Tryptophan and Kynurenine Enhances the Stemness and Osteogenic Differentiation of Bone Marrow-Derived Mesenchymal Stromal Cells In Vitro and In Vivo

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. BMSC Culture
2.2. Differentiation Assays
2.3. Flow-Cytometry (FCM)
2.4. Real-Time Reverse-Transcription Polymerase Chain Reaction (Real-Time RT-PCR)
2.5. Screening Process
2.5.1. First Screening
2.5.2. Second Screening
2.6. Immunocytochemistry
2.7. Cell Viability
2.8. Migration Assay
2.9. Animal Experiments
2.10. Colony-Forming Assay
2.11. Micro-Computed Tomographic Analysis
2.12. Histological Analysis
2.13. Statistical Analysis
3. Results
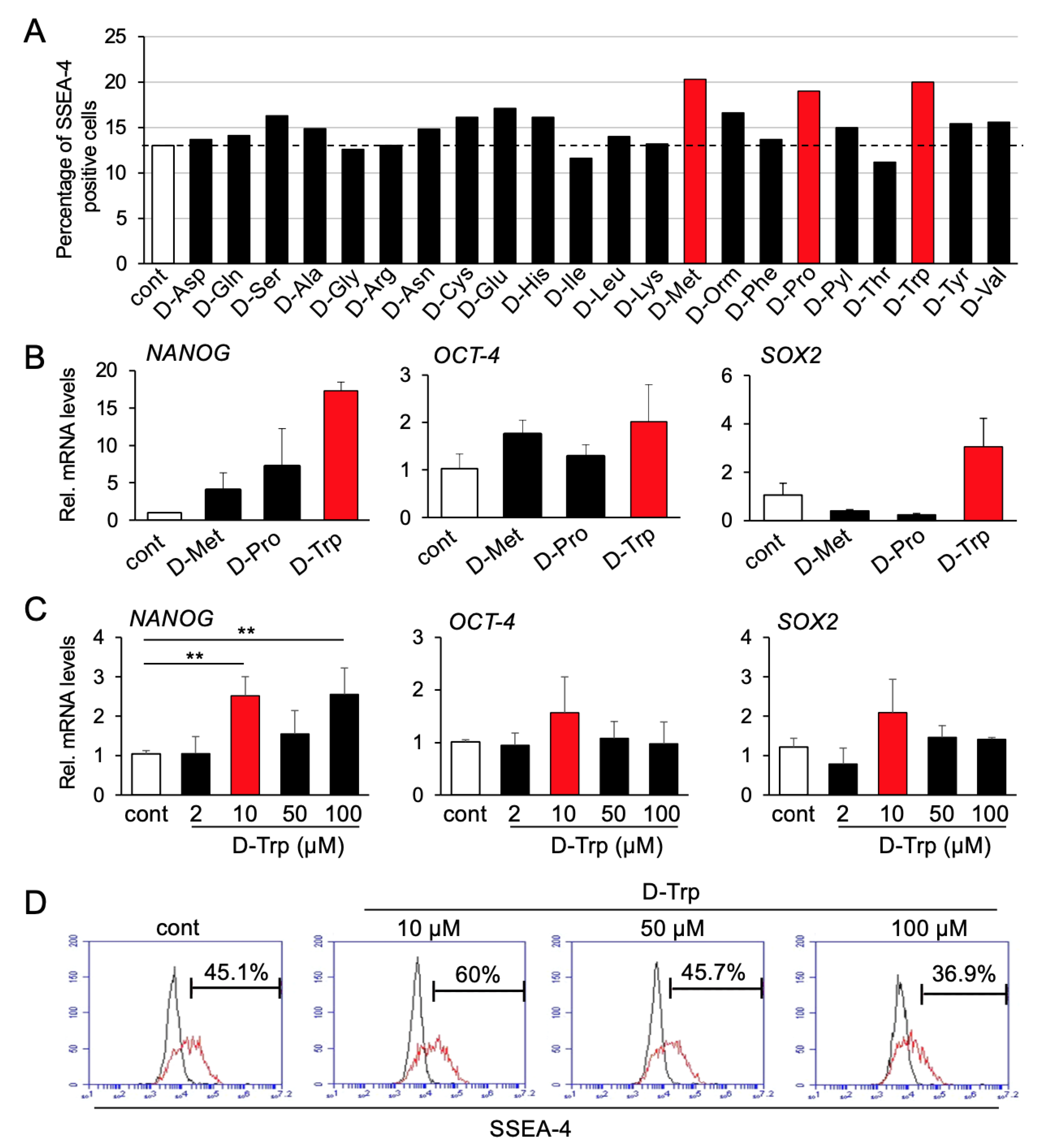
3.1. Screening Process
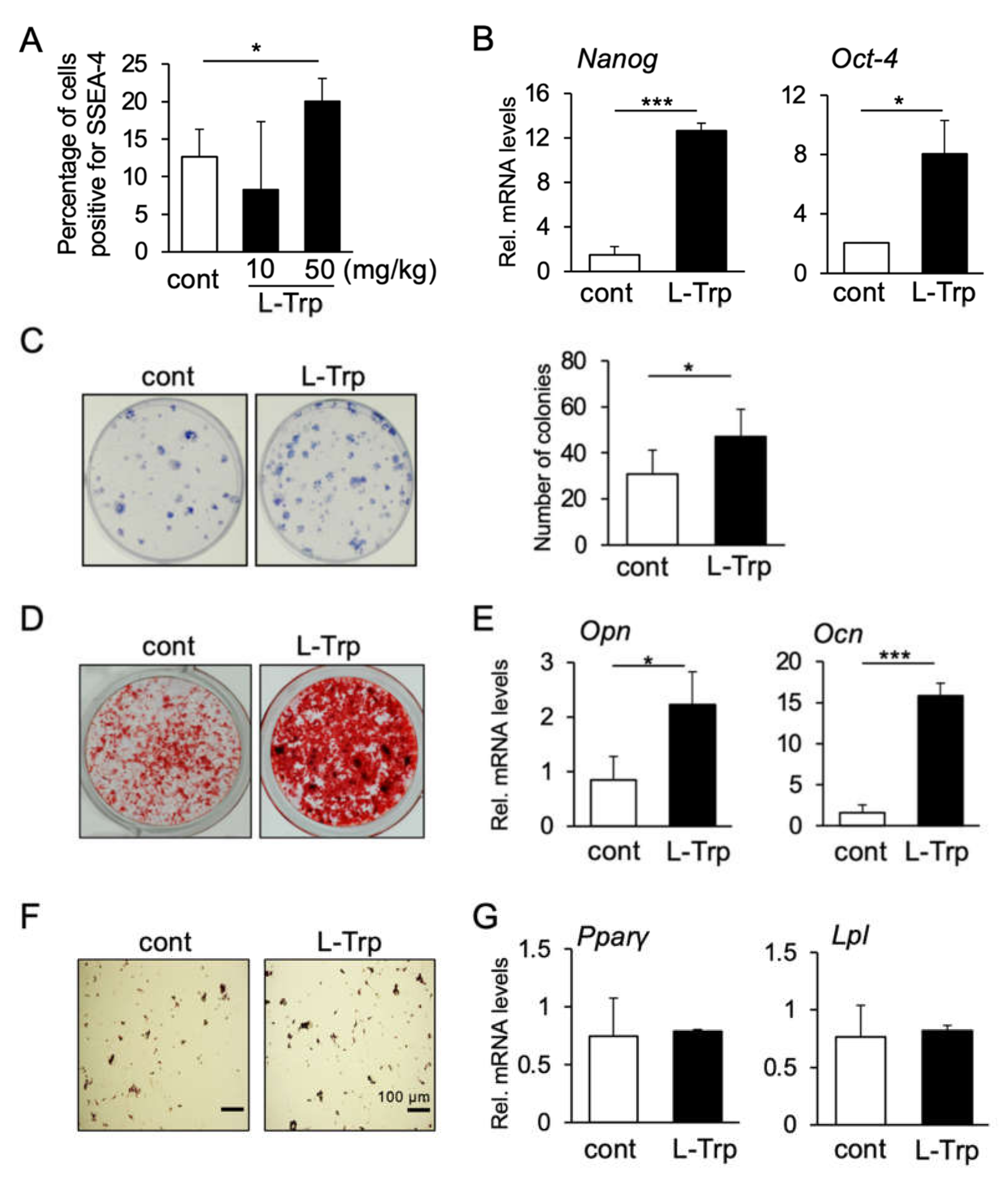
3.2. L-Tryptophan Enhances Migration, Colony Formation and Osteogenic Differentiation of hBMSCs In Vitro
3.3. L-Tryptophan Enhances Migration, Colony Formation and Osteogenic Differentiation of mBMSCs In Vivo
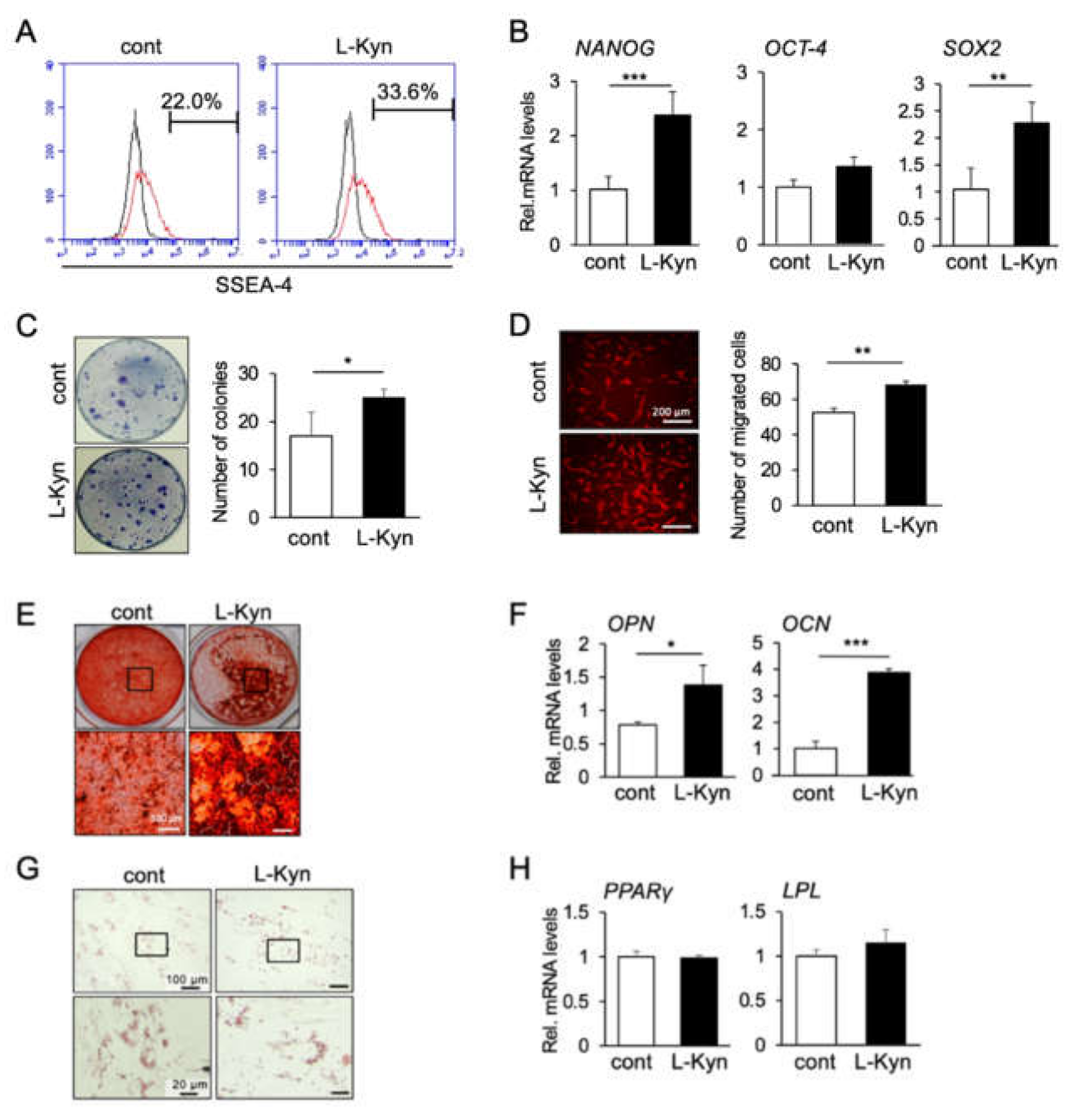
3.4. L-Kynurenine, A Main Metabolite of L-Tryptophan, Enhances the Stem Cell Phenotype of hBMSCs
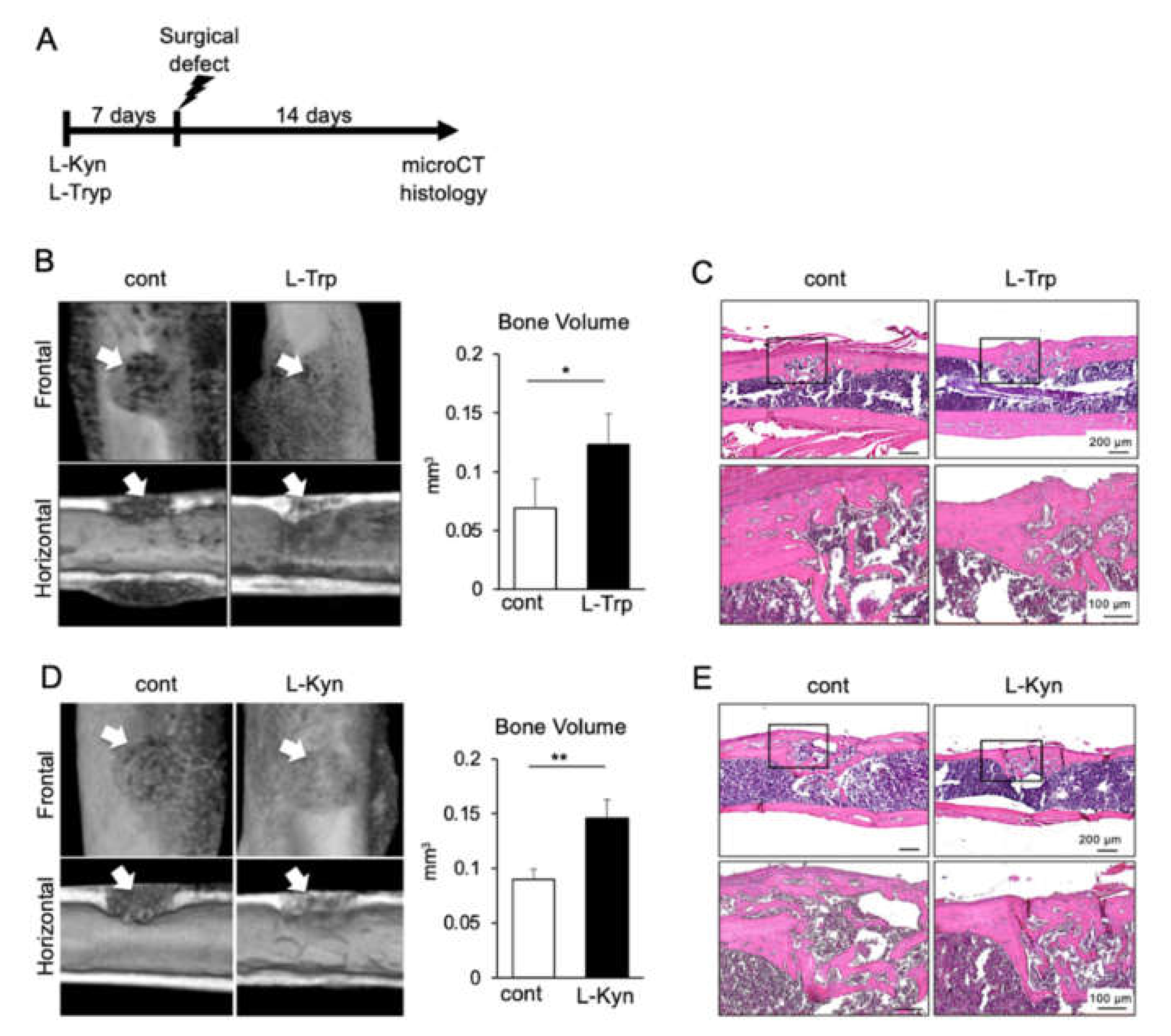
3.5. Both L-Tryptophan and L-Kynurenine Accelerate Bone Regeneration in Mouse Femur Surgical Defect
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oh, J.; Lee, D.Y.; Wagers, J.A. Stem cell aging: Mechanism, regulator and therapeuticopportunities. Nat. Med. 2014, 20, 870–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, M.C.; Han Calvi, M.L. Impact of aging on bone, marrow and their interactions. Bone 2019, 119, 1–7. [Google Scholar] [CrossRef]
- Thompson, J.F.; Morris, C.J.; Smith, I.K. New naturally occurring amino acids. Annu. Rev. Biochem. 1969, 38, 137–158. [Google Scholar] [CrossRef] [PubMed]
- Wagner, I.; Musso, H. New naturally-occurring amino-acids. Angew. Chem. Int. Ed. 1983, 22, 816–828. [Google Scholar] [CrossRef]
- Lide, D.R. Handbook of Chemistry and Physics, 72nd ed.; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Christensen, H.N. Role of amino acid transport and countertransport in nutrition and metabolism. Physiol. Rev. 1990, 70, 43–77. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L.; Stryer, L. Biochemistry, 5th ed.; W.H. Freeman: New York, NY, USA, 2002. [Google Scholar]
- Cheng, J.; Li, W.; Kang, B.; Zhou, Y.; Song, J.; Dan, S.; Yang, Y.; Zhang, X.; Li, J.; Yin, S.; et al. Tryptophan derivatives regulate the transcription of Oct4 in stem-like cancer cells. Nat. Commun. 2015, 6, 7209. [Google Scholar] [CrossRef] [Green Version]
- Watson, R.R.; Zibadi, S. Bioactive Dietary Factors and Plant Extracts in Dermatology; Humana Press: New York, NY, USA, 2013; p. 25. [Google Scholar]
- Taylor, L.; Curthoys, N.P. Glutamine metabolism: Role in acid-base balance. Biochem. Mol. Biol. Educ. 2004, 32, 291–304. [Google Scholar] [CrossRef]
- Shiraki, N.; Shiraki, Y.; Tsuyama, T.; Obata, F.; Miura, M.; Nagae, G.; Aburatani, H.; Kume, K.; Endo, F.; Kume, S. Methionine metabolism regulates maintenance and differentiation of human pluripotent stem cells. Cell Metab. 2014, 19, 780–794. [Google Scholar] [CrossRef] [Green Version]
- Ochocki, J.D.; Simon, M.C. Nutrient-sensing pathways and metabolic regulation in stem cells. J. Cell Biol. 2013, 203, 23–33. [Google Scholar] [CrossRef]
- Conigrave, A.D.; Brown, E.M.; Rizzoli, R. Dietary protein and bone health: Roles of amino acid-sensing receptors in the control of calcium metabolism and bone homeostasis. Annu. Rev. Nutr. 2008, 28, 131–155. [Google Scholar] [CrossRef]
- Ammann, P.; Laib, A.; Bonjour, J.P.; Meyer, J.M.; Ruegsegger, P.; Rizzoli, R. Dietary essential amino acid supplements increase bone strength by influencing bone mass and bone microarchitecture in ovariectomized adult rats fed an isocaloric low-protein diet. J. Bone Miner. Res. 2002, 17, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.J.; Hoekman, K. Arginine supplementation in the prevention and treatment of osteoporosis. Med. Hypothes. 1994, 43, 339–342. [Google Scholar] [CrossRef]
- Vidal, C.; Li, W.; Santner-Nanan, B.; Lim, C.K.; Guillemin, G.J.; Ball, H.J.; Hunt, N.H.; Nanan, R.; Duque, G. The Kynurenine pathway of Tryptophan degradation is activated during osteoblastogenesis. Stem Cells 2015, 33, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, K.; Mayer, B.; Sworder, B.J.; Kuznetsov, S.A.; Mezey, E. A practical guide to culturing mouse and human bone marrow stromal cells. Curr. Protoc. Immunol. 2013, 22F, 1–13. [Google Scholar] [CrossRef]
- Pham, H.T.; Kram, V.; Dar, Q.A.; Komori, T.; Youngmi, J.; Mohassel, P.; Rooney, J.; Li, L.; Kilts, T.M.; Bonnemann, C.; et al. Collagen VIα2 chain deficiency causes trabecular bone loss by potentially promoting osteclast differentiation through enhanced TNFα signaling. Sci. Rep. 2020, 10, 13794. [Google Scholar] [CrossRef]
- Ono, M.; Inkson, C.A.; Kilts, T.M.; Young, M.F. WISP-1/CCN4 regulates osteogenesis by enhancing BMP-2 activity. J. Bone Miner. Res. 2011, 26, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M.; Fujisawa, T.; Ono, M.; Hara, E.S.; Pham, H.T.; Nakajima, R.; Sonoyama, W.; Kuboki, T. A short-term treatment with tumor necrosis factor-alpha enhances stem cell phenotype of human dental pulp cells. Stem Cell Res. Ther. 2014, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Hara, E.S.; Ono, M.; Eguchi, T.; Kubota, S.; Pham, H.T.; Sonoyama, W.; Tajima, S.; Takigawa, M.; Calderwood, S.K.; Kuboki, T. miRNA-720 controls stem cell phenotype, proliferation and differentiation of human dental pulp cells. PLoS ONE 2013, 8, e83545. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, Y.; Ono, M.; Maeda, A.; Kilts, T.M.; Hara, E.S.; Khattab, H.; Ueda, J.; Aoyama, E.; Oohashi, T.; Takigawa, M.; et al. CCN4/WISP-1 positively regulates chondrogenesis by controlling TGF-beta3 function. Bone 2016, 83, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Gang, E.J.; Bosnakovski, D.; Figueiredo, C.A.; Visser, J.W.; Perlinggeiro, R.C.R. SSEA-4 identifies mesenchyma stem cells from bone marrow. Blood 2007, 109, 1743–1751. [Google Scholar] [CrossRef]
- Rodda, D.J.; Chew, J.L.; Lim, L.H.; Loh, Y.H.; Wang, B.; Ng, H.H.; Robson, P. Transcriptional regulation of nanog by OCT4 and SOX2. J. Biol. Chem. 2005, 280, 24731–24737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, E.S.; Ono, M.; Pham, H.T.; Sonoyama, W.; Kubota, S.; Takigawa, M.; Matsumoto, T.; Young, M.F.; Olsen, B.R.; Kuboki, T. Fluocinolone acetonide is a potent synergistic factor of TGF-beta 3-associated chondrogenesis of bone marrow-derived mesenchymal stem cells for articular surface regeneration. J. Bone Miner. Res. 2015, 30, 1585–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.C. Cell migration. Methods Mol. Biol. 2005, 294, 15–22. [Google Scholar] [PubMed]
- Matsushita, Y.; Sakamoto, K.; Tamamura, Y.; Shibata, Y.; Minamizato, T.; Kihara, T.; Ito, M.; Katsube, K.; Hiraoka, S.; Koseki, H.; et al. CCN3 protein participates in bone regeneration as an inhibitory factor. J. Biol. Chem. 2013, 288, 19973–19985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aung, K.T.; Akiyama, K.; Kunitomo, M.; Mun, A.Y.; Tosa, I.; Nguyen, H.T.T.; Zhang, J.; Kohno, T.; Ono, M.; Hara, E.S.; et al. Aging-affected MSC functions and severity of periodontal tissue destruction in a ligature-induced mouse periodontitis model. Int. J. Mol. Sci. 2020, 21, 8103. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Ono, M.; Oida, Y.; Hara, E.S.; Komori, T.; Akiyama, K.; Nguyen, H.T.T.; Aung, K.T.; Pham, H.T.; Tosa, I.; et al. Bone marrow cells inhibit BMP-2-induced osteoblast activity in the marrow environment. J. Bone Miner. Res. 2019, 34, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Morozov, L. Mirror symmetry breaking in biochemical evolution. Orig. Life 1979, 9, 187–217. [Google Scholar] [CrossRef]
- Botting, N.P. Chemistry and neurochemistry of the kynurenine pathway of tryptophan metabolism. Chem. Soc. Rev. 1995, 24, 401. [Google Scholar] [CrossRef]
- Nakajima, R.; Ono, M.; Hara, E.S.; Oida, Y.; Shinkawa, S.; Pham, H.T.; Akiyama, K.; Sonoyama, W.; Maekawa, K.; Kuboki, T. Mesenchymal stem/progenitor cell isolation from tooth extraction sockets. J. Dent. Res. 2014, 93, 1133–1140. [Google Scholar] [CrossRef] [Green Version]
- Sibilia, V.; Pagani, F.; Lattuada, N.; Greco, A.; Guidobono, F. Linking chronic tryptophan deficiency with impaired bone metabolism and reduced bone accrual in growing rats. J. Cell. Biochem. 2009, 107, 890–898. [Google Scholar] [CrossRef]
- Galli, C.; Macaluso, G.; Passeri, G. Serotonin: A novel bone mass controller may have implications for alveolar bone. J. Negat. Results Biomed. 2013, 12, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inose, H.; Zhou, B.; Yadav, V.K.; Guo, X.E.; Karsenty, G.; Ducy, P. Efficacy of serotonin inhibition in mouse models of bone loss. J. Bone Miner. Res. 2011, 26, 2002–2011. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.K.; Balaji, S.; Suresh, P.S.; Liu, X.S.; Lu, X.; Li, Z.; Guo, X.E.; Mann, J.J.; Balapure, A.K.; Gershon, M.D.; et al. Pharmacological inhibition of gut-derived serotonin synthesis is a potential bone anabolic treatment for osteoporosis. Nat. Med. 2010, 16, 308–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, T.W.; Darlington, L.G. Endogenous kynurenines as targets for drug discovery and development. Nat. Rev. Drug Discov. 2002, 1, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Apalset, E.M.; Gjesdal, C.G.; Ueland, P.M.; Oyen, J.; Meyer, K.; Midttun, O.; Eide, G.E.; Tell, G.S. Interferon gamma (IFN-gamma)-mediated inflammation and the kynurenine pathway in relation to risk of hip fractures: The Hordaland Health Study. Osteoporos. Int. 2014, 25, 2067–2075. [Google Scholar] [CrossRef]
- Apalset, E.M.; Gjesdal, C.G.; Ueland, P.M.; Midttun, O.; Ulvik, A.; Eide, G.E.; Meyer, K.; Tell, G.S. Interferon (IFN)-gamma-mediated inflammation and the kynurenine pathway in relation to bone mineral density: The Hordaland Health Study. Clin. Exp. Immunol. 2014, 176, 452–460. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | D-Trp (10 μM) | L-Trp (10 μM) | |
|---|---|---|---|
| Stem Cell Marker | |||
| SSEA-4+ cells | 1.39 ± 0.03 (***) | 1.21 ± 0.08 (*) | |
| NANOG mRNA levels | 2.25 ± 0.59 (***) | 3.57 ±0.96 (***) | |
| OCT-4 mRNA levels | 1.23 ± 0.43 (NS) | 1.78 ± 0.86 (NS) | |
| SOX2 mRNA levels | 1.45 ± 0.34 (*) | 2.74 ± 1.07 (**) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, H.T.; Ono, M.; Hara, E.S.; Nguyen, H.T.T.; Dang, A.T.; Do, H.T.; Komori, T.; Tosa, I.; Hazehara-Kunitomo, Y.; Yoshioka, Y.; et al. Tryptophan and Kynurenine Enhances the Stemness and Osteogenic Differentiation of Bone Marrow-Derived Mesenchymal Stromal Cells In Vitro and In Vivo. Materials 2021, 14, 208. https://0-doi-org.brum.beds.ac.uk/10.3390/ma14010208
Pham HT, Ono M, Hara ES, Nguyen HTT, Dang AT, Do HT, Komori T, Tosa I, Hazehara-Kunitomo Y, Yoshioka Y, et al. Tryptophan and Kynurenine Enhances the Stemness and Osteogenic Differentiation of Bone Marrow-Derived Mesenchymal Stromal Cells In Vitro and In Vivo. Materials. 2021; 14(1):208. https://0-doi-org.brum.beds.ac.uk/10.3390/ma14010208
Chicago/Turabian StylePham, Hai Thanh, Mitsuaki Ono, Emilio Satoshi Hara, Ha Thi Thu Nguyen, Anh Tuan Dang, Hang Thuy Do, Taishi Komori, Ikue Tosa, Yuri Hazehara-Kunitomo, Yuya Yoshioka, and et al. 2021. "Tryptophan and Kynurenine Enhances the Stemness and Osteogenic Differentiation of Bone Marrow-Derived Mesenchymal Stromal Cells In Vitro and In Vivo" Materials 14, no. 1: 208. https://0-doi-org.brum.beds.ac.uk/10.3390/ma14010208