Pulsed Ultraviolet Light Reduces Immunoglobulin E Binding to Atlantic White Shrimp (Litopenaeus setiferus) Extract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Preparation of Atlantic White Shrimp Crude Protein Extract
2.3. Preparation of Boiled Shrimp Extract
2.4. Treatment of Shrimp Extract with PUV
2.5. Electrophoresis of Treated Shrimp Extract
2.6. Determination of IgE- and IgG-Binding to Tropomyosin with Western Blot
2.7. Determination of IgE Binding to Treated Shrimp Extract with Dot Blot
2.8. Determination of IgE Binding to Treated Shrimp Extracts with Indirect ELISA
2.9. Statistical Analysis
3. Results and Discussion
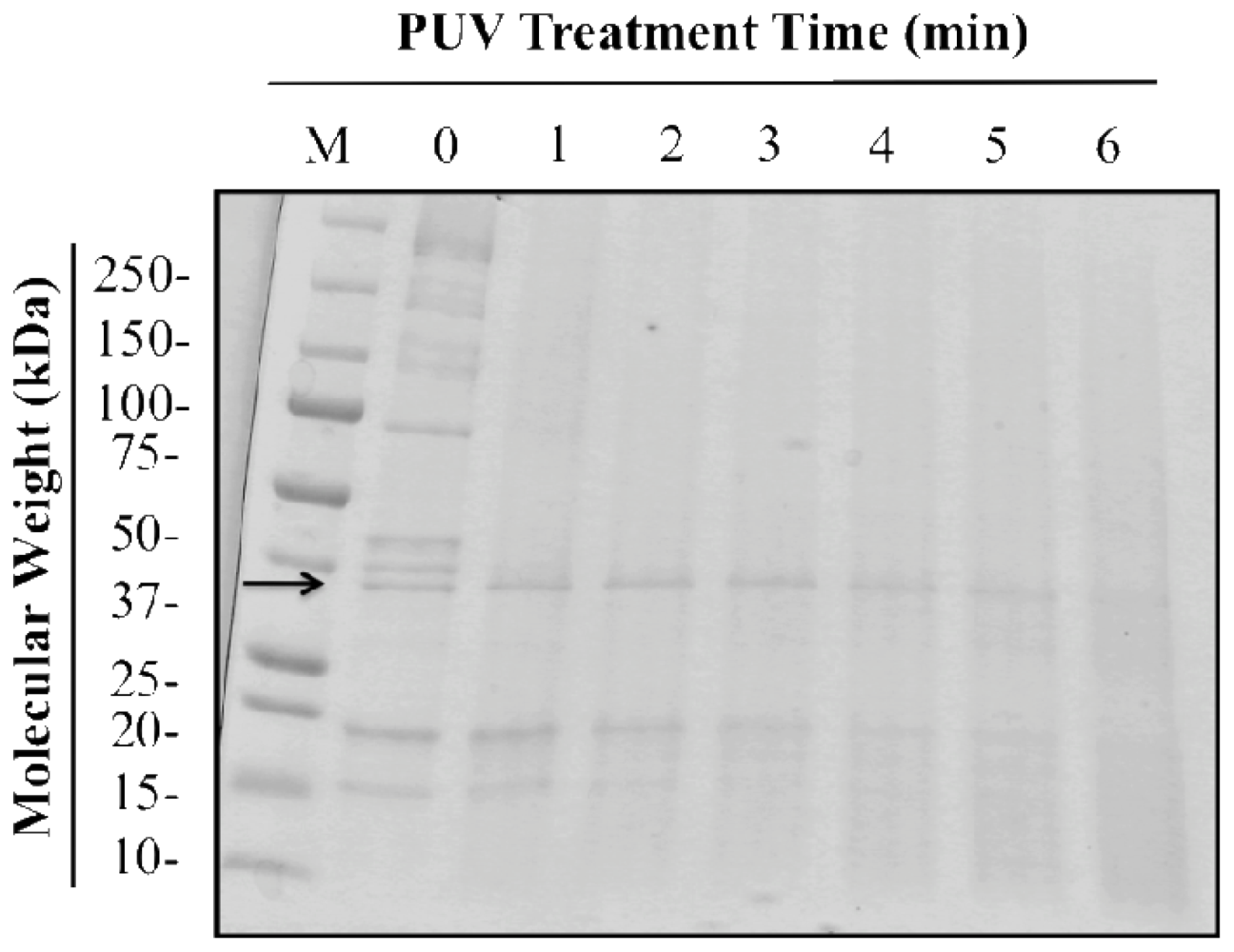
3.1. Optimal PUV Treatment Time for Shrimp Extract
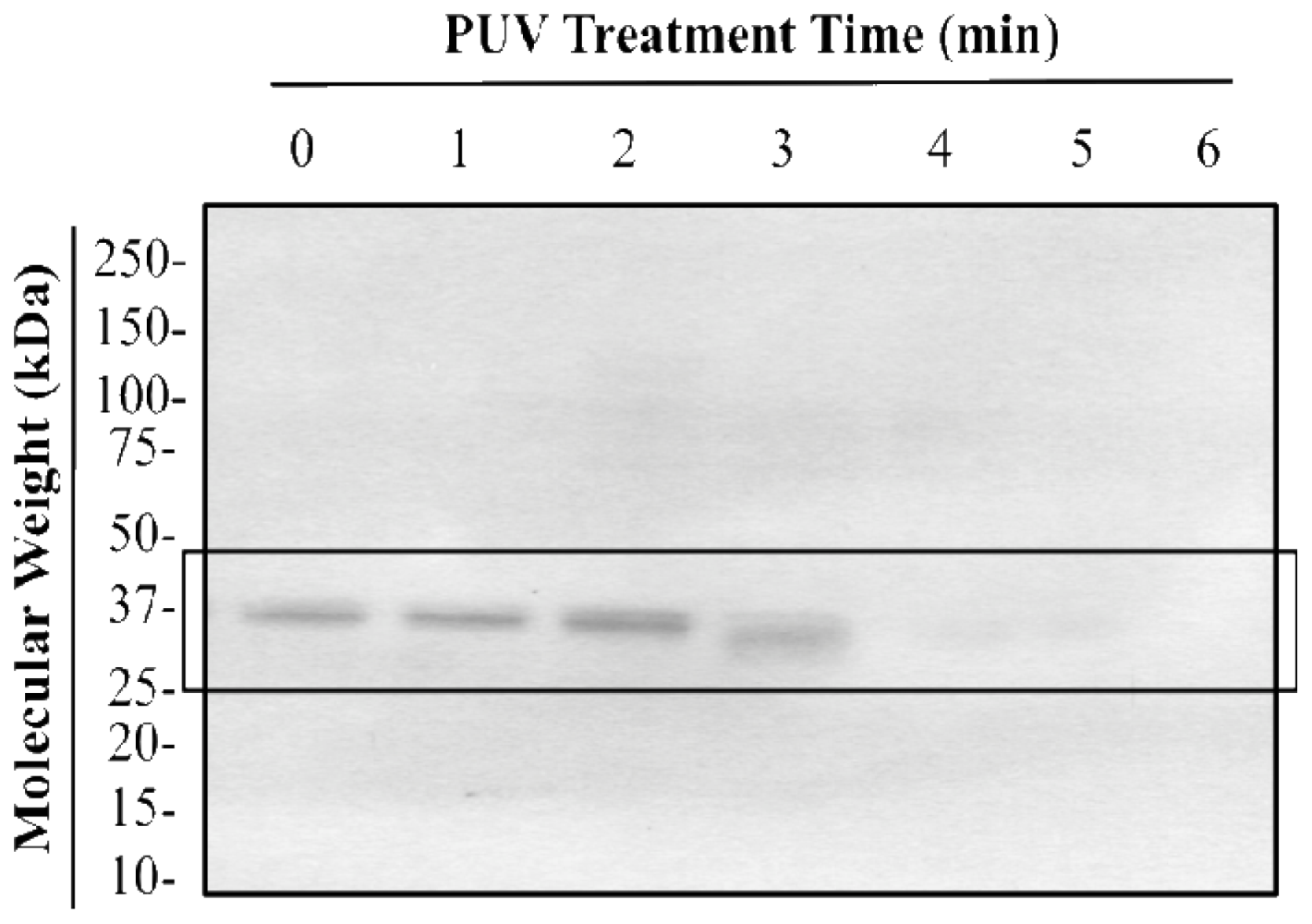
3.2. Changes in Tropomyosin Band Intensity of Untreated, Boiled, PUV-Treated, and [Boiled+PUV]-Treated Shrimp Extracts
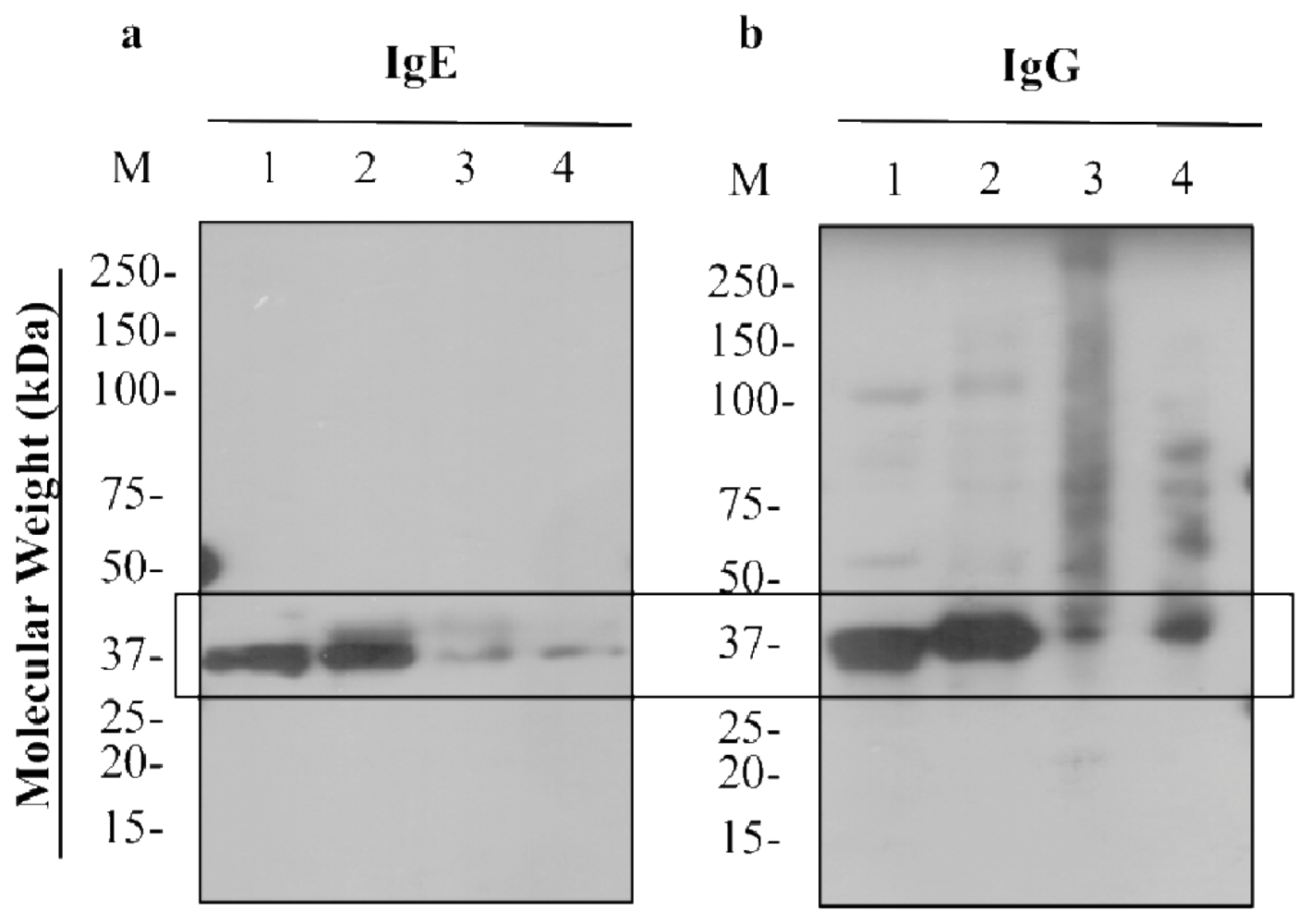
3.3. IgE- and IgG-binding to Untreated, Boiled, PUV-treated, and [Boiled+PUV]-Treated Shrimp Extracts
3.3.1. Western Blot
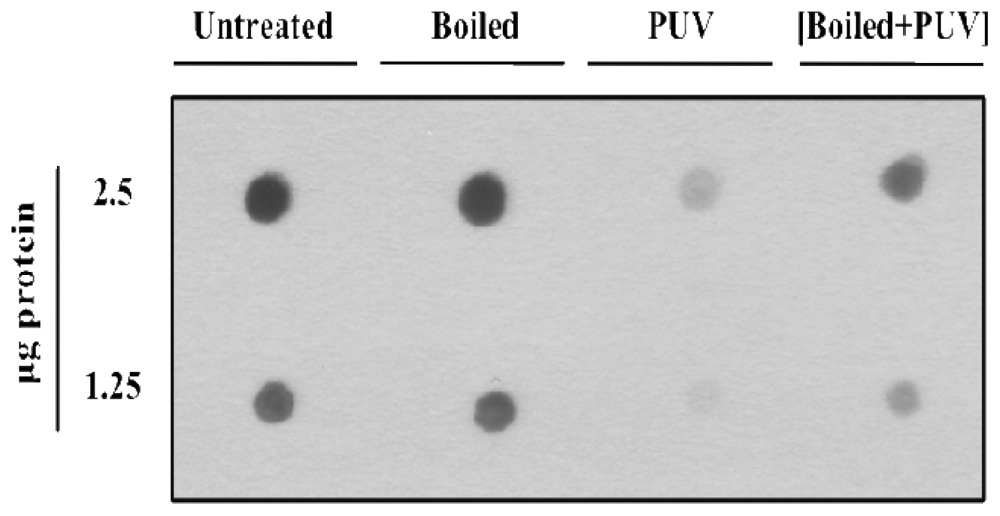
3.3.2. Dot Blot
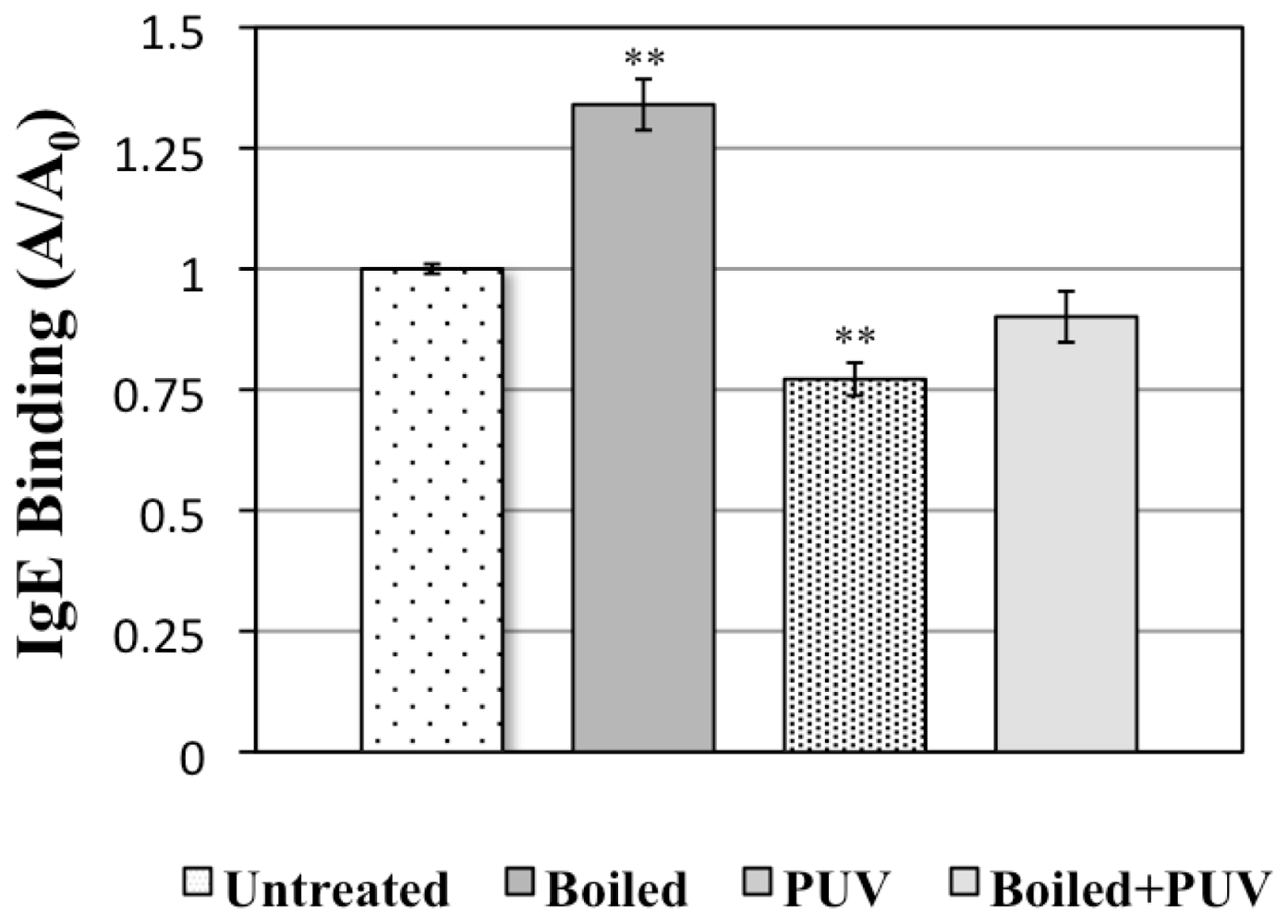
3.3.3. Indirect ELISA
3.4. Temperature and Volume Changes Following Treatment of Shrimp Extract
4. Conclusions
Acknowledgments
References
- Ellman, LK; Chatchatee, P; Sicherer, SH; Sampson, HA. Food hypersensitivity in two groups of children and young adults with atopic dermatitis evaluated a decade apart. Pediatr. Allergy. Immunol 2002, 13, 295–298. [Google Scholar]
- Sicherer, SH; Sampson, HA. Food allergy. J. Allergy. Clin. Immunol 2010, 125, S116–125. [Google Scholar]
- Shanti, KN; Martin, BM; Nagpal, S; Metcalfe, DD; Rao, PV. Identification of tropomyosin as the major shrimp allergen and characterization of its IgE-binding epitopes. J. Immunol. 1993, 151, 5354–5363. [Google Scholar]
- Jeoung, BJ; Reese, G; Hauck, P; Oliver, JB; Daul, CB; Lehrer, SB. Quantification of the major brown shrimp allergen Pen a 1 (tropomyosin) by a monoclonal antibody-based sandwich ELISA. J. Allergy. Clin. Immunol 1997, 100, 229–234. [Google Scholar]
- Reese, G; Ayuso, R; Lehrer, SB. Tropomyosin: an invertebrate pan-allergen. Int. Arch. Allergy. Immunol 1999, 119, 247–258. [Google Scholar]
- Jeong, KY; Hong, CS; Yong, TS. Allergenic tropomyosins and their cross-reactivities. Protein Pept. Lett 2006, 13, 835–845. [Google Scholar]
- Ayuso, R; Grishina, G; Bardina, L; Carrillo, T; Blanco, C; Ibanez, MD; Sampson, HA; Beyer, K. Myosin light chain is a novel shrimp allergen, Lit v 3. J. Allergy. Clin. Immunol 2008, 122, 795–802. [Google Scholar]
- Yu, CJ; Lin, YF; Chiang, BL; Chow, LP. Proteomics and immunological analysis of a novel shrimp allergen, Pen m 2. J. Immunol 2003, 170, 445–453. [Google Scholar]
- Garcia-Orozco, KD; Aispuro-Hernandez, E; Yepiz-Plascencia, G; Calderon-de-la-Barca, AM; Sotelo-Mundo, RR. Molecular characterization of arginine kinase, an allergen from the shrimp Litopenaeus vannamei. Int. Arch. Allergy. Immunol 2007, 144, 23–28. [Google Scholar]
- Shiomi, K; Sato, Y; Hamamoto, S; Mita, H; Shimakura, K. Sarcoplasmic calcium-binding protein: identification as a new allergen of the black tiger shrimp Penaeus monodon. Int. Arch. Allergy. Immunol 2008, 146, 91–98. [Google Scholar]
- Daul, CB; Slattery, M; Reese, G; Lehrer, SB. Identification of the major brown shrimp (Penaeus aztecus) allergen as the muscle protein tropomyosin. Int. Arch. Allergy. Immunol 1994, 105, 49–55. [Google Scholar]
- Skripak, JM; Sampson, HA. Towards a cure for food allergy. Curr. Opin. Immunol 2008, 20, 690–696. [Google Scholar]
- Tanabe, S. Epitope Peptides and Immunotherapy. Curr. Protein Pept. Sci 2007, 8, 109–118. [Google Scholar]
- Li, Z; Lin, H; Cao, LM; Jameel, K. Effect of high intensity ultrasound on the allergenicity of shrimp. J. Zhejiang Univ. Sci. B 2006, 7, 251–256. [Google Scholar]
- Zhenxing, L; Hong, L; Limin, C; Jamil, K. The influence of gamma irradiation on the allergenicity of shrimp (Penaeus vannamei). J. Food Eng 2007, 79, 945–949. [Google Scholar]
- Hildebrandt, S; Schutte, L; Stoyanov, S; Hammer, G; Steinhart, H; Paschke, A. In vitro determination of the allergenic potential of egg white in processed meat. J. Allergy. (Cairo) 2010, 2, 155–165. [Google Scholar]
- Chung, SY; Yang, W; Krishnamurthy, K. Effects of pulsed UV-light on peanut allergens in extracts and liquid peanut butter. J. Food Sci 2008, 73, C400–404. [Google Scholar]
- Yang, W; Mwakatage, NR; Goodrich-Schneider, R; Krishnamurthy, K; Rababah, TM. Mitigation of major peanut allergens by pulsed ultraviolet light. Food Bioprocess Technol 2011. [Google Scholar] [CrossRef]
- Yang, WW; Chung, SY; Ajayi, O; Krishnamurthy, K; Konan, K; Goodrich-Schneider, R. Use of pulsed ultraviolet light to reduce the allergenic potency of soybean extracts. J. Food Eng. 2010, 6, 1–12. [Google Scholar]
- Shriver, SK; Yang, W. Thermal and nonthermal methods for allergen control. Food Eng. Rev 2011, 3, 26–43. [Google Scholar]
- Krishnamurthy, K; Demirci, A; Irudayaraj, JM. Inactivation of Staphylococcus aureus in milk using flow-through pulsed UV-light treatment system. J. Food Sci 2007, 72, M233–239. [Google Scholar]
- Wakatage, NR. Efficacy of Pulsed UV Light Treatment on Removal of Peanut Allergens. MS Thesis, Department of Food and Animal Sciences, Alabama A&M University, Normal, AL, USA, 2008. [Google Scholar]
- Li, Y. Effect of Pulsed Ultraviolet Light, High Hydrostatic Pressure and Non-thermal Plasma on the Antigenicity of Almond. MS Thesis, Department of Agricultural and Biological Engineering, University of Florida, Gainesville, FL, USA, 2011. [Google Scholar]
- Nooji, J. Reduction of Wheat Allergen Potency by Pulsed Ultraviolet Light, High Hydrostactic Pressure and Nonthermal Plasma. MS Thesis, Department of Food Science and Human Nutrition, University of Florida, Gainesville, FL, USA, 2011. [Google Scholar]
- Shriver, SK. Effect of Selected Emerging Nonthermal Processing Methods on the Allergen Reactivity of ATLANTIC White Shrimp (Litopenaeus Setiferus). MS Thesis, Department of Food Science and Human Nutrition, University of Florida, Gainesville, FL, USA, 2011. [Google Scholar]
- Fiedorowicz, M; Tomasik, P; Lii, CY. Degradation of starch by polarised light. Carbohydr. Polymer 2001, 45, 79–87. [Google Scholar]
- Greenberg, JR. Ultraviolet light-induced crosslinking of mRNA to proteins. Nucleic Acids Res 1979, 6, 715–32. [Google Scholar]
- Kramer, GF; Norman, HA; Krizek, DT; Mirecki, RM. Influence of UV-B radiation on polyamines, lipid peroxidation and membrane lipids in cucumber. Phytochemistry 1991, 30, 2101–2108. [Google Scholar]
- Friso, G; Barbato, R; Giacometti, GM; Barber, J. Degradation of D2 protein due to UV-B irradiation of the reaction centre of photosystem II. FEBS Letters 1994, 339, 217–221. [Google Scholar]
- Motoyama, K; Suma, Y; Ishizaki, S; Nagashima, Y; Lu, Y; Ushio, H; Shiomi, K. Identification of tropomyosins as major allergens in Antarctic krill and mantis shrimp and their amino acid sequence characteristics. Mar. Biotechnol 2008, 10, 709–718. [Google Scholar]
- Motoyama, K; Suma, Y; Ishizaki, S; Nagashima, Y; Shiomi, K. Molecular cloning of tropomyosins identified as allergens in six species of crustaceans. J. Agric. Food Chem 2007, 55, 985–991. [Google Scholar]
- Carnes, J; Ferrer, A; Huertas, AJ; Andreu, C; Larramendi, CH; Fernandez-Caldas, E. The use of raw or boiled crustacean extracts for the diagnosis of seafood allergic individuals. Ann. Allergy Asthma Immunol 2007, 98, 349–354. [Google Scholar]
- Daul, CB; Morgan, JE; Hughes, J; Lehrer, SB. Provocation-challenge studies in shrimp-sensitive individuals. J. Allerg. Clin. Immunol 1988, 81, 1180–1186. [Google Scholar]
- Naqpal, S; Rajappa, L; Metcalfe, DD; Rao, PV. Isolation and characterization of heat-stable allergens from shrimp (Penaeus indicus). J. Allerg. Clin. Immunol 1989, 83, 26–36. [Google Scholar]
- Liu, G-M; Cheng, H; Nesbit, JB; Su, W-J; Gao, M-J; Maleki, SJ. Effects of Boiling on the IgE-Binding Properties of Tropomyosin of Shrimp (Litopenaeus vannamei). J. Food. Sci 2010, 75, T1–T5. [Google Scholar]
- Laemmli, UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]
- Samson, KTR; Chen, FH; Miura, K; Odajima, Y; Likura, Y; Rivas, MN; Minoguchi, K; Adachi, M. IgE Binding to Raw and Boiled Shrimp Proteins in Atopic and Nonatopic Patients with Adverse Reactions to Shrimp. Int. Arch. Allergy. Immunol. 2004, 133, 225–232. [Google Scholar]
- Muranov, K; Maloletkina, O; Poliansky, N; Kleymenov, S; Rozhkov, S; Goryunov, A; Ostrovsky, M; Kurganov, B. Mechanism of aggregation of UV-irradiated XL–crystallin. Exper. Eye. Res 2011, 92, 76–86. [Google Scholar]
- Gomez-Lopez, V; Ragaert, P; Debevere, J; Devlieghere, F. Pulsed light for food decontamination: a review. Trends Food Sci. Technol 2007, 18, 464–473. [Google Scholar]
- Chung, S-Y; Kato, Y; Champagne, ET. Polyphenol oxidase/caffeic acid may reduce the allergenic properties of peanut allergens. J Sci Food Agr 2005, 2631–2637. [Google Scholar]
- Maleki, SJ; Chung, SY; Champagne, ET; Raufman, JP. The effects of roasting on the allergenic properties of peanut proteins. J. Allergy. Clin. Immunol 2000, 106, 763–768. [Google Scholar]
- Taheri-Kafrani, A; Gaudin, JC; Rabesona, H; Nioi, C; Agarwal, D; Drouet, M; Chobert, JM; Bordbar, AK; Haertlft, T. Effects of heating and glycation of beta-lactoglobulin on its recognition by IgE of sera from cow milk allergy patients. J. Agr. Food. Chem 2009, 57, 4974–4982. [Google Scholar]
- Nakamura, A; Watanabe, K; Ojima, T; Ahn, DH; Saeki, H. Effect of maillard reaction on allergenicity of scallop tropomyosin. J. Agr. Food. Chem 2005, 53, 7559–7564. [Google Scholar]
- Nakamura, A; Sasaki, F; Watanabe, K; Ojima, T; Ahn, DH; Saeki, H. Changes in allergenicity and digestibility of squid tropomyosin during the Maillard reaction with ribose. J. Agr. Food. Chem 2006, 54, 9529–9534. [Google Scholar]
- Hansen, KS; Ballmer-Weber, BK; Luttkopf, D; Skov, PS; Wuthrich, B; Bindslev-Jensen, C; Vieths, S; Poulsen, LK. Roasted hazelnuts--allergenic activity evaluated by double-blind, placebo-controlled food challenge. Allergy 2003, 58, 132–138. [Google Scholar]
- Nowak-Wegrzyn, A; Bloom, KA; Sicherer, SH; Shreffler, WG; Noone, S; Wanich, N; Sampson, HA. Tolerance to extensively heated milk in children with cow's milk allergy. J. Allergy Clin. Immunol 2008, 122, 342–347. [Google Scholar]
- Keklik, NM; Demirci, A; Puri, VM. Inactivation of Listeria monocytogenes on unpackaged and vacuum-packaged chicken frankfurters using pulsed UV-light. J. Food Sci 2009, 74, M431–M439. [Google Scholar]






© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shriver, S.; Yang, W.; Chung, S.-Y.; Percival, S. Pulsed Ultraviolet Light Reduces Immunoglobulin E Binding to Atlantic White Shrimp (Litopenaeus setiferus) Extract. Int. J. Environ. Res. Public Health 2011, 8, 2569-2583. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph8072569
Shriver S, Yang W, Chung S-Y, Percival S. Pulsed Ultraviolet Light Reduces Immunoglobulin E Binding to Atlantic White Shrimp (Litopenaeus setiferus) Extract. International Journal of Environmental Research and Public Health. 2011; 8(7):2569-2583. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph8072569
Chicago/Turabian StyleShriver, Sandra, Wade Yang, Si-Yin Chung, and Susan Percival. 2011. "Pulsed Ultraviolet Light Reduces Immunoglobulin E Binding to Atlantic White Shrimp (Litopenaeus setiferus) Extract" International Journal of Environmental Research and Public Health 8, no. 7: 2569-2583. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph8072569