Protective Effect of Anthocyanins from Lingonberry on Radiation-induced Damages

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Anthocyanins from Lingonberry
2.3. Animals
2.4. Irradiation
2.5. Blood Cell Count
2.6. Thymus and Spleen Indices

2.7. Measurement of Carbon Clearance and Phagocytic Index (PI)

2.8. Splenocyte Proliferation Index
2.9. Effect of ALB on Bone Marrow Micronuclei Formation
2.10. Statistical Analysis
3. Results
3.1. Effect of ALB on Body Weight and Peripheral Blood Counts of Irradiated Mice


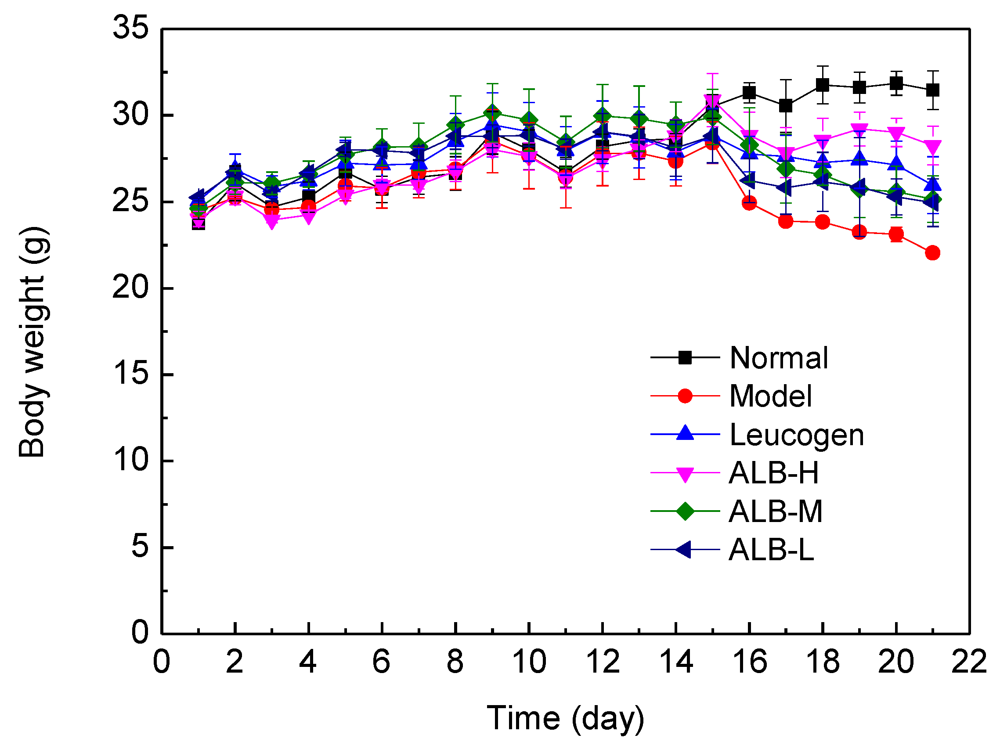{kind=link}
{kind=link}
{kind=link}
| Group | WBC (109/L) | RBC (1012/L) | HGB (G/L) | PLT (109/L) |
|---|---|---|---|---|
| Normal | 5.81 ± 0.66 | 9.62 ± 1.48 | 141.14 ± 14.7 | 811.71 ± 106.84 |
| Model | 0.45 ± 0.26** | 3.54 ± 0.92** | 28.14 ± 3.33** | 63.29 ± 19.46** |
| Leucogen (1.4 mg/kg) | 0.81 ± 0.26**▲ | 9.17 ± 1.40▲▲ | 39.29 ± 9.79**▲ | 208.14 ± 83.14**▲▲ |
| ALB-H (200 mg/kg) | 0.71 ± 0.12**▲ | 9.28 ± 1.10*▲▲ | 102.14 ± 7.76**▲▲ | 219.43 ± 73.89**▲▲ |
| ALB-M (100 mg/kg) | 0.59 ± 0.18** | 7.64 ± 1.11▲▲ | 67.43 ± 6.50**▲▲ | 286.86 ± 85.55**▲▲ |
| ALB-L (50 mg/kg) | 0.61 ± 0.17** | 8.07 ± 0.91*▲▲ | 62.71 ± 9.50**▲▲ | 127.57 ± 58.55**▲ |
3.2. Effect of ALB on Immune Function
| Group | n | Spleen index | Thymus index | Phagocytic capacity |
|---|---|---|---|---|
| Normal | 10 | 4.24 ± 0.17 | 3.06 ± 0.17 | 7.33 ± 0.25 |
| Model | 10 | 2.02 ± 0.11** | 1.22 ± 0.03** | 5.92 ± 0.38** |
| Leucogen (1.4 mg/kg) | 10 | 2.16 ± 0.18** | 1.49 ± 0.18**▲ | 6.57 ± 0.34*▲ |
| ALB-H (200 mg/kg) | 10 | 3.01 ± 0.22**▲▲ | 2.01 ± 0.23**▲▲ | 6.68 ± 0.32*▲ |
| ALB-M (100 mg/kg) | 10 | 2.27 ± 0.19**▲ | 1.45 ± 0.09**▲▲ | 7.03 ± 0.51▲ |
| ALB-L (50 mg/kg) | 10 | 2.23 ± 0.17**▲ | 1.39 ± 0.18** | 7.02 ± 0.39▲▲ |
3.3. Effect of ALB on Phagocytotic Function of Mononuclear Phagocytic System (MPS)
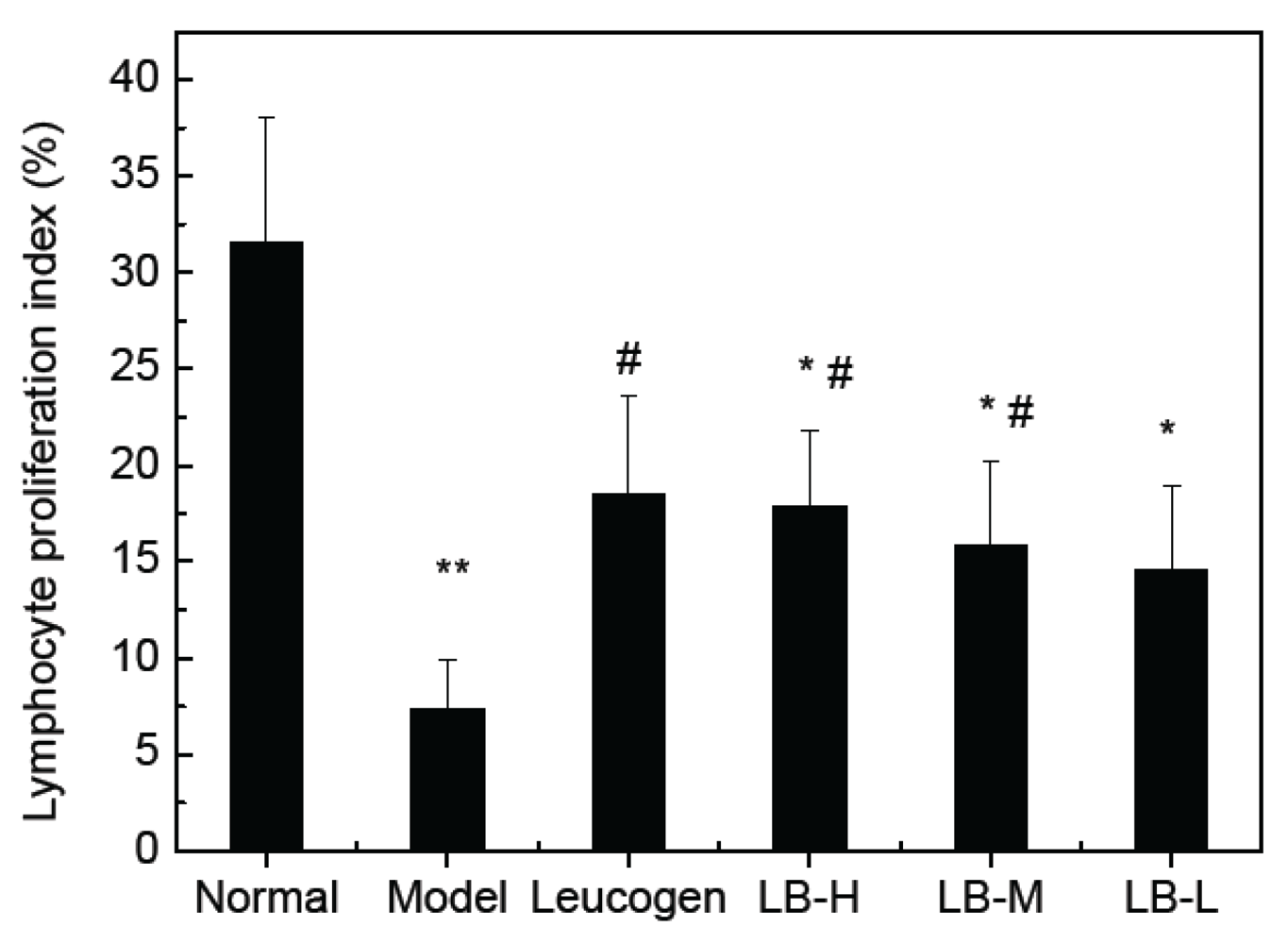
3.4. Effect of ALB on Splenocyte Proliferation

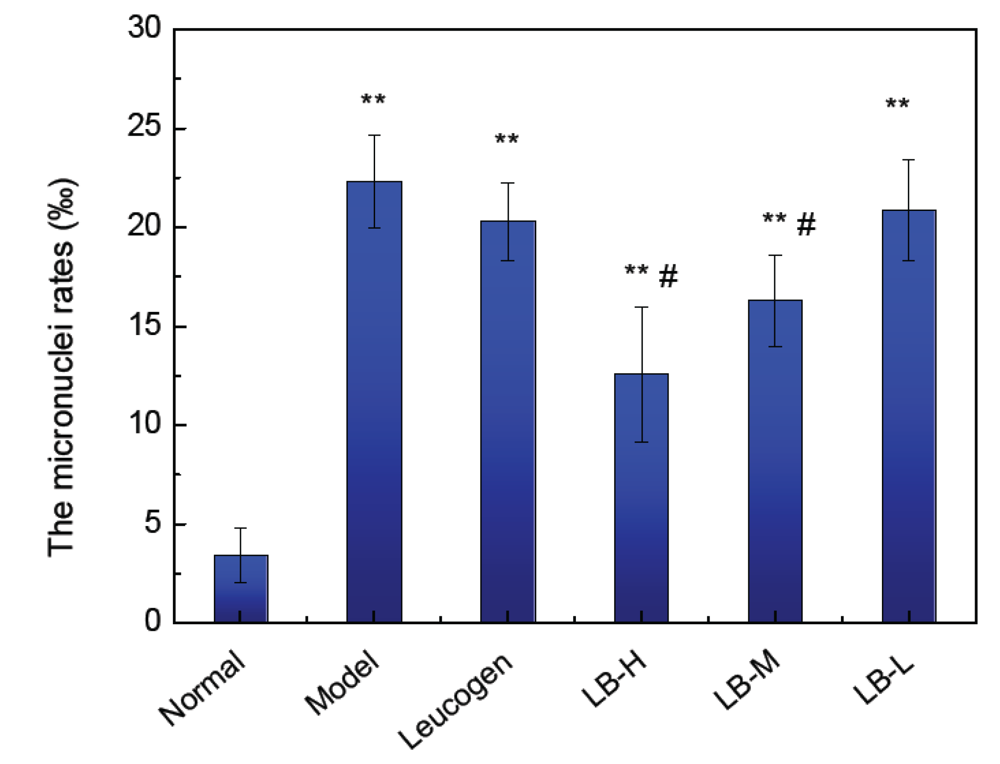
3.5. Effect of ALB on Bone Marrow Micronucleus Formation in Mice

4. Discussion
Conflict of Interest
Acknowledgements
References
- Verma, A.R.; Vijayakumar, M.; Rao, C.V.; Mathela, C.S. In vitro and in vivo antioxidant properties and DNA damage protective activity of green fruit of Ficus glomerata. Food Chem. Toxicol. 2010, 48, 704–709. [Google Scholar] [CrossRef]
- Yao, L.; Wang, Z.; Zhao, H.; Cheng, C.; Fu, X.; Liu, J.; Yang, X. Protective effects of polysaccharides from soybean meal against X-ray radiation induced damage in mouse spleen lymphocytes. Int. J. Mol. Sci. 2011, 12, 8096–8104. [Google Scholar] [CrossRef]
- Li, Y.-R.; Cao, W.; Guo, J.; Miao, S.; Ding, G.-R.; Li, K.-C.; Wang, J.; Guo, G.-Z. Comparative investigations on the protective effects of rhodioside, ciwujianoside-B and astragaloside IV on radiation injuries of the hematopoietic system in mice. Phytother. Res. 2011, 25, 644–653. [Google Scholar]
- Sangsliwan, T.; Haghdoost, S. The nucleotide pool, a target for low-dose gamma-ray-induced oxidative stress. Radiat. Res. 2008, 170, 776–783. [Google Scholar]
- Yang, R.Y.; Pei, X.R.; Wang, J.B.; Zhang, Z.F.; Zhao, H.F.; Li, Q.; Zhao, M.; Li, Y. Protective effect of a marine oligopeptide preparation from Chum Salmon (Oncorhynchus keta) on radiation-induced immune suppression in mice. J. Sci. Food Agr. 2010, 90, 2241–2248. [Google Scholar] [CrossRef]
- Weiss, J.F.; Landauer, M.R. Protection against ionizing radiation by antioxidant nutrients and phytochemicals. Toxicology 2003, 189, 1–20. [Google Scholar] [CrossRef]
- Hosseinimehr, S.M. Foundation review: Trends in the development of radioprotective agents. Drug Discov. Today 2007, 12, 794–805. [Google Scholar] [CrossRef]
- Ignat, I.; Volf, I.; Popa, V.I. A critical review of methods for characterisation of polyphenolic compounds in fruits and vegetables. Food Chem. 2011, 126, 1821–1835. [Google Scholar]
- Zuo, L.-L.; Wang, Z.-Y.; Fan, Z.-L.; Tian, S.-Q.; Liu, J.-R. Evaluation of antioxidant and antiproliferative three actinidia (Actinidia. kolomikta, Actinidia. Actinidia. chinensis) extracts in vitro. Int. J. Mol. Sci. 2012, 13, 5506–5518. [Google Scholar] [CrossRef]
- Szajdek, A.; Borowska, E.J. Bioactive compounds and health-promoting properties of berry fruits: A Review. Plant. Food Hum. Nutr. 2008, 63, 147–156. [Google Scholar] [CrossRef]
- Lehtonen, H.M.; Lehtinen, O.; Suomela, J.P.; Viitanen, M.; Kallio, H. Flavonol glycosides of sea buckthorn (Hippophae. rhamnoides ssp sinensis) and lingonberry (Vaccinium. vitis-idaea) are bioavailable in humans and monoglucuronidated for excretion. J. Agr. Food Chem. 2010, 58, 620–627. [Google Scholar]
- Li, R.; Wang, P.; Guo, Q.-Q.; Wang, Z.-Y. Anthocyanin composition and content of the Vaccinium uliginosum berry. Food Chem. 2011, 125, 116–120. [Google Scholar] [CrossRef]
- Lehtonen, H.M.; Rantala, M.; Suomela, J.P.; Viitanen, M.; Kallio, H. Urinary excretion of the main anthocyanin in lingonberry (vaccinium vitis-idaea), Cyanidin 3-O-Galactoside, and its metabolites. J. Agr. Food Chem. 2009, 57, 4447–4451. [Google Scholar] [CrossRef]
- Fan, Z.-L.; Wang, Z.-Y.; Liu, J.-R. Cold-field fruit extracts exert different antioxidant and antiproliferative activities in vitro. Food Chem. 2011, 129, 402–407. [Google Scholar] [CrossRef]
- Li, X.; Song, H.; Yao, S.; Jia, C.; Yang, Y.; Zhu, W. Quantitative analysis and recovery optimisation of flavonoids and anthocyanins in sugar-making process of sugarcane industry. Food Chem. 2011, 125, 150–157. [Google Scholar] [CrossRef]
- Kalpana, K.B.; Devipriya, N.; Srinivasan, M.; Vishwanathan, P.; Thayalan, K.; Menon, V.P. Evaluating the radioprotective effect of hesperidin in the liver of Swiss albino mice. Eur. J. Pharmacol. 2011, 658, 206–212. [Google Scholar] [CrossRef]
- Wei, X.; Su, F.; Su, X.; Hu, T.; Hu, S. Stereospecific antioxidant effects of ginsenoside Rg3 on oxidative stress induced by cyclophosphamide in mice. Fitoterapia 2012, 83, 636–642. [Google Scholar]
- Patel, P.; Asdaq, S.M.B. Immunomodulatory activity of methanolic fruit extract of Aegle marmelos in experimental animals. Saudi. Pharm. J. 2010, 18, 161–165. [Google Scholar] [CrossRef]
- Srinivasan, M.; Devipriya, N.; Kalpana, K.B.; Menon, V.P. Lycopene: An antioxidant and radioprotector against gamma-radiation-induced cellular damages in cultured human lymphocytes. Toxicology 2009, 262, 43–49. [Google Scholar]
- Landauer, M.R.; Srinivasan, V.; Seed, T.M. Genistein treatment protects mice from ionizing radiation injury. J. Appl. Toxicol. 2003, 23, 379–385. [Google Scholar] [CrossRef]
- Laterveer, L.; Lindley, I.J.D.; Hamilton, M.S.; Willemze, R.; Fibbe, W.E. Interleukin-8 induces rapid mobilization of mobilization of hematopoietic stem-cells with radioprotective capacity and long-term myelolymphoid repopulating ability. Blood 1995, 85, 2269–2275. [Google Scholar]
- Verma, P.; Sharma, P.; Parmar, J.; Sharma, P.; Agrawal, A.; Goyal, P.K. Amelioration of radiation-induced hematological and biochemical alterations in swiss albino mice by panax ginseng extract. Integr. Cancer Ther. 2011, 10, 77–84. [Google Scholar] [CrossRef]
- Tiwari, P.; Kumar, A.; Balakrishnan, S.; Kushwaha, H.S.; Mishra, K.P. Radiation-induced micronucleus formation and DNA damage in human lymphocytes and their prevention by antioxidant thiols. Mutat. Res.-Gen. Tox. En. 2009, 676, 62–68. [Google Scholar] [CrossRef]
- Godevac, D.; Tesevic, V.; Vajs, V.; Milosavljević, S.; Stanković, M. Antioxidant properties of raspberry seed extracts on micronucleus distribution in peripheral blood lymphocytes. Food Chem. Toxicol. 2009, 47, 2853–2859. [Google Scholar] [CrossRef]
- Rithidech, K.N.; Tungjai, M.; Whorton, E.B. Protective effect of apigenin on radiation-induced chromosomal damage in human lymphocytes. Mutat. Res. Gen. Tox. En. 2005, 585, 96–104. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Z.Y.; Zhang, Z.; Wang, X. Purified Auricularia auricular-judae polysaccharide (AAP I-a) prevents oxidative stress in an ageing mouse model. Carbohyd. Polym. 2011, 84, 638–648. [Google Scholar] [CrossRef]
- Seyed Jalal, H. Flavonoids and genomic instability induced by ionizing radiation. Drug Discov. Today 2010, 15, 907–918. [Google Scholar] [CrossRef]
- Srinivasan, M.; Sudheer, A.R.; Rajasekaran, K.N. Effect of curcumin analog on gamma-radiation-induced cellular changes in primary culture of isolated rat hepatocytes in vitro. Environ. Toxicol. Phar. 2007, 24, 98–105. [Google Scholar] [CrossRef]
- Singh, B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Dhakarey, R.; Upadhyay, G.; Singh, H.B. Oxidative DNA damage protective activity, antioxidant and anti-quorum sensing potentials of Moringa oleifera. Food Chem. Toxicol. 2009, 47, 1109–1116. [Google Scholar] [CrossRef]
- Nzaramba, M.N.; Reddivari, L.; Bamberg, J.B.; Creighton Miller, J. Antiproliferative activity and cytotoxicity of Solanum jamesii tuber extracts on human colon and prostate cancer cells in vitro. J. Agric. Food Chem. 2009, 57, 8308–8315. [Google Scholar]
- Chen, J.-R.; Yang, Z.-Q.; Hu, T.-J.; Yan, Z.-T.; Niu, T.-X.; Wang, L.; Cui, D.-A.; Wang, M. Immunomodulatory activity in vitro and in vivo of polysaccharide from Potentilla anserina. Fitoterapia 2010, 81, 1117–1124. [Google Scholar] [CrossRef]
- Bent, H. The biochemistry and medical significance of the flavonoids. Pharmacol. Therapeut. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Shih, P.-H.; Chan, Y.-C.; Liao, J.-W.; Wang, M.-F.; Yen, G.-C. Antioxidant and cognitive promotion effects of anthocyanin-rich mulberry (Morus atropurpurea L.) on senescence-accelerated mice and prevention of Alzheimer’s disease. J. Nutr. Biochem. 2010, 21, 598–605. [Google Scholar] [CrossRef]
- Youdim, K.A.; Martin, A.; Joseph, J.A. Incorporation of the elderberry anthocyanins by endothelial cells increases protection against oxidative stress. Free Radical Bio. Med. 2000, 29, 51–60. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fan, Z.-L.; Wang, Z.-Y.; Zuo, L.-L.; Tian, S.-Q. Protective Effect of Anthocyanins from Lingonberry on Radiation-induced Damages. Int. J. Environ. Res. Public Health 2012, 9, 4732-4743. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph9124732
Fan Z-L, Wang Z-Y, Zuo L-L, Tian S-Q. Protective Effect of Anthocyanins from Lingonberry on Radiation-induced Damages. International Journal of Environmental Research and Public Health. 2012; 9(12):4732-4743. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph9124732
Chicago/Turabian StyleFan, Zi-Luan, Zhen-Yu Wang, Li-Li Zuo, and Shuang-Qi Tian. 2012. "Protective Effect of Anthocyanins from Lingonberry on Radiation-induced Damages" International Journal of Environmental Research and Public Health 9, no. 12: 4732-4743. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph9124732