Potential Role of CCN Proteins in Breast Cancer: Therapeutic Advances and Perspectives

 ,
,  ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Methods
3. Nomenclature
4. CCN Gene Family
5. Molecular Makeup of CCN Proteins
6. Potential Role of CCN Proteins in Tumorigenesis and Breast Cancer
6.1. CCN1: CYR61
6.2. CCN2: CTGF
6.3. CCN3: NOV
6.4. CCN4: WISP-1
6.5. CCN5: WISP-2
6.6. CCN6: WISP-3
7. MicroRNA Regulation of CCN Gene in Breast Cancer
8. Pharmacological Advances in Treating Breast Cancer by Targeting CCN Proteins
9. Future Perspectives and Limitations
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Naik, A.V.; Sellappan, K. Assessment of Genotoxic potential of Annonacin and Annona muricata L. extracts on human breast cancer (MCF-7) cells. Adv. Tradit. Med. 2021, 21, 779–789. [Google Scholar] [CrossRef]
- Kim, H.; Son, S.; Shin, I. Role of the CCN protein family in cancer. BMB Rep. 2018, 51, 486–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnott, J.A.; Lambi, A.G.; Mundy, C.; Hendesi, H.; Pixley, R.A.; Owen, T.A.; Safadi, F.F.; Popoff, S.N. The role of connective tissue growth factor (CTGF/CCN2) in skeletogenesis. Crit. Rev. Eukaryot. Gene Expr. 2011, 21, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Zucca-Matthes, G.; Urban, C.; Vallejo, A. Anatomy of the nipple and breast ducts. Gland Surg. 2016, 5, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.H.; Lin, F.L.; Hou, S.M.; Liu, J.F. Cyr61 promotes epithelial-mesenchymal transition and tumor metastasis of osteosarcoma by Raf-1/MEK/ERK/Elk-1/TWIST-1 signaling pathway. Mol. Cancer 2014, 13, 236. [Google Scholar] [CrossRef] [Green Version]
- Chien, W.; O’Kelly, J.; Lu, D.; Leiter, A.; Sohn, J.; Yin, D.; Karlan, B.; Vadgama, J.; Lyons, K.M.; Koeffler, H.P. Expression of connective tissue growth factor (CTGF/CCN2) in breast cancer cells is associated with increased migration and angiogenesis. Int. J. Oncol. 2011, 38, 1741–1747. [Google Scholar] [CrossRef] [Green Version]
- Barreto, S.C.; Ray, A.; Ag Edgar, P. Biological characteristics of CCN proteins in tumor development. J. BUON 2016, 21, 1359–1367. [Google Scholar] [PubMed]
- Chiang, K.C.; Yeh, C.N.; Chung, L.C.; Feng, T.H.; Sun, C.C.; Chen, M.F.; Jan, Y.Y.; Yeh, T.S.; Chen, S.C.; Juang, H.H. WNT-1 inducible signaling pathway protein-1 enhances growth and tumorigenesis in human breast cancer. Sci. Rep. 2015, 5, 8686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ye, L.; Owen, S.; Weeks, H.P.; Zhang, Z.; Jiang, W.G. Emerging role of CCN family proteins in tumorigenesis and cancer metastasis (Review). Int. J. Mol. Med. 2015, 36, 1451–1463. [Google Scholar] [CrossRef] [Green Version]
- Ouellet, V.; Tiedemann, K.; Mourskaia, A.; Fong, J.E.; Tran-Thanh, D.; Amir, E.; Clemons, M.; Perbal, B.; Komarova, S.V.; Siegel, P.M. CCN3 impairs osteoblast and stimulates osteoclast differentiation to favor breast cancer metastasis to bone. Am. J. Pathol. 2011, 178, 2377–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherbet, G.V. Metastasis Promoter S100A4 is a Potential Molecular Therapeutic Target. Cancer Genom. Proteom. 2006, 3, 203–216. [Google Scholar]
- Jun, J.I.; Lau, L.F. The matricellular protein CCN1 induces fibroblast senescence and restricts fibrosis in cutaneous wound healing. Nat. Cell Biol. 2010, 12, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.K.; Banerjee, S. CCN5/WISP-2: A micromanager of breast cancer progression. J. Cell Commun. Signal. 2012, 6, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Pal, A.; Kleer, C.G. On how CCN6 suppresses breast cancer growth and invasion. J. Cell Commun. Signal. 2012, 6, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- PubMed. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/ (accessed on 15 October 2021).
- Boumediene, A.; Oblet, C.; Oruc, Z.; Duchez, S.; Morelle, W.; Huynh, A.; Pourrat, J.; Aldigier, J.C.; Cogné, M. Gammopathy with IgA mesangial deposition provides a monoclonal model of IgA nephritogenicity and offers new insights into its molecular mechanisms. Nephrol. Dial. Transplant. 2011, 26, 3930–3937. [Google Scholar] [CrossRef] [Green Version]
- Inkscape Draw Freel; Developers IW: Los Angeles, CA, USA, 2021.
- Kleer, C.G. Dual roles of CCN proteins in breast cancer progression. J. Cell Commun. Signal. 2016, 10, 217–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kular, L.; Pakradouni, J.; Kitabgi, P.; Laurent, M.; Martinerie, C. The CCN family: A new class of inflammation modulators? Biochimie 2011, 93, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.I.; Lau, L.F. Taking aim at the extracellular matrix: CCN proteins as emerging therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 945–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.C.; Lau, L.F. Functions and mechanisms of action of CCN matricellular proteins. Int. J. Biochem. Cell Biol. 2009, 41, 771–783. [Google Scholar] [CrossRef] [Green Version]
- Home|HUGO Gene Nomenclature Committee. Available online: https://www.genenames.org/ (accessed on 15 October 2021).
- Jia, Q.; Xu, B.; Zhang, Y.; Ali, A.; Liao, X. CCN Family Proteins in Cancer: Insight Into Their Structures and Coordination Role in Tumor Microenvironment. Front. Genet. 2021, 12, 649387. [Google Scholar] [CrossRef]
- Wang, S.; Chong, Z.Z.; Shang, Y.C.; Maiese, K. WISP1 (CCN4) autoregulates its expression and nuclear trafficking of β-catenin during oxidant stress with limited effects upon neuronal autophagy. Curr. Neurovascular Res. 2012, 9, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Gray, M.R.; Oliveira, B.E.; Koch, M.; Castellot, J.J., Jr. CCN5 expression in mammals: I. Embryonic and fetal tissues of mouse and human. J. Cell Commun. Signal. 2007, 1, 127–143. [Google Scholar] [CrossRef] [Green Version]
- Kubota, S.; Takigawa, M. The CCN family acting throughout the body: Recent research developments. Biomol. Concepts 2013, 4, 477–494. [Google Scholar] [CrossRef]
- Planey, J.A.A.; Kathleen, D.; Sonia, L. CCN Family: Matricellular Proteins in Cartilage and Bone Development. Compos. Funct. Extracell. Matrix Hum. Body 2016, 161. [Google Scholar] [CrossRef] [Green Version]
- Russo, J.W.; Castellot, J.J. CCN5: Biology and pathophysiology. J. Cell Commun. Signal. 2010, 4, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Zuo, G.W.; Kohls, C.D.; He, B.C.; Chen, L.; Zhang, W.; Shi, Q.; Zhang, B.Q.; Kang, Q.; Luo, J.; Luo, X.; et al. The CCN proteins: Important signaling mediators in stem cell differentiation and tumorigenesis. Histol. Histopathol. 2010, 25, 795–806. [Google Scholar] [CrossRef]
- Takigawa, M. The CCN Proteins: An Overview. CCN Proteins 2017, 1489, 1–8. [Google Scholar] [CrossRef]
- Perbal, B. CCN proteins are part of a multilayer complex system: A working model. J. Cell Commun. Signal. 2019, 13, 437–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, L.F. Cell surface receptors for CCN proteins. J. Cell Commun. Signal. 2016, 10, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, Y.D.; Xiao, Y.L.; Li, M.H.; Wang, Y.; Kan, X.; Li, Q.Y.; Lu, J.G.; Jin, D.J. Cyr61/CCN1 overexpression induces epithelial-mesenchymal transition leading to laryngeal tumor invasion and metastasis and poor prognosis. Asian Pac. J. Cancer Prev. APJCP 2015, 16, 2659–2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinoza, I.; Menendez, J.A.; Kvp, C.M.; Lupu, R. CCN1 promotes vascular endothelial growth factor secretion through αvβ3 integrin receptors in breast cancer. J. Cell Commun. Signal. 2014, 8, 23–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Kelly, J.; Chung, A.; Lemp, N.; Chumakova, K.; Yin, D.; Wang, H.J.; Said, J.; Gui, D.; Miller, C.W.; Karlan, B.Y.; et al. Functional domains of CCN1 (Cyr61) regulate breast cancer progression. Int. J. Oncol. 2008, 33, 59–67. [Google Scholar] [CrossRef]
- Ren, W.; Sun, X.; Wang, K.; Feng, H.; Liu, Y.; Fei, C.; Wan, S.; Wang, W.; Luo, J.; Shi, Q.; et al. BMP9 inhibits the bone metastasis of breast cancer cells by downregulating CCN2 (connective tissue growth factor, CTGF) expression. Mol. Biol. Rep. 2014, 41, 1373–1383. [Google Scholar] [CrossRef] [PubMed]
- McCallum, L.; Price, S.; Planque, N.; Perbal, B.; Pierce, A.; Whetton, A.D.; Irvine, A.E. A novel mechanism for BCR-ABL action: Stimulated secretion of CCN3 is involved in growth and differentiation regulation. Blood 2006, 108, 1716–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, I.; Banerjee, S.; Mehta, S.; De, A.; Majumder, M.; Mayo, M.S.; Kambhampati, S.; Campbell, D.R.; Banerjee, S.K. Cysteine-rich 61-connective tissue growth factor-nephroblastoma-overexpressed 5 (CCN5)/Wnt-1-induced signaling protein-2 (WISP-2) regulates microRNA-10b via hypoxia-inducible factor-1α-TWIST signaling networks in human breast cancer cells. J. Biol. Chem. 2011, 286, 43475–43485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrand, N.; Stragier, E.; Redeuilh, G.; Sabbah, M. Glucocorticoids induce CCN5/WISP-2 expression and attenuate invasion in oestrogen receptor-negative human breast cancer cells. Biochem. J. 2012, 447, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhang, Y.; Varambally, S.; Chinnaiyan, A.M.; Banerjee, M.; Merajver, S.D.; Kleer, C.G. Inhibition of CCN6 (Wnt-1-induced signaling protein 3) down-regulates E-cadherin in the breast epithelium through induction of snail and ZEB1. Am. J. Pathol. 2008, 172, 893–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, A.; Huang, W.; Li, X.; Toy, K.A.; Nikolovska-Coleska, Z.; Kleer, C.G. CCN6 modulates BMP signaling via the Smad-independent TAK1/p38 pathway, acting to suppress metastasis of breast cancer. Cancer Res. 2012, 72, 4818–4828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wu, G.; Dai, H. The matricellular protein CCN1 regulates TNF-α induced vascular endothelial cell apoptosis. Cell Biol. Int. 2016, 40, 1–6. [Google Scholar] [CrossRef]
- Jeong, D.; Heo, S.; Sung Ahn, T.; Lee, S.; Park, S.; Kim, H.; Park, D.; Byung Bae, S.; Lee, S.S.; Soo Lee, M.; et al. Cyr61 expression is associated with prognosis in patients with colorectal cancer. BMC Cancer 2014, 14, 164. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhang, H.; Song, X.; Yang, Q. Metastatic heterogeneity of breast cancer: Molecular mechanism and potential therapeutic targets. Semin. Cancer Biol. 2020, 60, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yan, C.; Wu, W.; He, S.; Liu, M.; Liu, J.; Yang, X.; Ma, J.; Lu, Y.; Jia, L. RU486 Metabolite Inhibits CCN1/Cyr61 Secretion by MDA-MB-231-Endothelial Adhesion. Front. Pharmacol. 2019, 10, 1296. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Won, J.H.; Cheng, N.; Lau, L.F. The matricellular protein CCN1 in tissue injury repair. J. Cell Commun. Signal. 2018, 12, 273–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall-Glenn, F.; Lyons, K.M. Roles for CCN2 in normal physiological processes. Cell. Mol. Life Sci. CMLS 2011, 68, 3209–3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, F.; Carter, D.E.; Leask, A. Mechanical tension increases CCN2/CTGF expression and proliferation in gingival fibroblasts via a TGFβ-dependent mechanism. PLoS ONE 2011, 6, e19756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, D.P.; Lappano, R.; Albanito, L.; Madeo, A.; Maggiolini, M.; Picard, D. Estrogenic GPR30 signalling induces proliferation and migration of breast cancer cells through CTGF. EMBO J. 2009, 28, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.C.; Cheng, H.C.; Tang, C.H. CCN3 promotes prostate cancer bone metastasis by modulating the tumor-bone microenvironment through RANKL-dependent pathway. Carcinogenesis 2013, 34, 1669–1679. [Google Scholar] [CrossRef] [Green Version]
- Dobson, J.R.; Taipaleenmäki, H.; Hu, Y.J.; Hong, D.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B.; Pratap, J. hsa-mir-30c promotes the invasive phenotype of metastatic breast cancer cells by targeting NOV/CCN3. Cancer Cell Int. 2014, 14, 73. [Google Scholar] [CrossRef] [Green Version]
- Sabbah, M.; Prunier, C.; Ferrand, N.; Megalophonos, V.; Lambein, K.; De Wever, O.; Nazaret, N.; Lachuer, J.; Dumont, S.; Redeuilh, G. CCN5, a novel transcriptional repressor of the transforming growth factor β signaling pathway. Mol. Cell. Biol. 2011, 31, 1459–1469. [Google Scholar] [CrossRef] [Green Version]
- Ferrand, N.; Gnanapragasam, A.; Dorothee, G.; Redeuilh, G.; Larsen, A.K.; Sabbah, M. Loss of WISP2/CCN5 in estrogen-dependent MCF7 human breast cancer cells promotes a stem-like cell phenotype. PLoS ONE 2014, 9, e87878. [Google Scholar] [CrossRef]
- Tran, M.N.; Kleer, C.G. Matricellular CCN6 (WISP3) protein: A tumor suppressor for mammary metaplastic carcinomas. J. Cell Commun. Signal. 2018, 12, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeger, H.; Perbal, B. CCN family of proteins: Critical modulators of the tumor cell microenvironment. J. Cell Commun. Signal. 2016, 10, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Lorenzatti, G.; Huang, W.; Pal, A.; Cabanillas, A.M.; Kleer, C.G. CCN6 (WISP3) decreases ZEB1-mediated EMT and invasion by attenuation of IGF-1 receptor signaling in breast cancer. J. Cell Sci. 2011, 124, 1752–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petri, B.J.; Klinge, C.M. Regulation of breast cancer metastasis signaling by miRNAs. Cancer Metastasis Rev. 2020, 39, 837–886. [Google Scholar] [PubMed]
- O’Connor, L.M.; Sharon, F.M.; William, M.G.; Darran, P. miRNA Dysregulation in Breast Cancer. Cancer Res. 2013, 73, 6554–6562. [Google Scholar] [CrossRef] [Green Version]
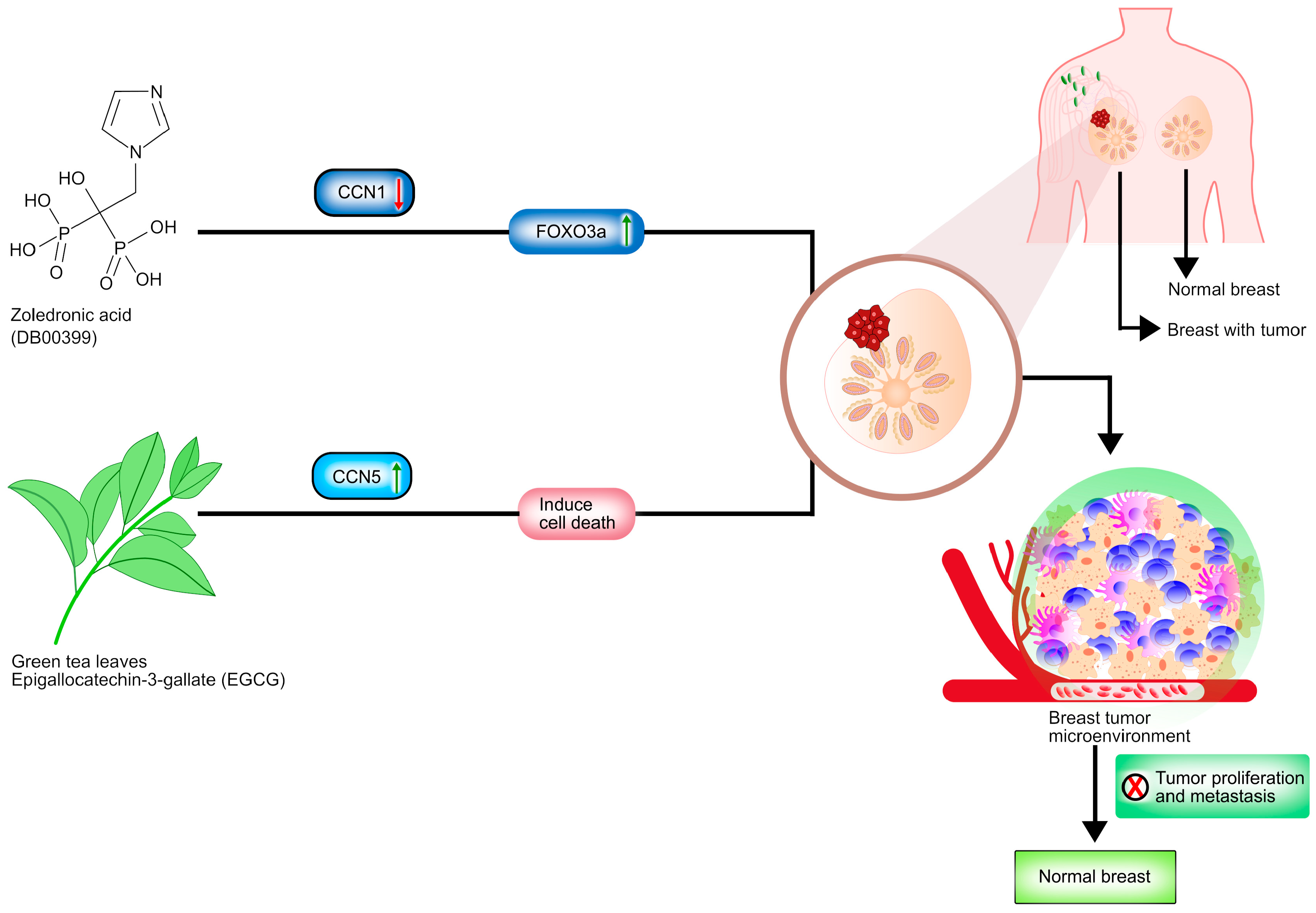
- Espinoza, I.; Liu, H.; Busby, R.; Lupu, R. CCN1, a candidate target for zoledronic acid treatment in breast cancer. Mol. Cancer Ther. 2011, 10, 732–741. [Google Scholar] [CrossRef] [Green Version]
- Zoledronic Acid. Available online: https://go.drugbank.com/drugs/DB00399 (accessed on 15 October 2021).
- Jiang, Y.; Zou, L.; Lu, W.Q.; Zhang, Y.; Shen, A.G. Foxo3a expression is a prognostic marker in breast cancer. PLoS ONE 2013, 8, e70746. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Hu, X. Dosing of zoledronic acid with its anti-tumor effects in breast cancer. J. Bone Oncol. 2015, 4, 98–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.; Haque, I.; Ray, P.; Ghosh, A.; Dutta, D.; Quadir, M.; De, A.; Gunewardena, S.; Chatterjee, I.; Banerjee, S.; et al. CCN5 activation by free or encapsulated EGCG is required to render triple-negative breast cancer cell viability and tumor progression. Pharmacol. Res. Perspect. 2021, 9, e00753. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.S.; Spicer, D.V.; Hawes, D.; Tseng, C.C.; Yang, C.S.; Pike, M.C.; Wu, A.H. Biological effects of green tea capsule supplementation in pre-surgery postmenopausal breast cancer patients. Front. Oncol. 2013, 3, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, Y.; Rayburn, E.R.; Hill, D.L.; Rinehart, J.J.; Zhang, R. Dexamethasone as a chemosensitizer for breast cancer chemotherapy: Potentiation of the antitumor activity of adriamycin, modulation of cytokine expression, and pharmacokinetics. Int. J. Oncol. 2007, 30, 947–953. [Google Scholar] [CrossRef]
- Pang, J.M.; Huang, Y.C.; Sun, S.P.; Pan, Y.R.; Shen, C.Y.; Kao, M.C.; Wang, R.H.; Wang, L.H.; Lin, K.T. Effects of synthetic glucocorticoids on breast cancer progression. Steroids 2020, 164, 108738. [Google Scholar] [CrossRef]
- Yu, L.; Zhao, J.; Gao, L. Predicting Potential Drugs for Breast Cancer based on miRNA and Tissue Specificity. Int. J. Biol. Sci. 2018, 14, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, D.P.; de Farias, G.C.; de Sousa, E.B.; de Mattos Coelho-Aguiar, J.; Lobo, J.C.; Casado, P.L.; Duarte, M.E.; Abreu, J.G., Jr. New strategy to control cell migration and metastasis regulated by CCN2/CTGF. Cancer Cell Int. 2014, 14, 61. [Google Scholar] [CrossRef] [Green Version]
- Abd El Kader, T.; Kubota, S.; Janune, D.; Nishida, T.; Hattori, T.; Aoyama, E.; Perbal, B.; Kuboki, T.; Takigawa, M. Anti-fibrotic effect of CCN3 accompanied by altered gene expression profile of the CCN family. J. Cell Commun. Signal. 2013, 7, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berschneider, B.; Königshoff, M. WNT1 inducible signaling pathway protein 1 (WISP1): A novel mediator linking development and disease. Int. J. Biochem. Cell Biol. 2011, 43, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ji, X.; Liu, W.; Huang, B.; Luo, W.; et al. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018, 5, 77–106. [Google Scholar] [CrossRef]
- Dixon, J.M. Endocrine resistance in breast cancer. New J. Sci. 2014, 2014, 390618. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
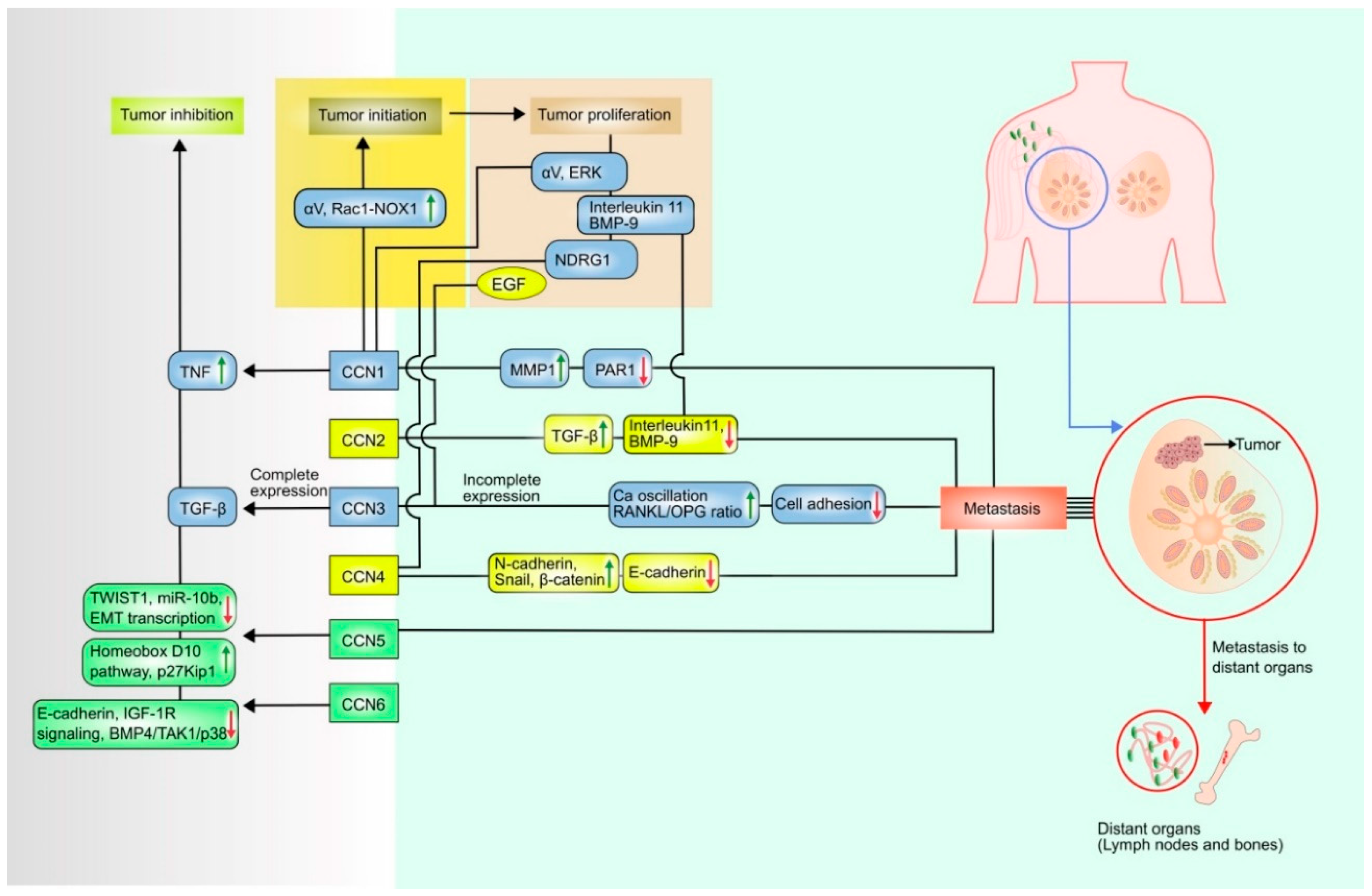
| Proteins | Role | Upregulated Factors | Downregulated Factors |
|---|---|---|---|
| CCN1 | Initiation | αV integrin proteins, [34] | N/A |
| Proliferation | αV integrin proteins [34,35] | N/A | |
| Metastasis | MMP-1 [36] | PAR 1 [37] | |
| Tumor Inhibition | TNF [13] | N/A | |
| CCN2 | Initiation | N/A | N/A |
| Proliferation | N/A | Binding with interleukin 11, BMP-9 [38] | |
| Metastasis | TGF-β, ERK [3] | Binding with interleukin 11, BMP-9 [38] | |
| Tumor inhibition | N/A | N/A | |
| CCN3 | Initiation | N/A | N/A |
| Proliferation | N/A | ||
| Metastasis | Ca oscillation, Receptor activator of nuclear factor kappa-Β ligand (RANKL)/Osteo-protegrin (OPG) [11] | N/A | |
| Tumor inhibition | TGF-β [39] | N/A | |
| CCN4 | Initiation | N/A | N/A |
| Proliferation | N/A | N-myc downstream regulated 1 (NDRG) [9] | |
| Metastasis | N-cadherin, snail, β-catenin [8,9] | E-cadherin [8,9] | |
| Tumor inhibition | N/A | N/A | |
| CCN5 | Initiation | N/A | N/A |
| Proliferation | N/A | N/A | |
| Metastasis | N/A | N/A | |
| Tumor inhibition | Homeobox D10 pathway, p27Kip1 [40] | TWIST 1, miR-10b [40], Epithelial mesenchymal transition (EMT) transcription [41] | |
| CCN6 | Initiation | N/A | N/A |
| Proliferation | N/A | N/A | |
| Metastasis | N/A | N/A | |
| Tumor inhibition | N/A | E-cadherin expression, IGF-1R Signaling [42], BMP4/TAK1/p38 [43] |
| Gene | MicroRNA | Role in Breast Cancer |
|---|---|---|
| CCN3 | miR-30c [53] | “Complete” expression can inhibit breast cancer progression [12] whereas, overexpression leads to aggressive behavior of the cancer [11] |
| CCN2 | miR-124-3p, miR-18a-5p, miR-145-5p [59] | Can cause migration and angiogenesis of breast cancer cells [7] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, K.A.; Hasib, T.A.; Paul, S.K.; Saddam, M.; Mimi, A.; Saikat, A.S.M.; Faruque, H.A.; Rahman, M.A.; Uddin, M.J.; Kim, B. Potential Role of CCN Proteins in Breast Cancer: Therapeutic Advances and Perspectives. Curr. Oncol. 2021, 28, 4972-4985. https://0-doi-org.brum.beds.ac.uk/10.3390/curroncol28060417
Ahmed KA, Hasib TA, Paul SK, Saddam M, Mimi A, Saikat ASM, Faruque HA, Rahman MA, Uddin MJ, Kim B. Potential Role of CCN Proteins in Breast Cancer: Therapeutic Advances and Perspectives. Current Oncology. 2021; 28(6):4972-4985. https://0-doi-org.brum.beds.ac.uk/10.3390/curroncol28060417
Chicago/Turabian StyleAhmed, Kazi Ahsan, Tasnin Al Hasib, Shamrat Kumar Paul, Md. Saddam, Afsana Mimi, Abu Saim Mohammad Saikat, Hasan Al Faruque, Md. Ataur Rahman, Md. Jamal Uddin, and Bonglee Kim. 2021. "Potential Role of CCN Proteins in Breast Cancer: Therapeutic Advances and Perspectives" Current Oncology 28, no. 6: 4972-4985. https://0-doi-org.brum.beds.ac.uk/10.3390/curroncol28060417