Investigation and Functional Enrichment Analysis of the Human Host Interaction Network with Common Gram-Negative Respiratory Pathogens Predicts Possible Association with Lung Adenocarcinoma

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
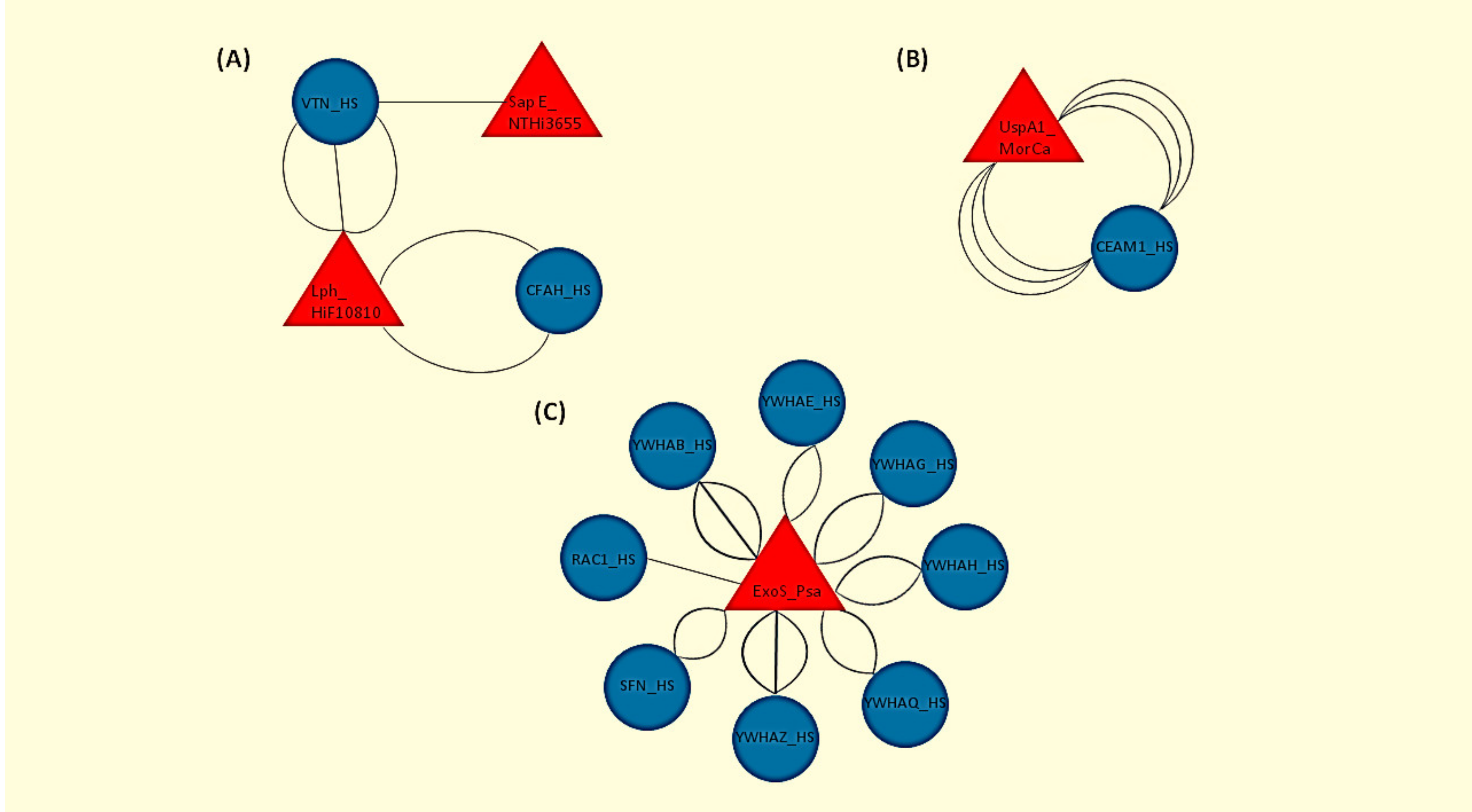
3.1. Identification of the Host–Pathogen Interactomes
3.2. ToppFun FEA Results for the CEACAM1, CFH, VTN, RAC1, YWHAB, YWHAE, YWHAG, YWHAH, YWHAQ, YWHAZ and SFN Genes, Relative to BP GOs, CC GOs and Diseases
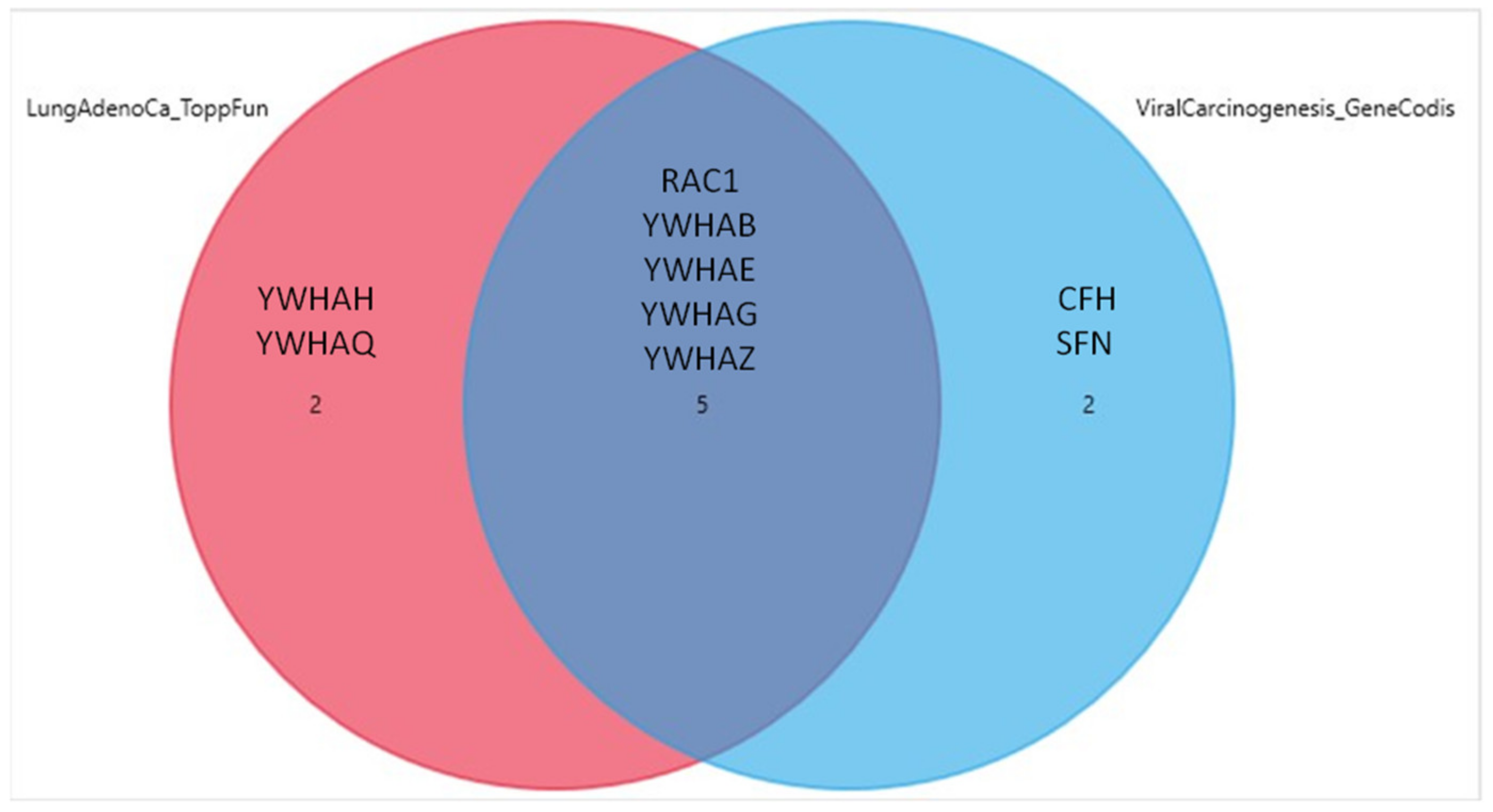
3.3. GeneCodis FEA Results for the CEACAM1, CFH, VTN, RAC1, YWHAB, YWHAE, YWHAG, YWHAH, YWHAQ, YWHAZ and SFN Genes Relative to BP GOs, CC GOs and KEGG Pathways
4. Discussion
4.1. Biological Interpretation of the Human-Gram (−) Pathogens Interactome
4.2. Gene Involvement in Apoptotic Pathways, Cellular Junctions, Cell Cycle, Carcinogenesis and Lung Adenocarcinoma: FEA Interpretation
4.3. Novelty, Weaknesses and Future Directions of Our Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sriram, K.B.; Cox, A.J.; Clancy, R.L.; Slack, M.P.E.; Cripps, A.W. Nontypeable Haemophilus influenzae and chronic obstructive pulmonary disease: A review for clinicians. Crit. Rev. Microbiol. 2018, 44, 125–142. [Google Scholar] [CrossRef] [PubMed]
- Vickery, T.W.; Ramakrishnan, V.R. Bacterial Pathogens and the Microbiome. Otolaryngol. Clin. N. Am. 2017, 50, 29–47. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, H.; Wallis, S.; Coatesworth, A.P. Acute otitis media. Postgrad. Med. 2015, 127, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Fujitani, S.; Sun, H.-Y.; Yu, V.L.; Weingarten, J.A. Pneumonia due to Pseudomonas aeruginosa: Part I: Epidemiology, Clinical Diagnosis, and Source. Chest 2011, 139, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Riesbeck, K. Complement evasion by the human respiratory tract pathogens Haemophilus influenzae and Moraxella catarrhalis. FEBS Lett. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, R.; Economopoulou, A.; Dias, J.G.; Bancroft, E.; Ramliden, M.; Celentano, L.P.; Steindl, G.; Martiny, D.; Grammens, T.; Georgieva, T.; et al. Epidemiology of invasive Haemophilus influenzae disease, Europe, 2007–2014. Emerg. Infect. Dis. 2017, 23, 396–404. [Google Scholar] [CrossRef] [Green Version]
- Soeters, H.M.; Blain, A.; Pondo, T.; Doman, B.; Farley, M.M.; Harrison, L.H.; Lynfield, R.; Miller, L.; Petit, S.; Reingold, A.; et al. Current Epidemiology and Trends in Invasive Haemophilus influenzae Disease—United States, 2009–2015. Clin. Infect. Dis. 2018, 67, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Blakeway, L.V.; Tan, A.; Peak, I.R.; Seib, K.L. Virulence determinants of Moraxella catarrhalis: Distribution and considerations for vaccine development. Microbiology 2017, 163, 1371–1384. [Google Scholar] [CrossRef]
- Murphy, T.F.; Brauer, A.L.; Grant, B.J.B.; Sethi, S. Moraxella catarrhalis in chronic obstructive pulmonary disease: Burden of disease and immune response. Am. J. Respir. Crit. Care Med. 2005, 172, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.T.; Mörgelin, M.; Forsgren, A.; Riesbeck, K. Haemophilus influenzae Survival during Complement-Mediated Attacks Is Promoted by Moraxella catarrhalis Outer Membrane Vesicles. J. Infect. Dis. 2007, 195, 1661–1670. [Google Scholar] [CrossRef] [Green Version]
- Rodrigo-Troyano, A.; Sibila, O. The respiratory threat posed by multidrug resistant Gram-negative bacteria. Respirology 2017, 22, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Aksamit, T.; Aliberti, S.; De Soyza, A.; Elborn, J.; Goeminne, P.; Hill, A.T.; Menendez, R.; Polverino, E. Challenges in managing Pseudomonas aeruginosa in non-cystic fibrosis bronchiectasis. Respir. Med. 2016, 117, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallström, T.; Blom, A.M.; Zipfel, P.F.; Riesbeck, K. Nontypeable Haemophilus influenzae Protein E Binds Vitronectin and Is Important for Serum Resistance. J. Immunol. 2009, 183, 2593–2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleury, C.; Su, Y.-C.; Hallström, T.; Sandblad, L.; Zipfel, P.F.; Riesbeck, K. Identification of a Haemophilus influenzae Factor H–Binding Lipoprotein Involved in Serum Resistance. J. Immunol. 2014, 192, 5913–5923. [Google Scholar] [CrossRef] [PubMed]
- Spaniol, V.; Heiniger, N.; Troller, R.; Aebi, C. Outer membrane protein UspA1 and lipooligosaccharide are involved in invasion of human epithelial cells by Moraxella catarrhalis. Microbes Infect. 2008, 10, 3–11. [Google Scholar] [CrossRef]
- Würtele, M.; Wolf, E.; Pederson, K.J.; Buchwald, G.; Ahmadian, M.R.; Barbieri, J.T.; Wittinghofer, A. How the Pseudomonas aeruginosa ExoS toxin downregulates Rac. Nat. Struct. Mol. Biol. 2001, 8, 23–26. [Google Scholar] [CrossRef]
- Paulsson, M.; Riesbeck, K. How bacteria hack the matrix and dodge the bullets of immunity. Eur. Respir. Rev. 2018, 27, 1–8. [Google Scholar] [CrossRef]
- Ammari, M.G.; Gresham, C.R.; McCarthy, F.M.; Nanduri, B. HPIDB 2.0: A curated database for host-pathogen interactions. Database 2016, 2016, baw103. [Google Scholar] [CrossRef]
- Bateman, A. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef]
- Tabas-Madrid, D.; Nogales-Cadenas, R.; Pascual-Montano, A. GeneCodis3: A non-redundant and modular enrichment analysis tool for functional genomics. Nucleic Acids Res. 2012, 40, W478–W483. [Google Scholar] [CrossRef] [PubMed]
- Al-Jubair, T.; Mukherjee, O.; Oosterhuis, S.; Singh, B.; Su, Y.-C.; Fleury, C.; Blom, A.M.; Törnroth-Horsefield, S.; Riesbeck, K. Haemophilus influenzae Type f Hijacks Vitronectin Using Protein H To Resist Host Innate Immunity and Adhere to Pulmonary Epithelial Cells. J. Immunol. 2015, 195, 5688–5695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conners, R.; Hill, D.J.; Borodina, E.; Agnew, C.; Daniell, S.J.; Burton, N.M.; Sessions, R.B.; Clarke, A.R.; Catto, L.; Lammie, D.; et al. The Moraxella adhesin UspA1 binds to its human CEACAM1 receptor by a deformable trimeric coiled-coil. EMBO J. 2008, 27, 1779–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, D.J.; Edwards, A.M.; Rowe, H.A.; Virji, M. Carcinoembryonic antigen-related cell adhesion molecule (CEACAM)-binding recombinant polypeptide confers protection against infection by respiratory and urogenital pathogens. Mol. Microbiol. 2005, 55, 1515–1527. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, M.L.; Trollér, U.; Hallberg, B. 14-3-3 proteins are required for the inhibition of Ras by exoenzyme S. Biochem. J. 2000, 349, 697–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriksson, M.L.; Francis, M.S.; Peden, A.; Aili, M.; Stefansson, K.; Palmer, R.; Aitken, A.; Hallberg, B. A nonphosphorylated 14-3-3 binding motif on exoenzyme S that is functional in vivo. Eur. J. Biochem. 2002, 269, 4921–4929. [Google Scholar] [CrossRef]
- Timmerman, I.; Heemskerk, N.; Kroon, J.; Schaefer, A.; Van Rijssel, J.; Hoogenboezem, M.; Van Unen, J.; Goedhart, J.; Gadella, T.W.J.; Yin, T.; et al. A local VE-cadherin and Trio-based signaling complex stabilizes endothelial junctions through Rac1. J. Cell Sci. 2015, 128, 3041–3054. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, J.S.; Hansen, M.D.; Nelson, W.J. Spatio-Temporal Regulation of Rac1 Localization and Lamellipodia Dynamics during Epithelial Cell-Cell Adhesion. Dev. Cell 2002, 3, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Wojciak-Stothard, B.; Torondel, B.; Zhao, L.; Renné, T.; Leiper, J.M. Modulation of Rac1 Activity by ADMA/DDAH Regulates Pulmonary Endothelial Barrier Function. Mol. Biol. Cell. 2009, 20, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Smyth, J.W.; Zhang, S.-S.; Sanchez, J.M.; Lamouille, S.; Vogan, J.M.; Hesketh, G.G.; Hong, T.; Tomaselli, G.F.; Shaw, R.M. A 14-3-3 Mode-1 Binding Motif Initiates Gap Junction Internalization During Acute Cardiac Ischemia. Traffic 2014, 15, 684–699. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.A.; Takemoto, L.J.; Takemoto, D.J. Inhibition of gap junction activity through the release of the C1B domain of protein kinase Cgamma (PKCgamma) from 14-3-3: Identification of PKCgamma-binding sites. J. Biol. Chem. 2004, 279, 52714–52725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.C.; Rodriguez, L.G.; Guan, J.L. Identification of a novel interaction between integrin β1 and 14-3-3β. Oncogene 2000, 20, 346–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, J.; Harada, H.; Yang, E.; Jockel, J.; Korsmeyer, S.J. Serine Phosphorylation of Death Agonist BAD in Response to Survival Factor Results in Binding to 14-3-3 Not BCL-XL. Cell 1996, 87, 619–628. [Google Scholar] [CrossRef] [Green Version]
- Masters, S.C.; Yang, H.; Datta, S.R.; Greenberg, M.E.; Fu, H. 14-3-3 Inhibits Bad-Induced Cell Death through Interaction with Serine-136. Mol. Pharmacol. 2001, 60, 1325–1331. [Google Scholar] [CrossRef] [Green Version]
- Porter, G.; Khuri, F.R.; Fu, H. Dynamic 14-3-3/client protein interactions integrate survival and apoptotic pathways. Semin. Cancer Biol. 2006, 16, 193–202. [Google Scholar] [CrossRef]
- Gardino, A.K.; Yaffe, M.B. 14-3-3 proteins as signaling integration points for cell cycle control and apoptosis. Semin. Cell Dev. Biol. 2011, 22, 688–695. [Google Scholar] [CrossRef]
- Knights, A.J.; Funnell, A.P.W.; Crossley, M.; Pearson, R.C. Holding Tight: Cell Junctions and Cancer Spread. Trends Cancer Res. 2012, 8, 61–69. [Google Scholar]
- Bhat, A.A.; Uppada, S.; Achkar, I.W.; Hashem, S.; Yadav, S.K.; Shanmugakonar, M.; Al-Naemi, H.A.; Haris, M.; Uddin, S. Tight Junction Proteins and Signaling Pathways in Cancer and Inflammation: A Functional Crosstalk. Front. Physiol. 2019, 9, 1942. [Google Scholar] [CrossRef] [Green Version]
- Beaufort, N.; Corvazier, E.; Mlanaoindrou, S.; De Bentzmann, S.; Pidard, D. Disruption of the Endothelial Barrier by Proteases from the Bacterial Pathogen Pseudomonas aeruginosa: Implication of Matrilysis and Receptor Cleavage. PLoS ONE 2013, 8, e75708. [Google Scholar] [CrossRef] [Green Version]
- Rejman, J.; Di Gioia, S.; Bragonzi, A.; Conese, M. Pseudomonas aeruginosaInfection Destroys the Barrier Function of Lung Epithelium and Enhances Polyplex-Mediated Transfection. Hum. Gene Ther. 2007, 18, 642–652. [Google Scholar] [CrossRef]
- Akunuru, S.; Palumbo, J.; Zhai, Q.J.; Zheng, Y. Rac1 Targeting Suppresses Human Non-Small Cell Lung Adenocarcinoma Cancer Stem Cell Activity. PLoS ONE 2011, 6, e16951. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Zhang, Y.; Miao, Y.; Zhao, Y.; Zhang, P.-X.; Jiang, G.; Zhang, J.-Y.; Han, Y.; Lin, X.-Y.; et al. Abnormal expression of p120-catenin, E-cadherin, and small GTPases is significantly associated with malignant phenotype of human lung cancer. Lung Cancer 2009, 63, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, J.; Pofahl, R.; Pfister, H.; Haase, I. Deletion of epidermal Rac1 inhibits HPV-8 induced skin papilloma formation and facilitates HPV-8- and UV-light induced skin carcinogenesis. Oncotarget 2016, 7, 57841–57850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajona, D.; Castaño, Z.; Garayoa, M.; Zudaire, E.; Pajares, M.J.; Martínez, A.; Cuttitta, F.; Montuenga, L.M.; Pío, R. Expression of Complement Factor H by Lung Cancer Cells. Cancer Res. 2004, 64, 6310–6318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.J.; Martínez, A.; Unsworth, E.J.; Thiele, C.J.; Moody, T.W.; Elsasser, T.; Cuttitta, F. Adrenomedullin expression in human tumor cell lines. Its potential role as an autocrine growth factor. J. Biol. Chem. 1996, 271, 23345–23351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiba-Ishii, A.; Kano, J.; Morishita, Y.; Sato, Y.; Minami, Y.; Noguchi, M. High expression of stratifin is a universal abnormality during the course of malignant progression of early-stage lung adenocarcinoma. Int. J. Cancer 2011, 129, 2445–2453. [Google Scholar] [CrossRef]
- Raungrut, P.; Wongkotsila, A.; Champoochana, N.; Lirdprapamongkol, K.; Svasti, J.; Thongsuksai, P. Knockdown of 14-3-3γ Suppresses Epithelial–Mesenchymal Transition and Reduces Metastatic Potential of Human Non-small Cell Lung Cancer Cells. Anticancer. Res. 2018, 38, 3507–3514. [Google Scholar] [CrossRef]
- Khorrami, A.; Bagheri, M.S.; Tavallaei, M.; Gharechahi, J. The functional significance of 14-3-3 proteins in cancer: Focus on lung cancer. Horm. Mol. Biol. Clin. Investig. 2017, 32. [Google Scholar] [CrossRef]
- Tong, S.; Xia, T.; Fan, K.; Jiang, K.; Zhai, W.; Li, J.-S.; Wang, S.-H.; Wang, J.-J. Loss of Par3 promotes lung adenocarcinoma metastasis through 14-3-3ζ protein. Oncotarget 2016, 7, 64260–64273. [Google Scholar] [CrossRef] [Green Version]
- Dalal, S.N.; Yaffe, M.B.; DeCaprio, J.A. 14-3-3 Family Members Act Coordinately to Regulate Mitotic Progression. Cell Cycle 2004, 3, 670–675. [Google Scholar] [CrossRef] [Green Version]
- Gowing, S.D.; Chow, S.C.; Cools-Lartigue, J.J.; Chen, C.B.; Najmeh, S.; Goodwin-Wilson, M.; Jiang, H.Y.; Bourdeau, F.; Beauchamp, A.; Angers, I.; et al. Gram-Negative Pneumonia Augments Non–Small Cell Lung Cancer Metastasis through Host Toll-like Receptor 4 Activation. J. Thorac. Oncol. 2019, 14, 2097–2108. [Google Scholar] [CrossRef] [PubMed]
- Chow, S.C.; Gowing, S.D.; Cools-Lartigue, J.J.; Chen, C.B.; Berube, J.; Yoon, H.W.; Chan, C.H.F.; Rousseau, M.C.; Bourdeau, F.; Giannias, B.; et al. Gram negative bacteria increase non-small cell lung cancer metastasis via toll-like receptor 4 activation and mitogen-activated protein kinase phosphorylation. Int. J. Cancer. 2015, 136, 1341–1350. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Bai, Y.; Zhao, S.; Liu, X.; Gao, Y.; Wang, L.; Liu, B.; Ma, D.; Ma, C. Gram-negative bacteria facilitate tumor progression through TLR4/IL-33 pathway in patients with non-small-cell lung cancer. Oncotarget 2018, 9, 13462–13473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, M.; Gu, X.; Han, Y.; Jin, M.; Ren, T. Gram-negative bacteria facilitate tumor outgrowth and metastasis by promoting lipid synthesis in lung cancer patients. J. Thorac. Dis. 2016, 8, 1943–1955. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Human Protein | Pathogen | Bacterial Protein | Type of Interaction | Method |
|---|---|---|---|---|
| CEACAM1 | MorCa | UspA1 | direct interaction | X-ray scattering, Molecular sieving, enzyme-linked Immunosorbent Assay(ELISA), Far Western Blotting (FWB), isothermal titration, calorimetry, cosedimentation |
| CFAH | Hif 10810 | Lipoprotein binding FH | direct interaction, physical association | FWB, ELISA |
| VTN | Hif 10810, NTHi strain 3655 | Surface-adhesin protein E, Lipoprotein binding FH | direct interaction, physical association | ELISA, bio-layer interferometry, fluorescence activated cell sorting |
| YWHAG | Psa | Exoenzyme S | association | pull down, experimental interaction |
| YWHAE, YWHAQ, YWHAH | Psa | Exoenzyme S | association, direct interaction | pull down, experimental interaction |
| YWHAB, YWHAZ | Psa | Exoenzyme S | association, direct interaction | pull down, experimental interaction, X-ray crystallography |
| SFN | Psa | Exoenzyme S | association, direct interaction | pull down, experimental interaction |
| RAC1 | Psa | Exoenzyme S | direct interaction | X-ray crystallography |
| ID | BP | P | FDR B&H | FDR B&Y | Bonferroni | Genes from Input | Genes in Annotation |
|---|---|---|---|---|---|---|---|
| GO:1900740 | positive regulation of protein insertion into mitochondrial membrane involved in apoptotic signaling pathway | 1.094 × 10−18 | 6.744 × 10−16 | 5.189 × 10−15 | 1.349 × 10−15 | YWHAB, YWHAE, YWHAG, YWHAH, YWHAQ, YWHAZ, SFN | 27 |
| GO:1900739 | regulation of protein insertion into mitochondrial membrane involved in apoptotic signaling pathway | 1.094 × 10−18 | 6.744 × 10−16 | 5.189 × 10−15 | 1.34910−15 | YWHAB, YWHAE, YWHAG, YWHAH, YWHAQ, YWHAZ, SFN | 27 |
| GO:0001844 | protein insertion into mitochondrial membrane involved in apoptotic signaling pathway | 3.237 × 10−18 | 1.330 × 10−15 | 1.024 × 10−14 | 3.991 × 10−15 | YWHAB, YWHAE, YWHAG, YWHAH, YWHAQ, YWHAZ, SFN | 31 |
| GO:1901030 | positive regulation of mitochondrial outer membrane permeabilization involved in apoptotic signaling pathway | 1.266 × 10−17 | 3.903 × 10−15 | 3.003 × 10−14 | 1.561 × 10−14 | YWHAB, YWHAE, YWHAG, YWHAH, YWHAQ, YWHAZ, SFN | 37 |
| GO:1901028 | regulation of mitochondrial outer membrane permeabilization involved in apoptotic signaling pathway | 9.037 × 10−17 | 1.857 × 10−14 | 1.429 × 10−13 | 1.114 × 10−13 | YWHAB, YWHAE, YWHAG, YWHAH, YWHAQ, YWHAZ, SFN | 48 |
| ID | CC | P | FDR B&H | FDR B&Y | Bonferroni | Genes from Input | Genes in Annotation |
|---|---|---|---|---|---|---|---|
| GO:0005912 | adherens junction | 3.191 × 10−9 | 2.683 × 10−7 | 1.482 × 10−6 | 4.467 × 10−7 | RAC1, CEACAM1, YWHAB, YWHAE, YWHAG, YWHAQ, YWHAZ | 560 |
| GO:0070161 | anchoring junction | 3.833 × 10−9 | 2.683 × 10−7 | 1.482 × 10−6 | 5.366 × 10−7 | RAC1, CEACAM1, YWHAB, YWHAE, YWHAG, YWHAQ, YWHAZ | 575 |
| GO:0005925 | focal adhesion | 2.594 × 10−8 | 1.048 × 10−6 | 5.785 × 10−6 | 3.632 × 10−6 | RAC1, YWHAB, YWHAE, YWHAG, YWHAQ, YWHAZ | 411 |
| GO:0030055 | cell-substrate junction | 2.993 × 10−8 | 1.048 × 10−6 | 5.785 × 10−6 | 4.190 × 10−6 | RAC1, YWHAB, YWHAE, YWHAG, YWHAQ, YWHAZ | 421 |
| GO:0030054 | cell junction | 4.686 × 10−8 | 1.312 × 10−6 | 7.245 × 10−6 | 6.560 × 10−6 | CEACAM1, RAC1, YWHAB, YWHAE, YWHAG, YWHAH, YWHAQ, YWHAZ | 1352 |
| ID | Disease | P | FDR B&H | FDR B&Y | Bonferroni | Genes from Input | Genes in Annotation |
|---|---|---|---|---|---|---|---|
| C0152013 | Adenocarcinoma of lung (disorder) | 1.879 × 10−6 | 7.698 × 10−4 | 5.393 × 10−3 | 1.163 × 10−3 | CFH, RAC1, SFN, YWHAB, YWHAE, YWHAG, YWHAZ | 1123 |
| C1720452 | Soft drusen | 2.487 × 10−6 | 7.698 × 10−4 | 5.393 × 10−3 | 1.540 × 10−3 | CFH, VTN | 4 |
| C0268731 | Renal glomerular disease | 5.155 × 10−5 | 7.575 × 10−3 | 5.307 × 10−2 | 3.191 × 10−2 | CFH, VTN, YWHAE | 113 |
| C0005586 | Bipolar Disorder | 6.287 × 10−5 | 7.575 × 10−3 | 5.307 × 10−2 | 3.892 × 10−2 | RAC1, VTN, YWHAE, YWHAH, YWHAZ | 723 |
| C0017662 | Glomerulonephritis, Membranoproliferative | 6.310 × 10−5 | 7.575 × 10−3 | 5.307 × 10−2 | 3.906 × 10−2 | CFH, VTN | 18 |
| Lung Adenocarcinoma | Apoptotic Processes | Focal Adhesion | Adherens Junctions | Anchoring Junctions | Cell-Substrate Junctions | Cell Junction |
|---|---|---|---|---|---|---|
| RAC1 | − | + | + | + | + | + |
| CFH | − | − | − | − | − | − |
| SFN | + | − | − | − | − | − |
| YWHAB | + | + | + | + | + | + |
| YWHAE | + | + | + | + | + | + |
| YWHAG | + | + | + | + | + | + |
| YWHAZ | + | + | + | + | + | + |
| Terms | Annotations | Term’s Genes Found | Term’s Genes | Genes Universe | Hyp pVal | Hyp pValAdj | Genes |
|---|---|---|---|---|---|---|---|
| positive regulation of protein insertion into mitochondrial membrane involved in apoptotic signaling pathway | GO:1900740 | 7 | 27 | 17898 | 2.50 × 10−18 | 5.01 × 10−16 | YWHAQ, YWHAZ, YWHAH, YWHAG, YWHAE, YWHAB, SFN |
| membrane organization | GO:0061024 | 7 | 135 | 17898 | 3.82 × 10−13 | 3.82 × 10−11 | YWHAQ, YWHAZ, YWHAH, YWHAG, YWHAE, YWHAB, SFN |
| protein targeting | GO:0006605 | 5 | 48 | 17898 | 5.11 × 10−11 | 3.41 × 10−9 | YWHAQ, YWHAZ, YWHAG, YWHAE, YWHAB |
| substantia nigra development | GO:0021762 | 3 | 46 | 17898 | 2.58 × 10−6 | 0.000129202 | YWHAQ, YWHAH, YWHAE |
| negative regulation of protein dephosphorylation | GO:0035308 | 2 | 10 | 17898 | 1.54 × 10−5 | 0.000513729 | YWHAE, YWHAB |
| Terms | Annotations | Term’s Genes Found | Term’s Genes | Genes Universe | Hyp pVal | Hyp pValAdj | Genes |
|---|---|---|---|---|---|---|---|
| extracellular exosome | GO:0070062 | 11 | 2169 | 18876 | 4.51 × 10−11 | 2.98 × 10−9 | YWHAQ, YWHAZ, YWHAH, YWHAG, YWHAE, YWHAB, VTN, CEACAM1, RAC1, CFH, SFN |
| focal adhesion | GO:0005925 | 6 | 410 | 18876 | 4.27 × 10−8 | 1.41 × 10−6 | YWHAQ, YWHAZ, YWHAG, YWHAE, YWHAB, RAC1 |
| melanosome | GO:0042470 | 4 | 100 | 18876 | 2.38 × 10−7 | 5.23 × 10−6 | YWHAZ, YWHAE, YWHAB, RAC1 |
| mitochondrion | GO:0005739 | 7 | 1538 | 18876 | 5.79 × 10−6 | 9.55 × 10−5 | YWHAQ, YWHAZ, YWHAH, YWHAG, YWHAE, YWHAB, SFN |
| blood microparticle | GO:0072562 | 3 | 143 | 18876 | 6.72 × 10−5 | 0.00088691 | YWHAZ, VTN, CFH |
| Terms | Annotations | Term’s Genes Found | Term’s Genes | Genes Universe | Hyp pVal | Hyp pValAdj | Genes |
|---|---|---|---|---|---|---|---|
| Cell cycle | hsa04110 | 7 | 124 | 8013 | 2.07 × 10−11 | 1.20 × 10−9 | YWHAQ, YWHAZ, YWHAH, YWHAG, YWHAE, YWHAB, SFN |
| Viral carcinogenesis | hsa05203 | 7 | 201 | 8013 | 6.34 × 10−10 | 1.23 × 10−8 | YWHAQ, YWHAZ, YWHAH, YWHAG, YWHAE, YWHAB, RAC1 |
| PI3K-Akt signaling pathway | hsa04151 | 8 | 354 | 8013 | 5.59 × 10−10 | 1.62 × 10−8 | YWHAQ, YWHAZ, YWHAH, YWHAG, YWHAE, YWHAB, VTN, RAC1 |
| Oocyte meiosis | hsa04114 | 6 | 128 | 8013 | 2.94 × 10−9 | 4.27 × 10−8 | YWHAQ, YWHAZ, YWHAH, YWHAG, YWHAE, YWHAB |
| Hepatitis C | hsa05160 | 6 | 155 | 8013 | 9.37 × 10−9 | 9.06 × 10−8 | YWHAQ, YWHAZ, YWHAH, YWHAG, YWHAE, YWHAB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giannakou, L.-E.; Giannopoulos, A.-S.; Hatzoglou, C.; Gourgoulianis, K.I.; Rouka, E.; Zarogiannis, S.G. Investigation and Functional Enrichment Analysis of the Human Host Interaction Network with Common Gram-Negative Respiratory Pathogens Predicts Possible Association with Lung Adenocarcinoma. Pathophysiology 2021, 28, 20-33. https://0-doi-org.brum.beds.ac.uk/10.3390/pathophysiology28010003
Giannakou L-E, Giannopoulos A-S, Hatzoglou C, Gourgoulianis KI, Rouka E, Zarogiannis SG. Investigation and Functional Enrichment Analysis of the Human Host Interaction Network with Common Gram-Negative Respiratory Pathogens Predicts Possible Association with Lung Adenocarcinoma. Pathophysiology. 2021; 28(1):20-33. https://0-doi-org.brum.beds.ac.uk/10.3390/pathophysiology28010003
Chicago/Turabian StyleGiannakou, Lydia-Eirini, Athanasios-Stefanos Giannopoulos, Chrissi Hatzoglou, Konstantinos I. Gourgoulianis, Erasmia Rouka, and Sotirios G. Zarogiannis. 2021. "Investigation and Functional Enrichment Analysis of the Human Host Interaction Network with Common Gram-Negative Respiratory Pathogens Predicts Possible Association with Lung Adenocarcinoma" Pathophysiology 28, no. 1: 20-33. https://0-doi-org.brum.beds.ac.uk/10.3390/pathophysiology28010003