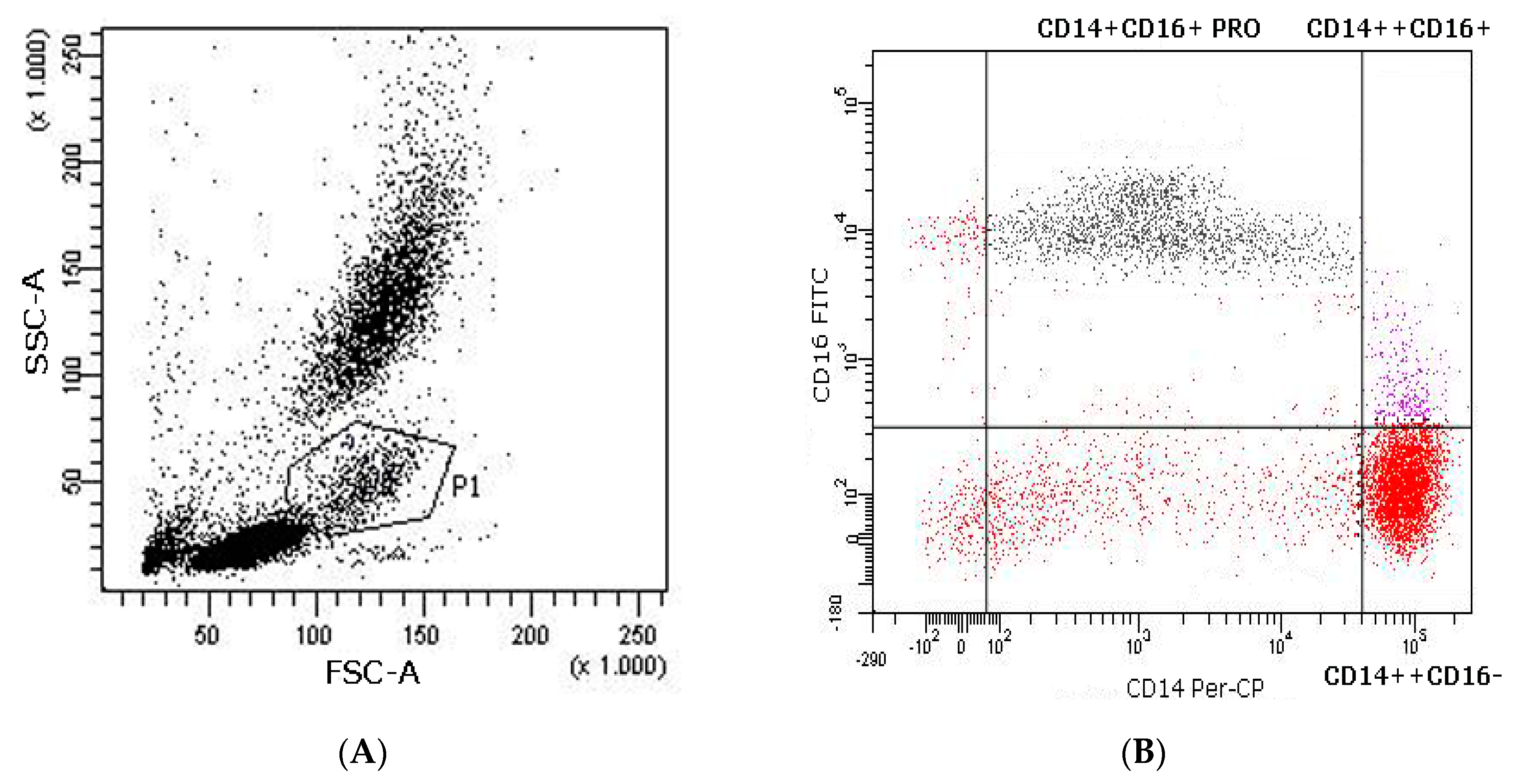
2.6.3. Flow Cytometry Analysis of DC Phenotype, Co-Stimulatory Molecules, and Subsets
2.6.4. Flow Cytometry Analysis of Monocyte Subpopulations in Peripheral Blood
2.6.5. Flow Cytometry Cell Cytokine Analysis
2.7. Statistical Analysis
3. Results
3.1. Participants
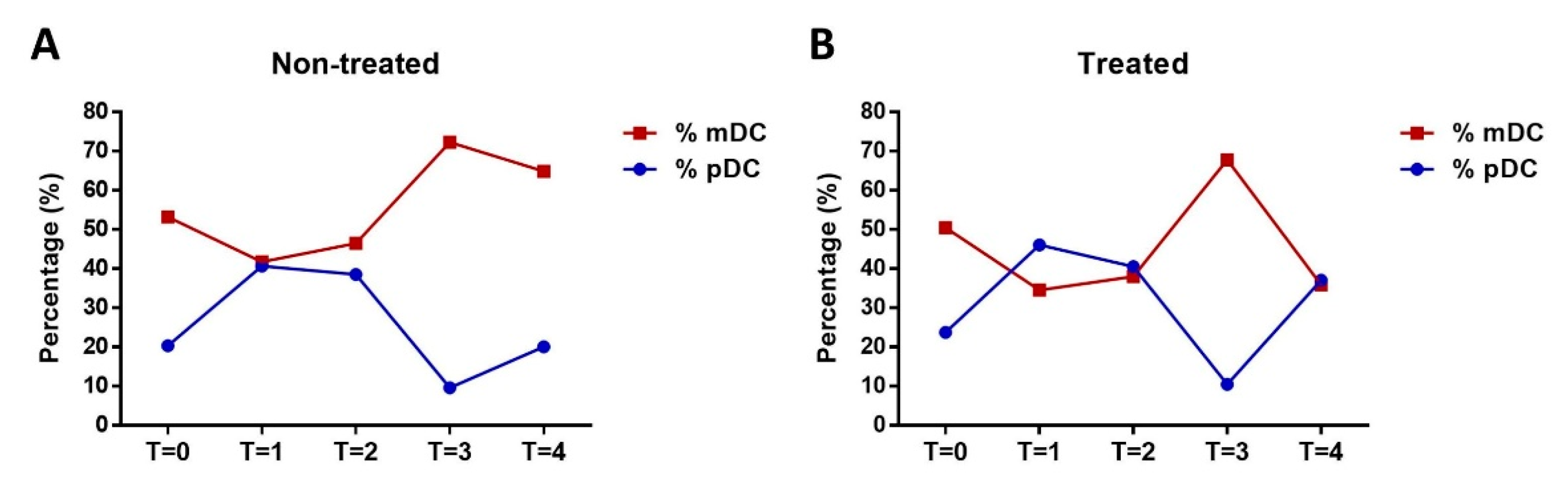
3.2. Analysis of pDC and mDC Populations and Monocytes after Exercise
3.3. Impact of Olive Oil Supplement on DC Maturation
3.4. Metabolic Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De la Torre-Robles, A.; Rivas, A.; Lorenzo-Tovar, M.L.; Monteagudo, C.; Mariscal-Arcas, M.; Olea-Serrano, F. Estimation of the Intake of Phenol Compounds from Virgin Olive Oil of a Population from Southern Spain. Food Addit. Contam. Part A 2014, 31, 1460–1469. [Google Scholar] [CrossRef]
- Mitjavila, M.T.; Fandos, M.; Salas-Salvadó, J.; Covas, M.-I.; Borrego, S.; Estruch, R.; Lamuela-Raventós, R.; Corella, D.; Martínez-Gonzalez, M.Á.; Sánchez, J.M.; et al. The Mediterranean Diet Improves the Systemic Lipid and DNA Oxidative Damage in Metabolic Syndrome Individuals. A Randomized, Controlled, Trial. Clin. Nutr. 2013, 32, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet. N. Engl. J. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef] [Green Version]
- Zamora-Ros, R.; Serafini, M.; Estruch, R.; Lamuela-Raventós, R.M.; Martínez-González, M.A.; Salas-Salvadó, J.; Fiol, M.; Lapetra, J.; Arós, F.; Covas, M.I.; et al. Mediterranean Diet and Non Enzymatic Antioxidant Capacity in the PREDIMED Study: Evidence for a Mechanism of Antioxidant Tuning. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 1167–1174. [Google Scholar] [CrossRef]
- Covas, M.-I.; Nyyssönen, K.; Poulsen, H.E.; Kaikkonen, J.; Zunft, H.-J.F.; Kiesewetter, H.; Gaddi, A.; de la Torre, R.; Mursu, J.; Bäumler, H.; et al. The Effect of Polyphenols in Olive Oil on Heart Disease Risk Factors: A Randomized Trial. Ann. Intern. Med. 2006, 145, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Ginter, E.; Simko, V. Plant Polyphenols in Prevention of Heart Disease. Bratisl. Lek. Listy 2012, 113, 476–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menendez, J. Crude Phenolic Extracts from Extra Virgin Olive Oil Circumvent de Novo Breast Cancer Resistance to HER1/HER2-Targeting Drugs by Inducing GADD45-Sensed Cellular Stress, G2/M Arrest and Hyperacetylation of Histone H3. Int. J. Oncol. 2011. [Google Scholar] [CrossRef] [Green Version]
- Menendez, J.A.; Vazquez-Martin, A.; Garcia-Villalba, R.; Carrasco-Pancorbo, A.; Oliveras-Ferraros, C.; Fernandez-Gutierrez, A.; Segura-Carretero, A. tabAnti-HER2 (erbB-2) Oncogene Effects of Phenolic Compounds Directly Isolated from Commercial Extra-Virgin Olive Oil (EVOO). BMC Cancer 2008, 8, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, S.K.; Tucker, G.A.; Brameld, J.M. Effects of Dietary Polyphenols on Gene Expression in Human Vascular Endothelial Cells. Proc. Nutr. Soc. 2018. [Google Scholar] [CrossRef]
- Arts, I.C.W.; Hollman, P.C.H. Polyphenols and Disease Risk in Epidemiologic Studies. Am. J. Clin. Nutr. 2005, 81 (Suppl. 1), 317S–325S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic Biochemical Mechanisms behind the Health Benefits of Polyphenols. Mol. Asp. Med. 2010, 31, 435–445. [Google Scholar] [CrossRef]
- Tangney, C.C.; Rasmussen, H.E. Polyphenols, Inflammation, and Cardiovascular Disease. Curr. Atheroscler. Rep. 2013, 15, 324. [Google Scholar] [CrossRef]
- Hoffman-Goetz, L.; Pedersen, B.K. Exercise and the Immune System: A Model of the Stress Response? Immunol. Today 1994, 15, 382–387. [Google Scholar] [CrossRef]
- Barros, E.S.; Nascimento, D.C.; Prestes, J.; Nóbrega, O.T.; Córdova, C.; Sousa, F.; Boullosa, D.A. Acute and Chronic Effects of Endurance Running on Inflammatory Markers: A Systematic Review. Front. Physiol. 2017, 8, 779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcante, P.A.M.; Gregnani, M.F.; Henrique, J.S.; Ornellas, F.H.; Araújo, R.C. Aerobic But Not Resistance Exercise Can Induce Inflammatory Pathways via Toll-Like 2 and 4: A Systematic Review. Sports Med. Open 2017, 3, 42. [Google Scholar] [CrossRef]
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of Dendritic Cells. Annu. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef]
- Liu, Y.J. Dendritic Cell Subsets and Lineages, and Their Functions in Innate and Adaptive Immunity. Cell 2001, 106, 259–262. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, F.; Johnson, P.; Williams, A.P. Enumeration and Phenotypic Assessment of Human Plasmacytoid and Myeloid Dendritic Cells in Whole Blood. Cytom. A 2010, 77, 328–337. [Google Scholar] [CrossRef]
- Ho, C.S.K. Surgical and Physical Stress Increases Circulating Blood Dendritic Cell Counts Independently of Monocyte Counts. Blood 2001, 98, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Nickel, T.; Emslander, I.; Sisic, Z.; David, R.; Schmaderer, C.; Marx, N.; Schmidt-Trucksäss, A.; Hoster, E.; Halle, M.; Weis, M.; et al. Modulation of Dendritic Cells and Toll-like Receptors by Marathon Running. Eur. J. Appl. Physiol. 2012, 112, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Suchánek, O.; Podrazil, M.; Fischerová, B.; Bočínská, H.; Budínský, V.; Stejskal, D.; Spíšek, R.; Bartůňková, J.; Kolář, P. Intensive Physical Activity Increases Peripheral Blood Dendritic Cells. Cell. Immunol. 2010, 266, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Deckx, N.; Wens, I.; Nuyts, A.H.; Hens, N.; De Winter, B.Y.; Koppen, G.; Goossens, H.; Van Damme, P.; Berneman, Z.N.; Eijnde, B.O.; et al. 12 Weeks of Combined Endurance and Resistance Training Reduces Innate Markers of Inflammation in a Randomized Controlled Clinical Trial in Patients with Multiple Sclerosis. Mediat. Inflamm. 2016, 2016, 6789276. [Google Scholar] [CrossRef] [PubMed]
- Deckx, N.; Wens, I.; Nuyts, A.H.; Lee, W.-P.; Hens, N.; Koppen, G.; Goossens, H.; Van Damme, P.; Berneman, Z.N.; Eijnde, B.O.; et al. Rapid Exercise-Induced Mobilization of Dendritic Cells Is Potentially Mediated by a Flt3L- and MMP-9-Dependent Process in Multiple Sclerosis. Mediat. Inflamm. 2015, 2015, 158956. [Google Scholar] [CrossRef]
- LaVoy, E.C.P.; Bollard, C.M.; Hanley, P.J.; O’Connor, D.P.; Lowder, T.W.; Bosch, J.A.; Simpson, R.J. A Single Bout of Dynamic Exercise by Healthy Adults Enhances the Generation of Monocyte-Derived-Dendritic Cells. Cell. Immunol. 2015, 295, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Morgado, J.M.; Rama, L.; Silva, I.; de Jesus Inácio, M.; Henriques, A.; Laranjeira, P.; Pedreiro, S.; Rosado, F.; Alves, F.; Gleeson, M.; et al. Cytokine Production by Monocytes, Neutrophils, and Dendritic Cells Is Hampered by Long-Term Intensive Training in Elite Swimmers. Eur. J. Appl. Physiol. 2012, 112, 471–482. [Google Scholar] [CrossRef]
- Jeukendrup, A.E.; Saris, W.H.; Wagenmakers, A.J. Fat metabolism during exercise: A review. Part I: Fatty acid mobilization and muscle metabolism. Int. J. Sports Med. 1998, 19, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.J.C.; Greenha, P.L.; Constantin-Teodosiu, D.; Saris, W.H.M.; Wagenmakers, A.J.M. The effects increasing exercise intensity on muscle fuel utilisation in humans. J. Physiol. 2001, 536, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.E.; Saris, W.H.; Wagenmakers, A.J. Fat metabolism during exercise: A review—Part II: Regulation metabolism and the effects training. Int. J. Sports Med. 1998, 19, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Weinbrenner, T.; Fitó, M.; Farré Albaladejo, M.; Saez, G.T.; Rijken, P.; Tormos, C.; Coolen, S.; De La Torre, R.; Covas, M.I. Bioavailability of Phenolic Compounds from Olive Oil and Oxidative/antioxidant Status at Postprandial State in Healthy Humans. Drugs Exp. Clin. Res. 2004, 30, 207–212. [Google Scholar] [PubMed]
- Lloberas, N.; Rama, I.; Llaudó, I.; Torras, J.; Cerezo, G.; Cassis, L.; Franquesa, M.; Merino, A.; Benitez-Ribas, D.; Cruzado, J.M.; et al. Dendritic Cells Phenotype Fitting under Hypoxia or Lipopolysaccharide; Adenosine 5′-Triphosphate-Binding Cassette Transporters Far beyond an Efflux Pump. Clin. Exp. Immunol. 2013, 172, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Trivax, J.E.; Franklin, B.A.; Goldstein, J.A.; Chinnaiyan, K.M.; Gallagher, M.J.; de Jong, A.T.; Colar, J.M.; Haines, D.E.; McCullough, P.A. Acute Cardiac Effects of Marathon Running. J. Appl. Physiol. 2010, 108, 1148–1153. [Google Scholar] [CrossRef] [Green Version]
- Kasapis, C.; Thompson, P.D. The Effects of Physical Activity on Serum C-Reactive Protein and Inflammatory Markers: A Systematic Review. J. Am. Coll. Cardiol. 2005, 45, 1563–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Nakaji, S.; Yamada, M.; Liu, Q.; Kurakake, S.; Okamura, N.; Kumae, T.; Umeda, T.; Sugawara, K. Impact of a Competitive Marathon Race on Systemic Cytokine and Neutrophil Responses. Med. Sci. Sports Exerc. 2003, 35, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Steensberg, A.; Fischer, C.; Keller, C.; Ostrowski, K.; Schjerling, P. Exercise and Cytokines with Particular Focus on Muscle-Derived IL-6. Exerc. Immunol. Rev. 2001, 7, 18–31. [Google Scholar]
- Lackermair, K.; Scherr, J.; Waidhauser, G.; Methe, H.; Hoster, E.; Nieman, D.C.; Hanley, A.; Clauss, S.; Halle, M.; Nickel, T. Influence of Polyphenol-Rich Diet on Exercise-Induced Immunomodulation in Male Endurance Athletes. Appl. Physiol. Nutr. Metab. 2017, 42, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Bowtell, J.; Kelly, V. Fruit-Derived Polyphenol Supplementation for Athlete Recovery and Performance. Sports Med. 2019, 49 (Suppl. 1), S3–S23. [Google Scholar] [CrossRef] [Green Version]
- Myburgh, K.H. Polyphenol Supplementation: Benefits for Exercise Performance or Oxidative Stress? Sports Med. 2014, 44 (Suppl. 1), S57–S70. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Supplement | Placebo Supplement | |
|---|---|---|
| Ingredients | 100 mL commercial orange juice 8 g modified starch 25 mL extra-virgin olive oil | 100 mL commercial orange juice 8 g modified starch |
| Energy (Kcal) | 277 | 52.8 |
| Fat (g) | 25 | 0.1 |
| Carbohydrates (g) | 12.6 | 12.6 |
| Subject | #1 | #2 | #3 |
|---|---|---|---|
| Age (years) | 51 | 34 | 39 |
| Weight (kg) | 94 | 65 | 71 |
| Height (cm) | 181 | 165 | 178 |
| BMI (kg·m−2) | 28.75 | 23.90 | 22.39 |
| Subjects | Non-Treated | Treated | ||||
|---|---|---|---|---|---|---|
| T2 | T3 | T4 | T2 | T3 | T4 | |
| #1 | 56.4 | 57.8 | 56 | 62.6 | 77.9 | 37.1 |
| CV1 | 1.29 | 2.63 | ||||
| #2 | 46.5 | 72.3 | 64.9 | 38 | 67.8 | 35.9 |
| CV2 | 0.29 | 1.1 | ||||
| #3 | 24.5 | 80.5 | 48.5 | 61.6 | 65.7 | 48.7 |
| CV3 | 0.57 | 4.15 | ||||
| Treated | Non-Treated | |||||
|---|---|---|---|---|---|---|
| Subject | #1 | #2 | #3 | #1 | #2 | #3 |
| MAXIMAL TEST | ||||||
| VE (L·min−1) | 154.1 | 78.8 | 152.4 | 146.5 | 93.5 | 132.9 |
| VO2/Kg (mL·kg·min−1) | 39.3 | 39.6 | 59 | 39.4 | 47.6 | 52.5 |
| QR | 1.127 | 1.064 | 1.13 | 1.121 | 1.016 | 1.087 |
| VCO2 (L·min−1) | 4.17 | 2.74 | 4.73 | 4.15 | 3.14 | 4.05 |
| FC (lat·min−1) | 159 | 177 | 173 | 152 | 183 | 171 |
| Metabolic rate (kcal·day−1) | 27,679 | 19,021 | 31,356 | 27,690 | 22,600 | 27,657 |
| SUBMAXIMAL TEST | ||||||
| 1M-45 | 4947 | 4924 | 5218 | 5072 | 4663 | 5058 |
| 1M-45-O2 | 275.6 | 268.7 | 392.4 | 279.5 | 294.3 | 360.3 |
| 1M-45-QR | 0.977 | 0.875 | 0.907 | 0.968 | 0.924 | 0.89 |
| %VO2 max | 77.1 | 74.2 | 77.1 | 79.9 | ||
| 64.01M-45-kcal | 641.3 | 429.8 | 726.8 | 666 | 445.5 | 645.8 |
| Kcal·metres | 0.130 | 0.087 | 0.139 | 0.131 | 0.096 | 0.128 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esquius, L.; Javierre, C.; Llaudó, I.; Rama, I.; Oviedo, G.R.; Massip-Salcedo, M.; Aguilar-Martínez, A.; Niño, O.; Lloberas, N. Impact of Olive Oil Supplement Intake on Dendritic Cell Maturation after Strenuous Physical Exercise: A Preliminary Study. Int. J. Environ. Res. Public Health 2021, 18, 4128. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18084128
Esquius L, Javierre C, Llaudó I, Rama I, Oviedo GR, Massip-Salcedo M, Aguilar-Martínez A, Niño O, Lloberas N. Impact of Olive Oil Supplement Intake on Dendritic Cell Maturation after Strenuous Physical Exercise: A Preliminary Study. International Journal of Environmental Research and Public Health. 2021; 18(8):4128. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18084128
Chicago/Turabian StyleEsquius, Laura, Casimiro Javierre, Inés Llaudó, Inés Rama, Guillermo R. Oviedo, Marta Massip-Salcedo, Alicia Aguilar-Martínez, Oscar Niño, and Núria Lloberas. 2021. "Impact of Olive Oil Supplement Intake on Dendritic Cell Maturation after Strenuous Physical Exercise: A Preliminary Study" International Journal of Environmental Research and Public Health 18, no. 8: 4128. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18084128