
Immune Response of Elite Enduro Racers to Laboratory and Racing Environments: The Influence of Training Impulse and Vibration

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
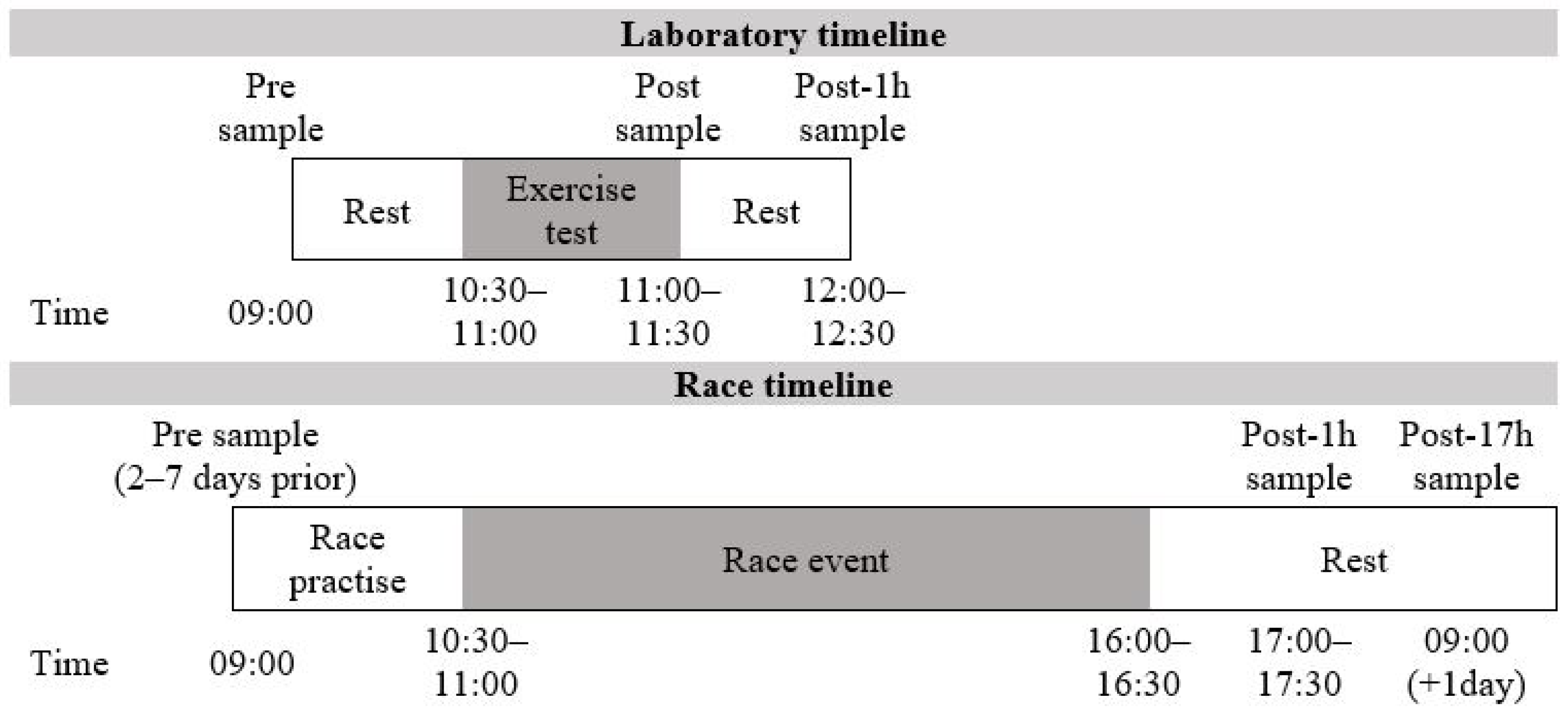
2.2. Laboratory Exercise Protocol
2.3. Race Event Protocol
2.4. Heart Rate and Location Data
2.5. Training Load
2.6. Seat Post Vibration Measurement
2.7. Blood Sampling
2.8. Blood Sample Analysis
2.9. Lymphocyte Phenotyping
2.10. Flow Cytometry and Gating
2.11. Data Analysis
3. Results
3.1. Overview
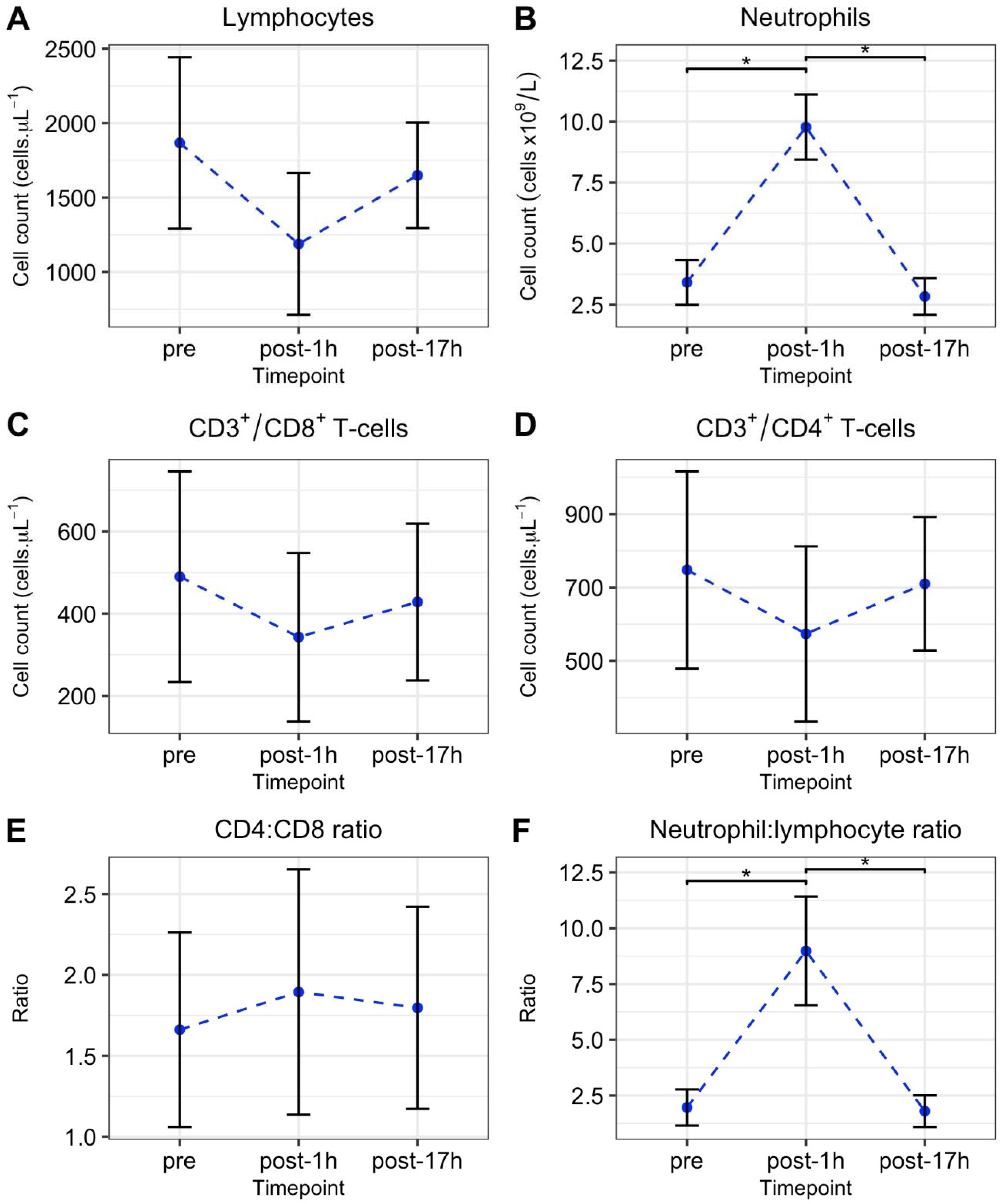
3.2. Laboratory Tests
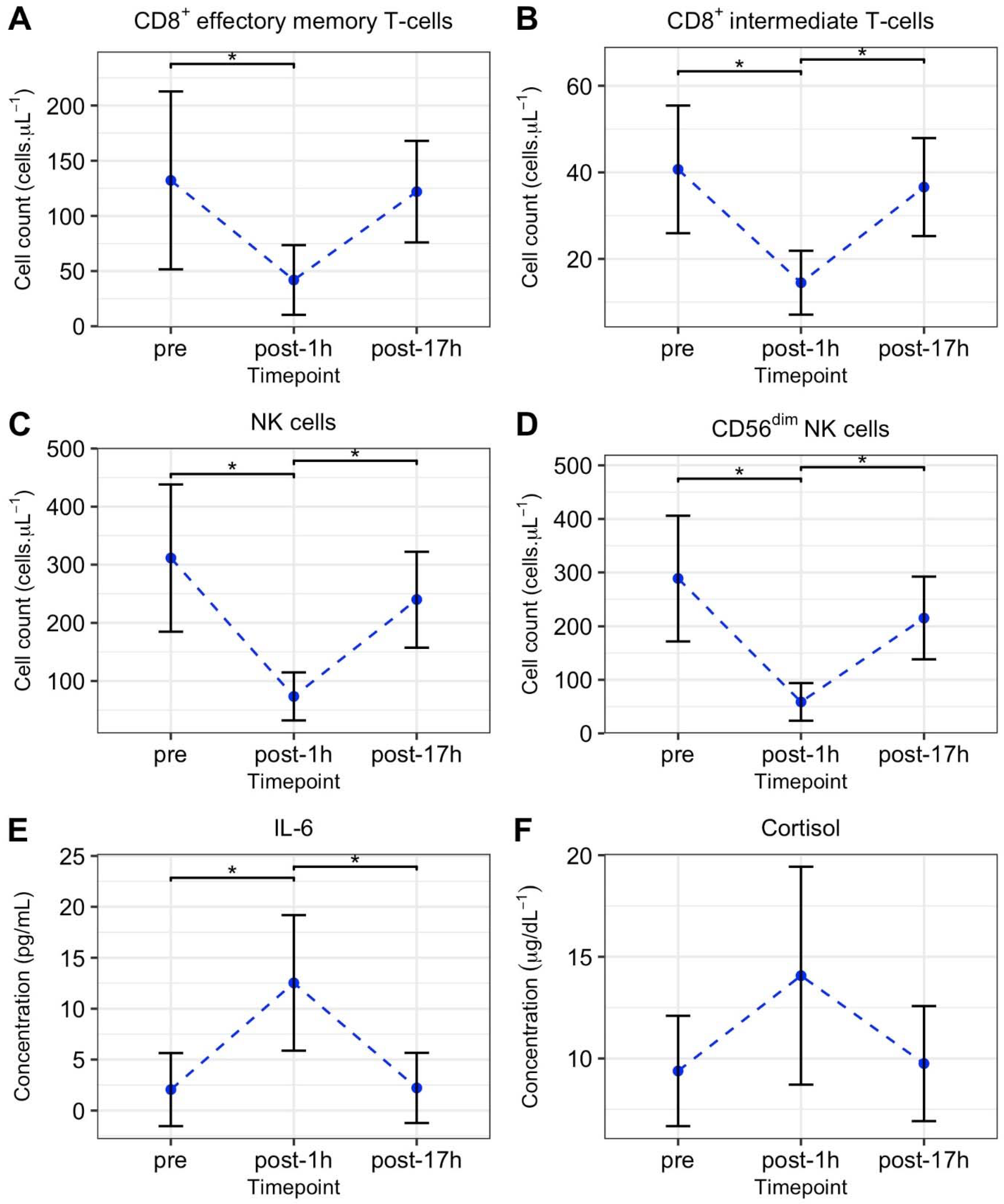
3.3. International Race Event
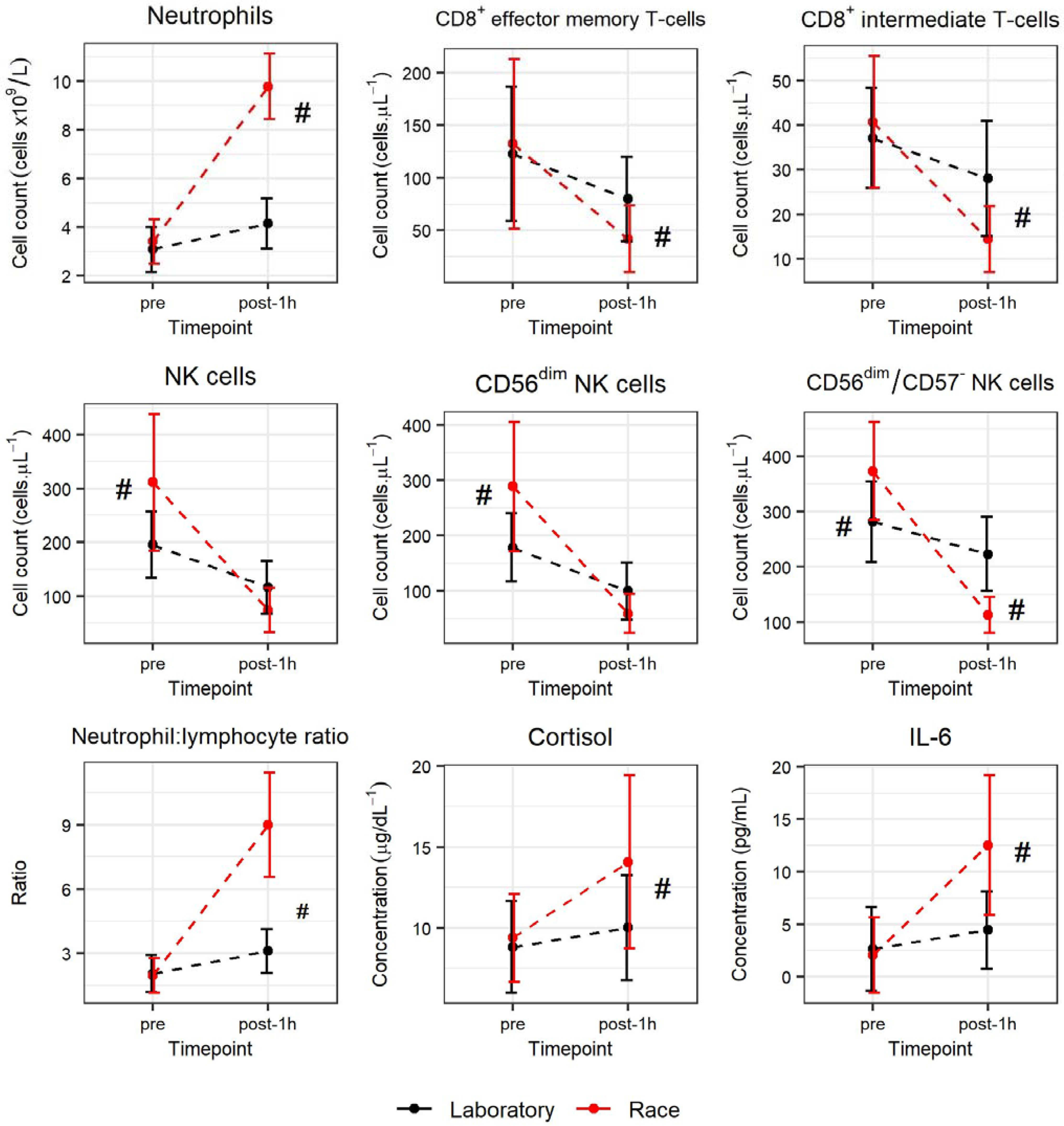
3.4. Comparison of Laboratory Test and International Race Event
3.5. Vibration, Training Impulse, and Redistribution of Lymphocyte Subpopulations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mårtensson, S.; Nordebo, K.; Malm, C. High Training Volumes are Associated with a Low Number of Self-Reported Sick Days in Elite Endurance Athletes. J. Sports Sci. Med. 2014, 13, 929–933. [Google Scholar]
- Raysmith, B.P.; Drew, M.K. Performance success or failure is influenced by weeks lost to injury and illness in elite Australian Track and Field athletes: A 5-year prospective study. J. Sci. Med. Sport 2016, 19, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Hellard, P.; Avalos, M.; Guimaraes, F.; Toussaint, J.-F.; Pyne, D.B. Training-Related Risk of Common Illnesses in Elite Swimmers over a 4-yr Period. Med. Sci. Sports Exerc. 2015, 47, 698–707. [Google Scholar] [CrossRef]
- Svendsen, I.S.; Killer, S.C.; Carter, J.M.; Randell, R.K.; Jeukendrup, A.E.; Gleeson, M. Impact of intensified training and carbohydrate supplementation on immunity and markers of overreaching in highly trained cyclists. Eur. J. Appl. Physiol. 2016, 116, 867–877. [Google Scholar] [CrossRef] [Green Version]
- Ingram, L.A.; Simpson, R.J.; Malone, E.; Florida-James, G.D. Sleep disruption and its effect on lymphocyte redeployment following an acute bout of exercise. Brain Behav. Immun. 2015, 47, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.P.; Turner, J.E. Debunking the Myth of Exercise-Induced Immune Suppression: Redefining the Impact of Exercise on Immunological Health Across the Lifespan. Front. Immunol. 2018, 9, 648. [Google Scholar] [CrossRef]
- Simpson, R.J. Aging, Persistent Viral Infections, and Immunosenescence can exercise ‘make space’? Exerc. Sport Sci. Rev. 2011, 39, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Tasdogan, A.; Ubellacker, J.M.; Zhang, J.; Nosyreva, E.D.; Du, L.; Murphy, M.M.; Hu, S.; Yi, Y.; Kara, N.; et al. A mechanosensitive peri-arteriolar niche for osteogenesis and lymphopoiesis. Nat. Cell Biol. 2021, 591, 438–444. [Google Scholar] [CrossRef]
- Simpson, R.J.; Cosgrove, C.; Ingram, L.A.; Florida-James, G.D.; Whyte, G.P.; Pircher, H.; Guy, K. Senescent T-lymphocytes are mobilised into the peripheral blood compartment in young and older humans after exhaustive exercise. Brain Behav. Immun. 2008, 22, 544–551. [Google Scholar] [CrossRef]
- Campbell, J.P.; Riddell, N.E.; Burns, V.E.; Turner, M.; Van Zanten, J.J.V.; Drayson, M.T.; Bosch, J.A. Acute exercise mobilises CD8+ T lymphocytes exhibiting an effector-memory phenotype. Brain Behav. Immun. 2009, 23, 767–775. [Google Scholar] [CrossRef]
- Peake, J.M.; Neubauer, O.; Walsh, N.P.; Simpson, R.J. Recovery of the immune system after exercise. J. Appl. Physiol. 2017, 122, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Graff, R.M.; Kunz, H.E.; Agha, N.H.; Baker, F.L.; Laughlin, M.; Bigley, A.B.; Markofski, M.M.; LaVoy, E.C.; Katsanis, E.; Bond, R.A.; et al. β2-adrenergic receptor signaling mediates the preferential mobilization of differentiated subsets of CD8+ T-cells, NK-cells and non-classical monocytes in response to acute exercise in humans. Brain Behav. Immun. 2018, 74, 143–153. [Google Scholar] [CrossRef]
- Schaller, K.; Mechau, D.; Scharmann, H.G.; Weiss, M.; Baum, M.; Liesen, H. Increased training load and the be-ta-adrenergic-receptor system on human lymphocytes. J. Appl. Physiol. 1999, 87, 317–324. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/10409590 (accessed on 21 February 2021). [CrossRef] [Green Version]
- Hill, E.E.; Zack, E.; Battaglini, C.; Viru, M.; Viru, A.; Hackney, A.C. Exercise and circulating Cortisol levels: The intensity threshold effect. J. Endocrinol. Investig. 2008, 31, 587–591. [Google Scholar] [CrossRef]
- Reihmane, D.; Jurka, A.; Tretjakovs, P.; Dela, F. Increase in IL-6, TNF-α, and MMP-9, but not sICAM-1, concentrations depends on exercise duration. Eur. J. Appl. Physiol. 2013, 113, 851–858. [Google Scholar] [CrossRef]
- Atanackovic, D.; Schnee, B.; Schuch, G.; Faltz, C.; Schulze, J.; Weber, C.S.; Schafhausena, P.; Bartelsa, K.; Bokemeyera, C.; Brunner-Weinzierl, M.C. Acute psychological stress alerts the adaptive immune response: Stress-induced mobilization of effec-tor T cells. J. Neuroimmunol. 2006, 176, 141–152. [Google Scholar] [CrossRef] [PubMed]
- McFarlin, B.K.; Mitchell, J.B. Exercise in hot and cold environments: Differential effects on leukocyte number and NK cell activity. Aviat. Space Environ. Med. 2003, 74, 1231–1236. [Google Scholar]
- Lippi, G.; Banfi, G.; Montagnana, M.; Salvagno, G.L.; Schena, F.; Guidi, G.C. Acute variation of leucocytes counts following a half-marathon run. Int. J. Lab. Hematol. 2008, 32, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, P.; Baumann, F.T.; Bloch, W.; Zopf, E.M.; Schulz, S.; Latsch, J.; Schollmayer, F.; Shimabukuro-Vornhagen, A.; Von Bergwelt-Baildon, M.; Schenk, A. Impact of a half marathon on cellular immune system, pro-inflammatory cytokine levels, and recovery behavior of breast cancer patients in the aftercare compared to healthy controls. Eur. J. Haematol. 2015, 96, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Haq, A.; Al-Hussein, K.; Lee, J.; Al-Sedairy, S. Changes in peripheral blood lymphocyte subsets associated with marathon running. Med. Sci. Sport. Exerc. 1993, 25, 186–190. [Google Scholar] [CrossRef]
- Castell, L.M.; Poortmans, J.R.; Leclercq, R.; Brasseur, M.; Duchateau, J.; Newsholme, E.A. Some aspects of the acute phase response after a marathon race, and the effects of glutamine supplementation. Eur. J. Appl. Physiol. Occup. Physiol. 1997, 75, 47–53. [Google Scholar] [CrossRef]
- Suzuki, K.; Nakaji, S.; Yamada, M.; Liu, Q.; Kurakake, S.; Okamura, N.; Kumae, T.; Umeda, T.; Sugawara, K. Impact of a Competitive Marathon Race on Systemic Cytokine and Neutrophil Responses. Med. Sci. Sports Exerc. 2003, 35, 348–355. [Google Scholar] [CrossRef]
- Rooney, B.V.; Bigley, A.B.; LaVoy, E.C.; Laughlin, M.; Pedlar, C.; Simpson, R.J. Lymphocytes and monocytes egress pe-ripheral blood within minutes after cessation of steady state exercise: A detailed temporal analysis of leukocyte extravasa-tion. Physiol. Behav. 2018, 194, 260–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratz, A.; Lewandrowski, K.B.; Siegel, A.J.; Chun, K.Y.; Flood, J.G.; Van Cott, E.M.; Lee-Lewandrowski, E. Effect of Marathon Running on Hematologic and Biochemical Laboratory Parameters, Including Cardiac Markers. Am. J. Clin. Pathol. 2002, 118, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, L.A.; Ingram, L.A.; Cunningham, J.; Malone, E.; Florida-James, G. Physiological characteristics and perfor-mance in elite vs. non-elite enduro mountain biking. J. Sci. Cycl. 2017, 6, 13–21. [Google Scholar] [CrossRef]
- Kirkwood, L.A.; Taylor, M.D.; Ingram, L.A.; Malone, E.; Florida-James, G.D. Elite mountain bike enduro competition: A study of rider hand-arm vibration exposure. J. Sci. Cycl. 2019, 8, 18–25. [Google Scholar] [CrossRef]
- Noguchi, R.; Ando, H. Immune responses (CD4 and CD8) to acute vibration stress. Kurume Med. J. 2002, 49, 87–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeusen, R.; Duclos, M.; Foster, C.; Fry, A.; Gleeson, M.; Nieman, D.; Raglin, J.; Rietjens, G.; Steinacker, J.; Urhausen, A. Prevention, diagnosis, and treatment of the overtraining syndrome: Joint consensus statement of the Eu-ropean College of Sport Science and the American College of Sports Medicine. Med. Sci. Sports Exerc. 2013, 45, 186–205. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, G.; Reilly, T. Circadian Variation in Sports Performance. Sports Med. 1996, 21, 292–312. [Google Scholar] [CrossRef]
- Bonaventura, J.M.; Sharpe, K.; Knight, E.; Fuller, K.L.; Tanner, R.K.; Gore, C.J. Reliability and Accuracy of Six Hand-Held Blood Lactate Analysers. J. Sports Sci. Med. 2014, 14, 203–214. [Google Scholar]
- Mulas, A.L.; Hoyos, J.; Hernández, A.S.; Earnest, C.; Chicharro, J.L. Tour de France versus Vuelta a Espana: Which is harder? Med. Sci. Sports Exerc. 2003, 35, 872–878. [Google Scholar] [CrossRef]
- Sanders, D.; Abt, G.; Hesselink, M.K.; Myers, T.; Akubat, I. Methods of Monitoring Training Load and Their Relationships to Changes in Fitness and Performance in Competitive Road Cyclists. Int. J. Sports Physiol. Perform. 2017, 12, 668–675. [Google Scholar] [CrossRef] [Green Version]
- ISO. ISO 2631-5: Mechanical vibration and shock-evalutation of human exposure to whole-body vibration. In Part 5: Method for Evalutation of Vibration Containing Multiple Shocks; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- Dill, D.B.; Costill, D.L. Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. J. Appl. Physiol. 1974, 37, 247–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, R.J.; Florida-James, G.D.; Whyte, G.P.; Guy, K. The effects of intensive, moderate and downhill treadmill running on human blood lymphocytes expressing the adhe-sion/activation molecules CD54 (ICAM-1), CD18 (beta2 integrin) and CD53. Eur. J. Appl. Physiol. 2006, 97, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Krüger, K.; Mooren, F.C. T cell homing and exercise. Exerc. Immunol. Rev. 2007, 13, 37–54. [Google Scholar] [PubMed]
- Appay, V.; Rowland-Jones, S.L. Lessons from the study of T-cell differentiation in persistent human virus infection. Semin. Immunol. 2004, 16, 205–212. [Google Scholar] [CrossRef]
- Cooper, M.A.; Fehniger, T.A.; Turner, S.C.; Chen, K.S.; Ghaheri, B.A.; Ghayur, T.; Carson, W.E.; Caligiuri, M.A. Human natural killer cells: A unique innate immunoregulatory role for the CD56bright subset. Immunobiology 2001, 97, 3146–3151. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.M.; White, M.J.; Goodier, M.R.; Riley, E.M. Functional Significance of CD57 Expression on Human NK Cells and Relevance to Disease. Front. Immunol. 2013, 4, 422. [Google Scholar] [CrossRef] [Green Version]
- Behrendt, S. lm.beta: Add Standardized Regression Coefficients to lm-Objects. 2014. Available online: https://cran.r-project.org/package=lm.beta (accessed on 8 July 2020).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Shephard, R.J. Adhesion Molecules, Catecholamines and Leucocyte Redistribution During and Following Exercise. Sports Med. 2003, 33, 261–284. [Google Scholar] [CrossRef] [PubMed]
- Krüger, K.; Lechtermann, A.; Fobker, M.; Völker, K.; Mooren, F. Exercise-induced redistribution of T lymphocytes is regulated by adrenergic mechanisms. Brain Behav. Immun. 2008, 22, 324–338. [Google Scholar] [CrossRef]
- Neubauer, O.; Sabapathy, S.; Lazarus, R.; Jowett, J.B.M.; Desbrow, B.; Peake, J.M.; Cameron-Smith, D.; Haseler, L.J.; Wagner, K.-H.; Bulmer, A.C. Transcriptome analysis of neutrophils after endurance exercise reveals novel signaling mechanisms in the immune response to physiological stress. J. Appl. Physiol. 2013, 114, 1677–1688. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.J.; Guy, K.; Whyte, G.P.; Middleton, N.; Black, J.R.; Ross, J.A.; Shave, R.; Florida-James, G.D. Lymphocyte Phenotype Alterations, Pro-Inflammatory Cytokines and Acute Phase Proteins Following Repeated Bouts of Mountainous Hill-Running. Med. Sci. Sports Exerc. 2006, 38, 2726–2727. [Google Scholar] [CrossRef]
- Santos, V.C.; Alves, S.R.; Pithon-Curi, T.C.; Curi, R.; Levada-Pires, A.C.; Cury-Boaventura, M.F. Changes in lymphocyte and neutrophil function induced by a marathon race. Cell Biochem. Funct. 2012, 31, 237–243. [Google Scholar] [CrossRef]
- Van Hall, G.; Steensberg, A.; Sacchetti, M.; Fischer, C.; Keller, C.; Schjerling, P.; Hiscock, N.; Møller, K.; Saltin, B.; Febbraio, M.A.; et al. Interleukin-6 Stimulates Lipolysis and Fat Oxidation in Humans. J. Clin. Endocrinol. Metab. 2003, 88, 3005–3010. [Google Scholar] [CrossRef]
- Kirschbaum, C.; Prussner, J.C.; Stone, A.A.; Federenko, I.; Gaab, J.; Lintz, D.; Schommer, N.; Hellhammer, D.H. Persistent High Cortisol Responses to Repeated Psychological Stress in a Subpopulation of Healthy Men. Psychosom. Med. 1995, 57, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K. Cytokine Response to Exercise and Its Modulation. Antioxidants 2018, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Duc, S.; Puel, F.; Bertucci, W. Vibration exposure on cobbles sectors during ParisRoubaix. Sci. Cycl. Caen 2016, 5, 19–20. [Google Scholar]
- Hurst, H.T.; Swarén, M.; Hébert-Losier, K.; Ericsson, F.; Sinclair, J.; Atkins, S.; Homlberg, H.C. GPS-Based Evaluation of Activity Profiles in Elite Downhill Mountain Biking and the Influence of Course Type. J. Sci. Cycl. 2013, 2, 25. [Google Scholar]
- Samuelson, B.; Jorfeldt, L.; Ahlborg, B. Influence of Vibration on Work Performance during Ergometer Cycling. Upsala J. Med Sci. 1989, 94, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Rønnestad, B.R.; Moen, M.; Gunnerød, S.; Øfsteng, S. Adding vibration to high-intensity intervals increase time at high oxygen uptake in well-trained cyclists. Scand. J. Med. Sci. Sports 2018, 28, 2473–2480. [Google Scholar] [CrossRef] [PubMed]
- Macdermid, P.W.; Fink, P.W.; Stannard, S.R. Transference of 3D accelerations during cross country mountain biking. J. Biomech. 2014, 47, 1829–1837. [Google Scholar] [CrossRef] [PubMed]
- Stupka, N.; Lowther, S.; Chorneyko, K.; Bourgeois, J.M.; Hogben, C.; Tarnopolsky, M.A. Gender differences in muscle inflammation after eccentric exercise. J. Appl. Physiol. 2000, 89, 2325–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subset | Identification | References | |
|---|---|---|---|
| T-cell (CD3+/CD4+ or CD3+/CD8+) | Naive | CD62L+/CD45RA+ | [7,36] |
| Central memory | CD62L+/CD45RA− | ||
| Effector memory | CD62L−/CD45RA− | ||
| EMRA | CD62L−/CD45RA+ | ||
| Early | CD27+/CD28+ | [37] | |
| Intermediate | CD27+/CD28− | ||
| Late | CD27−/CD28− | ||
| NK cell (CD3−/CD56+) | Cytotoxic | CD56dim | [38,39] |
| Regulatory | CD56bright | ||
| Early | CD56+/CD57− | ||
| Late | CD56+/CD57+ | ||
| Variable | Time Point | Effect of Time Point | ||
|---|---|---|---|---|
| Pre | Post | Post-1h | ||
| Lymphocytes (cells·µL−1) | 1581 ± 312 | 4432 ± 855 * | 1388 ± 265 # | F1.44,6.62 = 66.32, p < 0.001, ω2 = 0.86 |
| Neutrophils (cells·µL−1) | 3091 ± 921 | 5155 ± 1582 * | 4164 ± 1035 | F2,18 = 21.27, p < 0.001, ω2 = 0.28 |
| CD8+ T-cells (cells·µL−1) | 429 ± 142 | 1027 ± 329 * | 350 ± 107 # | F1.06,6.39 = 36.97, p < 0.001, ω2 = 0.65 |
| CD4+ T-cells (cells·µL−1) | 686 ± 177 | 1143 ± 219 * | 637 ± 236 # | F2,18 = 32.13, p < 0.001, ω2 = 0.51 |
| NK cells (cells·µL−1) | 196 ± 62 | 1605 ± 804 * | 116 ± 49 # | F1.01, 6.06 = 24.46, p = 0.002 ω2 = 0.67 |
| CD4:CD8 ratio | 1.7 ± 0.6 | 1.2 ± 0.5 | 1.9 ± 0.9 | F1.08,6.51 = 23.44, p = 0.002, ω2 = 0.8 |
| Neutrophil:lymphocyte ratio | 2.1 ± 0.9 | 1.2 ± 0.5 * | 3.1 ± 1.0 # | F2,18 = 31.97, p < 0.001, ω2 = 0.43 |
| Cortisol (µg·dL−1) | 8.8 ± 2.8 | 10.2 ± 2.7 | 10.02 ± 3.2 | F2,18 = 2.14, p = 0.16, ω2 = −0.05 |
| Cell Subset | ΔF(1,3) | ΔR2 | p | βLuTRIMP | βVIB |
|---|---|---|---|---|---|
| Neutrophils | 0.05 | 0.01 | 0.835 | −0.09 | 0.11 |
| Lymphocytes | 7.16 | 0.63 | 0.055 | −0.30 | −0.83 |
| CD8+ T-cells | 1.13 | 0.22 | 0.347 | −0.09 | −0.49 |
| CD4+ T-cells | 11.13 | 0.73 | 0.029 * | −0.18 | −0.89 |
| CD4+ Naïve (CD62L+/CD45RA+) | 34.25 | 0.89 | 0.004 * | −0.25 | −0.99 |
| CD4+ Senescent (CD62L−/CD45RA+) | 7.19 | 0.63 | 0.055 | −0.07 | −0.82 |
| CD4+ early (CD27+/CD28+) | 10.22 | 0.72 | 0.033 * | −0.21 | −0.88 |
| CD3−/CD56+ NK cells | 4.12 | 0.63 | 0.050 * | −0.44 | −0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirkwood, L.; Ingram-Sills, L.; Taylor, M.D.; Malone, E.; Florida-James, G. Immune Response of Elite Enduro Racers to Laboratory and Racing Environments: The Influence of Training Impulse and Vibration. Int. J. Environ. Res. Public Health 2021, 18, 4603. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18094603
Kirkwood L, Ingram-Sills L, Taylor MD, Malone E, Florida-James G. Immune Response of Elite Enduro Racers to Laboratory and Racing Environments: The Influence of Training Impulse and Vibration. International Journal of Environmental Research and Public Health. 2021; 18(9):4603. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18094603
Chicago/Turabian StyleKirkwood, Lewis, Lesley Ingram-Sills, Mark Dunlop Taylor, Eva Malone, and Geraint Florida-James. 2021. "Immune Response of Elite Enduro Racers to Laboratory and Racing Environments: The Influence of Training Impulse and Vibration" International Journal of Environmental Research and Public Health 18, no. 9: 4603. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18094603