
Dynamics of a Bacterial Community in the Anode and Cathode of Microbial Fuel Cells under Sulfadiazine Pressure


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
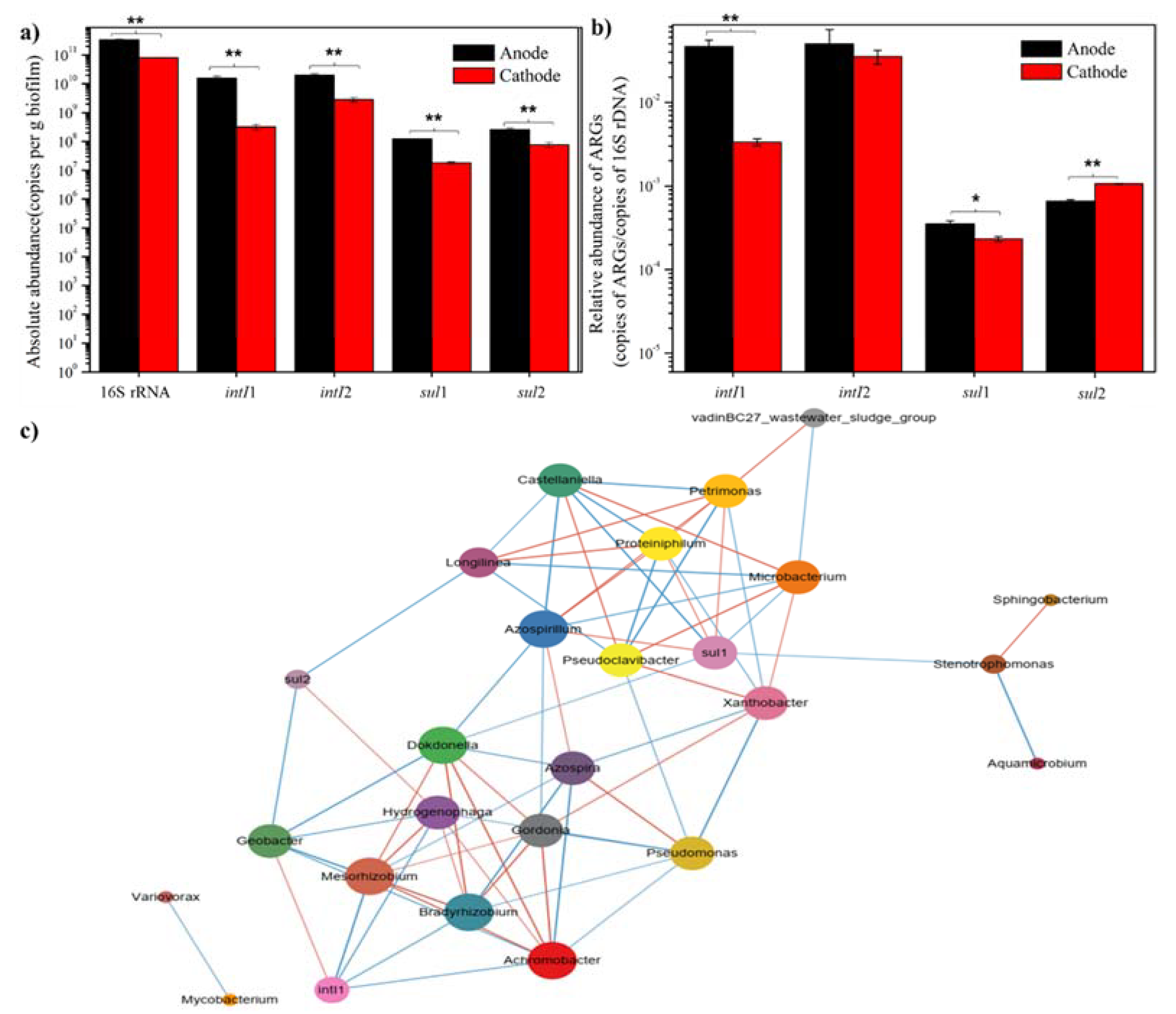{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. MFC Start-Up and Operation
2.3. Analytical Methods
2.3.1. Determination of Physicochemical Parameters and SDZ
2.3.2. Electrochemical Measurement
2.3.3. DNA Extraction and qPCR of ARGs
2.3.4. Bacterial Community Analysis
2.3.5. Data Analysis
3. Results and Discussion
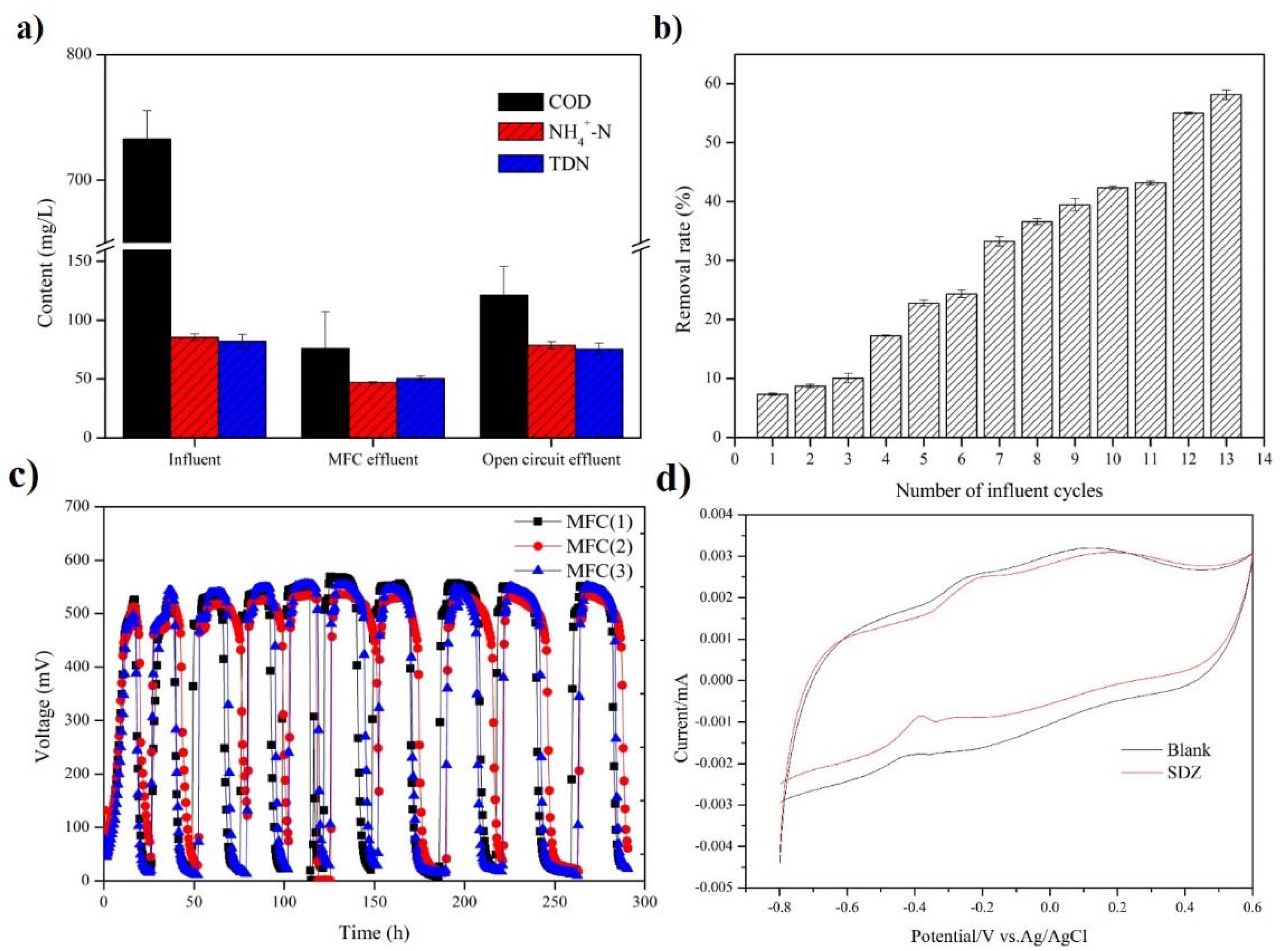
3.1. Performance of Air-Cathode MFCs under Sulfadiazine Pressure
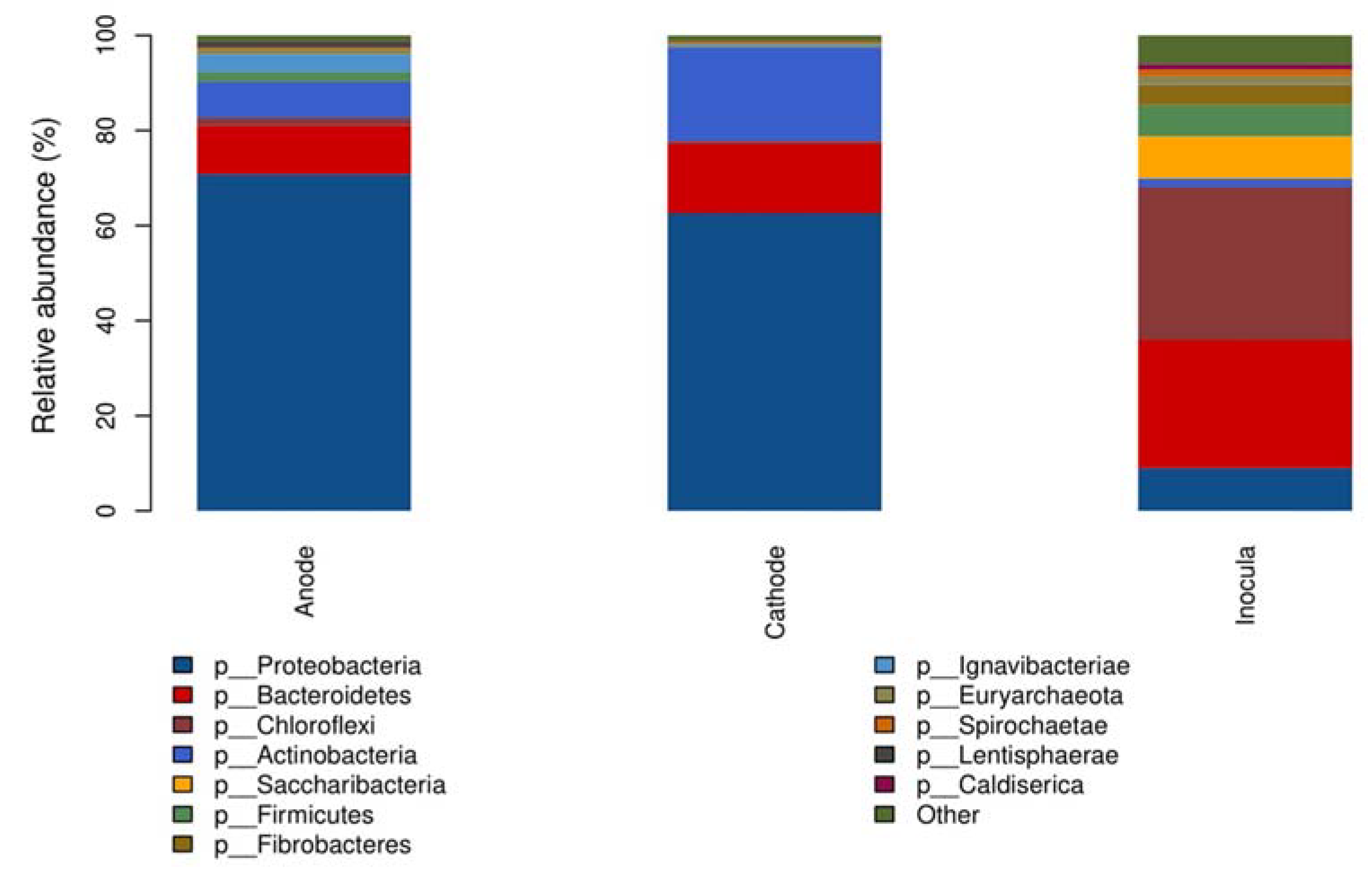
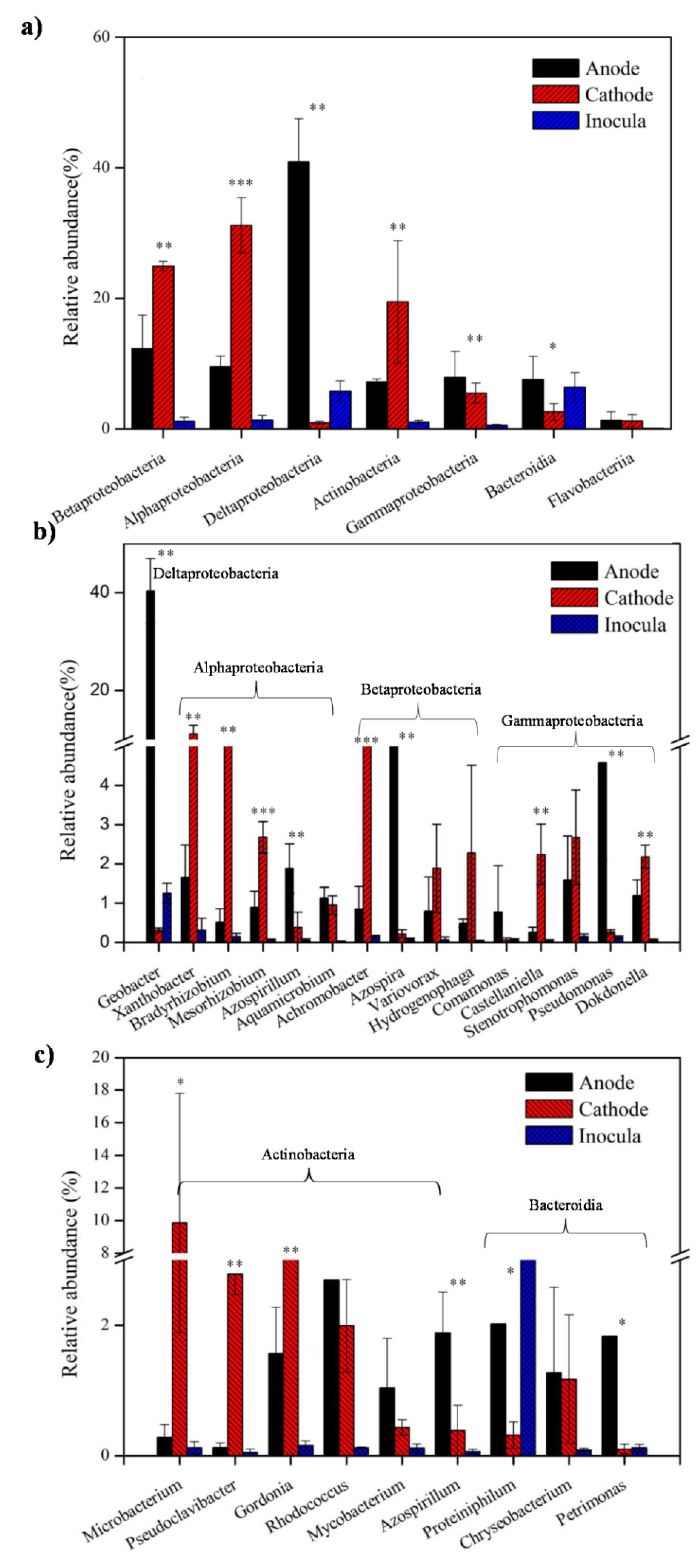
3.2. Bacterial Community Shift in the Anode and Cathode under Sulfadiazine Pressure
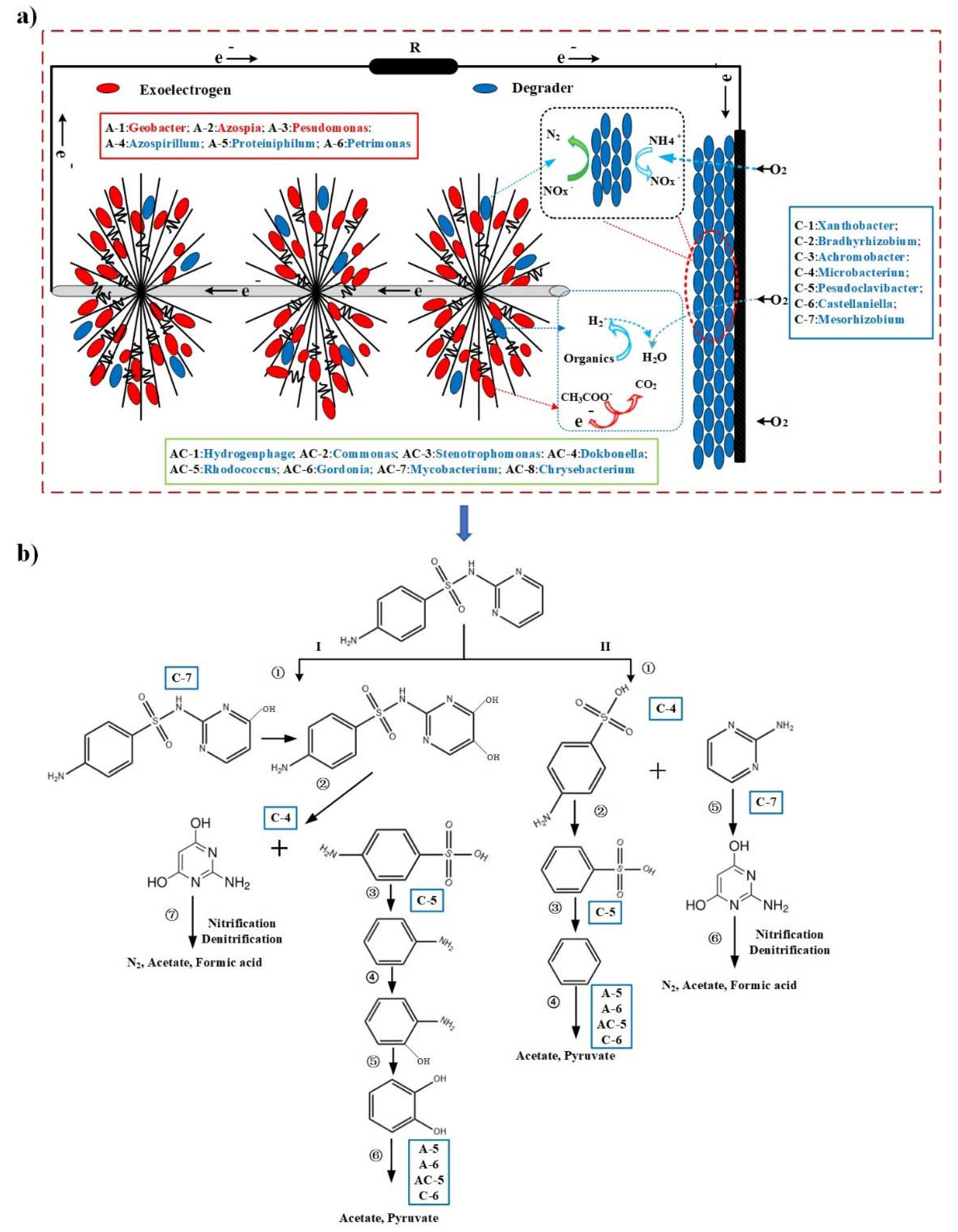
3.3. Potential Bacterial Roles in SDZ Degradation
3.4. ARGs in the Anode and Cathode under Sulfadiazine Pressure
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qiao, M.; Ying, G.; Singer, A.C.; Zhu, Y. Review of antibiotic resistance in China and its environment. Environ. Int. 2018, 110, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Logan, B.E.; Hamekers, B.; Rozendal, R.; Schroder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial fuel cells: Methodology and technology. Environ. Sci. Technol. 2018, 40, 5181–5192. [Google Scholar] [CrossRef]
- Wrighton, K.C.; Virdis, B.; Clauwaert, P.; Read, S.T.; Daly, R.A.; Boon, N.; Piceno, Y.; Andersen, G.L.; Coates, J.D.; Rabaey, K. Bacterial community structure corresponds to performance during cathodic nitrate reduction. ISME J. 2010, 4, 1443–1455. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.F.; Xiao, Y.; Yan, W.D.; Wang, S.H.; Zhao, F. The effect of bioelectrochemical systems on antibiotics removal and antibiotic resistance genes: A review. Chem. Eng. J. 2019, 358, 1421–1437. [Google Scholar] [CrossRef]
- Wang, L.; You, L.; Zhang, J.; Yang, T.; Zhang, W.; Zhang, Z.; Liu, P.; Wu, S.; Zhao, F.; Ma, J. Biodegradation of sulfadiazine in microbial fuel cells: Reaction mechanism, biotoxicity removal and the correlation with reactor microbes. J. Hazard. Mater. 2018, 360, 402–411. [Google Scholar] [CrossRef]
- Chen, J.; Hu, Y.; Huang, W.; Liu, Y.; Tang, M.; Zhang, L.; Sun, J. Biodegradation of oxytetracycline and electricity generation in microbial fuel cell with in situ dual graphene modified bioelectrode. Bioresour. Technol. 2018, 270, 482–488. [Google Scholar] [CrossRef]
- Wen, Q.; Kong, F.; Zheng, H.; Cao, D.; Ren, Y.; Yin, J. Electricity generation from synthetic penicillin wastewater in an air-cathode single chamber microbial fuel cell. Chem. Eng. J. 2011, 168, 572–576. [Google Scholar] [CrossRef]
- Xue, W.; Li, F.; Zhou, Q. Degradation mechanisms of sulfamethoxazole and its induction of bacterial community changes and antibiotic resistance genes in a microbial fuel cell. Bioresour. Technol. 2019, 289, 121632. [Google Scholar] [CrossRef]
- Song, X.; Jo, C.; Han, L.; Zhou, M. Recent advance in microbial fuel cell reactor configuration and coupling technologies for removal of antibiotic pollutants. Curr. Opin. Electrochem. 2022, 31, 100833. [Google Scholar] [CrossRef]
- Logan, B.E.; Rossi, R.; Ragab, A.; Saikaly, P.E. Electroactive microorganisms in bioelectrochemical systems. Nat. Rev. Microbiol. 2019, 17, 307–319. [Google Scholar] [CrossRef]
- Lovley, D.R. Happy together: Microbial communities that hook up to swap electrons. ISME J. 2017, 11, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Light, S.H.; Su, L.; Rivera-Lugo, R.; Cornejo, J.A.; Louie, A.; Iavarone, A.T.; Ajo-Franklin, C.M.; Portnoy, D.A. A flavin-based extracellular electron transfer mechanism in diverse gram-positive bacteria. Nature 2018, 562, 140–144. [Google Scholar] [CrossRef]
- Liu, W.; Cheng, S.; Sun, D.; Huang, H.; Chen, J.; Cen, K. Inhibition of microbial growth on air cathodes of single chamber microbial fuel cells by incorporating enrofloxacin into the catalyst layer. Biosens. Bioelectron. 2015, 72, 44–50. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhou, S.; Tang, J. In situ investigation of cathode and local biofilm microenvironments reveals important roles of OH- and oxygen transport in microbial fuel cells. Environ. Sci. Technol. 2013, 47, 4911–4917. [Google Scholar] [CrossRef]
- Yang, S.; Jia, B.; Liu, H. Effects of the Pt loading side and cathode-biofilm on the performance of a membrane-less and single-chamber microbial fuel cell. Bioresour. Technol. 2009, 100, 1197–1202. [Google Scholar] [CrossRef]
- Daghio, M.; Gandolfi, I.; Bestetti, G.; Franzetti, A.; Guerrini, E.; Cristiani, P. Anodic and cathodic microbial communities in single chamber microbial fuel cells. New Biotechnol. 2015, 32, 79–84. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Sun, Y.; Zhao, X.; Li, Y. Cathodic microbial community adaptation to the removal of chlorinated herbicide in soil microbial fuel cells. Environ. Sci. Pollut. Res. 2018, 25, 16900–16912. [Google Scholar] [CrossRef]
- Yuan, J.; Yuan, H.; Huang, S.; Liu, L.; Fu, F.; Zhang, Y.; Cheng, F.; Li, J. Comprehensive performance, bacterial community structure of single-chamber microbial fuel cell affected by COD/N ratio and physiological stratifications in cathode biofilm. Bioresour. Technol. 2021, 320, 124416. [Google Scholar] [CrossRef]
- Baran, W.; Adamek, E.; Ziemianska, J.; Sobczak, A. Effects of the presence of sulfonamides in the environment and their influence on human health. J. Hazard. Mater. 2011, 196, 1–15. [Google Scholar] [CrossRef]
- Song, T.; Zhu, C.; Xue, S.; Li, B.; Ye, J.; Geng, B.; Li, L.; Fahad Sardar, M.; Li, N.; Feng, S.; et al. Comparative effects of different antibiotics on antibiotic resistance during swine manure composting. Bioresour. Technol. 2020, 315, 123820. [Google Scholar] [CrossRef]
- Park, Y.; Yu, J.; Nguyen, V.K.; Park, S.; Kim, J.; Lee, T. Understanding complete ammonium removal mechanism in single-chamber microbial fuel cells based on microbial ecology. Sci. Total Environ. 2021, 764, 144231. [Google Scholar] [CrossRef]
- Zhao, X.; Li, X.; Li, Y.; Sun, Y.; Zhang, X.; Weng, L.; Ren, T.; Li, Y. Shifting interactions among bacteria, fungi and archaea enhance removal of antibiotics and antibiotic resistance genes in the soil bioelectrochemical remediation. Biotechnol. Biofuels 2019, 12, 8–15. [Google Scholar] [CrossRef]
- Cheng, D.; Ngo, H.H.; Guo, W.; Lee, D.; Duc Long, N.; Zhang, J.; Liang, S.; Varjani, S.; Wang, J. Performance of microbial fuel cell for treating swine wastewater containing sulfonamide antibiotics. Bioresour. Technol. 2020, 311, 123588. [Google Scholar] [CrossRef]
- Zhang, E.; Yu, Q.; Zhai, W.; Wang, F.; Scott, K. High tolerance of and removal of cefazolin sodium in single-chamber microbial fuel cells operation. Bioresour. Technol. 2018, 249, 76–81. [Google Scholar] [CrossRef]
- Bond, D.R.; Lovley, D.R. Electricity production by Geobacter sulfurreducens attached to electrodes. Appl. Environ. Microbiol. 2003, 69, 1548–1555. [Google Scholar] [CrossRef] [Green Version]
- Holmes, D.E.; Dang, Y.; Walker, D.J.F.; Lovley, D.R. The electrically conductive pili of Geobacter species are a recently evolved feature for extracellular electron transfer. Microb. Genom. 2016, 2, e000072. [Google Scholar] [CrossRef]
- Ishii, S.; Suzuki, S.; Norden-Krichmar, T.M.; Phan, T.; Wanger, G.; Nealson, K.H.; Sekiguchi, Y.; Gorby, Y.A.; Bretschger, O. Microbial population and functional dynamics associated with surface potential and carbon metabolism. ISME J. 2014, 8, 963–978. [Google Scholar] [CrossRef] [Green Version]
- Shen, R.; Liu, Z.; He, Y.; Zhang, Y.; Lu, J.; Zhu, Z.; Si, B.; Zhang, C.; Xing, X.-H. Microbial electrolysis cell to treat hydrothermal liquefied wastewater from cornstalk and recover hydrogen: Degradation of organic compounds and characterization of microbial community. Int. J. Hydrogen Energy. 2016, 41, 4132–4142. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Nghiem, L.D.; Oh, S. Aerobic biotransformation of the antibiotic ciprofloxacin by Bradyrhizobium sp. isolated from activated sludge. Chemosphere 2018, 211, 600–607. [Google Scholar] [CrossRef]
- Kumru, M.; Eren, H.; Catal, T.; Bermek, H.; Akarsubasi, A.T. Study of azo dye decolorization and determination of cathode microorganism profile in air-cathode microbial fuel cells. Environ. Technol. 2012, 33, 2167–2175. [Google Scholar] [CrossRef]
- Bashan, Y.; Holguin, G.; de-Bashan, L.E. Azospirillum-plant relationships: Physiological, molecular, agricultural, and environmental advances (1997–2003). Can. J. Microbiol. 2004, 50, 521–577. [Google Scholar] [CrossRef] [Green Version]
- Reis, P.J.; Reis, A.C.; Ricken, B.; Kolvenbach, B.A.; Manaia, C.M.; Corvini, P.F.; Nunes, O.C. Biodegradation of sulfamethoxazole and other sulfonamides by achromobacter denitrificans PR1. J. Hazard. Mater. 2014, 280, 741–749. [Google Scholar] [CrossRef]
- Deng, L.; Ren, Y.; Wei, C.; Wang, J. Biodegradation of pyrene by a novel strain of Castellaniella sp. under denitrifying condition. J. Environ. Chem. Eng. 2021, 9, 104970. [Google Scholar] [CrossRef]
- Borole, A.P.; Hamilton, C.Y.; Vishnivetskaya, T.; Leak, D.; Andras, C. Improving power production in acetate-fed microbial fuel cells via enrichment of exoelectrogenic organisms in flow-through systems. Biochem. Eng. J. 2009, 48, 71–80. [Google Scholar] [CrossRef]
- Huang, J.; Wang, Z.; Zhu, C.; Ma, J.; Zhang, X.; Wu, Z. Identification of microbial communities in open and closed circuit bioelectrochemical MBRs by high-throughput 454 pyrosequencing. PLoS ONE 2014, 9, e93842. [Google Scholar] [CrossRef]
- Xing, D.; Cheng, S.; Logan, B.E.; Regan, J.M. Isolation of the exoelectrogenic denitrifying bacterium Comamonas denitrificans based on dilution to extinction. Appl. Microbiol. Biotechnol. 2010, 85, 1575–1587. [Google Scholar] [CrossRef]
- Chen, J.; Xie, S. Overview of sulfonamide biodegradation and the relevant pathways and microorganisms. Sci. Total Environ. 2018, 640, 1465–1477. [Google Scholar] [CrossRef]
- Ryan, R.P.; Monchy, S.; Cardinale, M.; Taghavi, S.; Crossman, L.; Avison, M.B.; Berg, G.; van der Lelie, D.; Dow, J.M. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat. Rev. Microbiol. 2009, 7, 514–525. [Google Scholar] [CrossRef]
- Jiang, X.; Shen, J.; Xu, K.; Chen, D.; Mu, Y.; Sun, X.; Han, W.; Li, J.; Wang, L. Substantial enhancement of anaerobic pyridine bio-mineralization by electrical stimulation. Water Res. 2018, 130, 291–299. [Google Scholar] [CrossRef]
- Huerta, B.; Marti, E.; Gros, M.; Lopez, P.; Pompeo, M.; Armengol, J.; Barcelo, D.; Balcazar, J.L.; Rodriguez-Mozaz, S.; Marce, R. Exploring the links between antibiotic occurrence, antibiotic resistance, and bacterial communities in water supply reservoirs. Sci. Total Environ. 2013, 456–457, 161–170. [Google Scholar] [CrossRef]
- Chen, J.; Yang, Y.; Liu, Y.; Tang, M.; Wang, R.; Tian, Y.; Jia, C. Bacterial community shift and antibiotics resistant genes analysis in response to biodegradation of oxytetracycline in dual graphene modified bioelectrode microbial fuel cell. Bioresour. Technol. 2019, 276, 236–243. [Google Scholar] [CrossRef]
- Tappe, W.; Hofmann, D.; Disko, U.; Koeppchen, S.; Kummer, S.; Vereecken, H. A novel isolated Terrabacter-like bacterium can mineralize 2-aminopyrimidine, the principal metabolite of microbial sulfadiazine degradation. Biodegradation 2015, 26, 139–150. [Google Scholar] [CrossRef]
- Oyaert, M.; De Baere, T.; Breyne, J.; De Laere, E.; Marien, S.; Waets, P.; Laffut, W. First case of Pseudoclavibacter bifida bacteremia in an immunocompromised host with chronic obstructive pulmonary disease (COPD). J. Clin. Microbiol. 2013, 51, 1973–1976. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.J.; Kulakov, L.A.; Allen, C.C. Biodegradation and rhodococcus—Masters of catabolic versatility. Curr. Opin. Biotechnol. 2005, 16, 282–290. [Google Scholar] [CrossRef]
- Taskan, B.; Taskan, E. Inhibition of AHL-mediated quorum sensing to control biofilm thickness in microbial fuel cell by using Rhodococcus sp. BH4. Chemosphere 2021, 285, 131538. [Google Scholar] [CrossRef]
- Hamdan, H.Z.; Salam, D.A. Response of sediment microbial communities to crude oil contamination in marine sediment microbial fuel cells under ferric iron stimulation. Environ. Pollut. 2020, 263, 114658. [Google Scholar] [CrossRef]
- Arenskotter, M.; Broker, D.; Steinbuchel, A. Biology of the metabolically diverse genus Gordonia. Appl. Environ. Microbiol. 2004, 70, 3195–3204. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Quan, X.; Chen, L.; Zheng, Y. Application of slow-release carbon sources embedded in polymer for stable and extended power generation in microbial fuel cells. Chemosphere 2020, 244, 125515. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y. Application of ethanol-type fermentation in establishment of direct interspecies electron transfer: A practical engineering case study. Renew. Energy 2019, 136, 846–855. [Google Scholar] [CrossRef]
- Nishioka, T.; Elsharkawy, M.M.; Suga, H.; Kageyama, K.; Hyakumachi, M.; Shimizu, M. Development of culture medium for the isolation of flavobacterium and chryseobacterium from rhizosphere soil. Microbes Environ. 2016, 31, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.; Yan, N.; Liu, X.; Wang, W.; Zhang, Y.; Liu, R.; Rittmann, B.E. How UV photolysis accelerates the biodegradation and mineralization of sulfadiazine (SD). Biodegradation 2014, 25, 911–921. [Google Scholar] [CrossRef]
- Tadkaew, N.; Hai, F.I.; McDonald, J.A.; Khan, S.J.; Nghiem, L.D. Removal of trace organics by MBR treatment: The role of molecular properties. Water Res. 2011, 45, 2439–2451. [Google Scholar] [CrossRef] [Green Version]
- Tappe, W.; Herbst, M.; Hofmann, D.; Koeppchen, S.; Kummer, S.; Vereecken, H. Degradation of sulfadiazine by microbacterium lacus strain sdzm4, isolated from lysimeters previously manured with slurry from sulfadiazine-medicated pigs. Appl. Environ. Microb. 2013, 79, 2572–2577. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Mao, D.; Ryse, M.; Zhou, Q.; Zhang, H.; Xu, L.; Alvarez, P.J.J. Trends in antibiotic resistance genes occurrence in the Haihe river, China. Environ. Sci. Technol. 2010, 77, 7220–7225. [Google Scholar] [CrossRef]
- Ji, X.; Shen, Q.; Liu, F.; Ma, J.; Xu, G.; Wang, Y.; Wu, M. Antibiotic resistance gene abundances associated with antibiotics and heavy metals in animal manures and agricultural soils adjacent to feedlots in Shanghai; China. J. Hazard. Mater. 2012, 235–236, 178–185. [Google Scholar] [CrossRef]
- Liu, L.; Liu, Y.; Wang, Z.; Liu, C.; Huang, X.; Zhu, G. Behavior of tetracycline and sulfamethazine with corresponding resistance genes from swine wastewater in pilot-scale constructed wetlands. J. Hazard. Mater. 2014, 278, 304–310. [Google Scholar] [CrossRef]
- Su, Y.; Wang, J.; Xia, H.; Xie, B.; Li, X. Anaerobic/aerobic conditions determine antibiotic resistance genes removal patterns from leachate by affecting bacteria taxa-genes co-occurrence modules. Chemosphere 2019, 223, 28–38. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, Q.; Wang, X.; Logan, B.E. Treatment of carbon fiber brush anodes for improving power generation in air–cathode microbial fuel cells. J. Power Sources. 2010, 195, 1841–1844. [Google Scholar] [CrossRef]
- Liu, H.; Logan, B.E. Electricity generation using an air-cathode single chamber microbial fuel cell in the presence and absence of a proton exchange membrane. Environ. Sci. Technol. 2004, 38, 4040–4046. [Google Scholar] [CrossRef]
- Lovley, D.R.; Phillips, E. Novel mode of microbial energy metabolism: Organic carbon oxidation coupled to dissimilatory reduction of iron or manganese. Appl. Environ. Microb. 1988, 54, 1472–1480. [Google Scholar] [CrossRef] [Green Version]
- Pei, R.; Kim, S.C.; Carlson, K.H.; Pruden, A. Effect of river landscape on the sediment concentrations of antibiotics and corresponding antibiotic resistance genes (ARG). Water. Res. 2006, 40, 2427–2435. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Li, H.; Li, N.; Sardar, M.F.; Song, T.; Zhu, H.; Xing, X.; Zhu, C. Dynamics of a Bacterial Community in the Anode and Cathode of Microbial Fuel Cells under Sulfadiazine Pressure. Int. J. Environ. Res. Public Health 2022, 19, 6253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19106253
Yang Z, Li H, Li N, Sardar MF, Song T, Zhu H, Xing X, Zhu C. Dynamics of a Bacterial Community in the Anode and Cathode of Microbial Fuel Cells under Sulfadiazine Pressure. International Journal of Environmental Research and Public Health. 2022; 19(10):6253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19106253
Chicago/Turabian StyleYang, Zhenzhen, Hongna Li, Na Li, Muhammad Fahad Sardar, Tingting Song, Hong Zhu, Xuan Xing, and Changxiong Zhu. 2022. "Dynamics of a Bacterial Community in the Anode and Cathode of Microbial Fuel Cells under Sulfadiazine Pressure" International Journal of Environmental Research and Public Health 19, no. 10: 6253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19106253