
Xeno-Free Materials for Stabilizing Basic Fibroblast Growth Factor and Enhancing Cell Proliferation in Human Pluripotent Stem Cell Cultures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Synthesis of Sulfated Materials
2.3. Measurement of bFGF Concentration in Culture Media Using ELISA
2.4. Evaluation of Test Compounds Using ELISA
2.5. Precoating of Culture Substrates
2.6. Cells and Culture Conditions
2.7. Statistical Analysis
3. Results
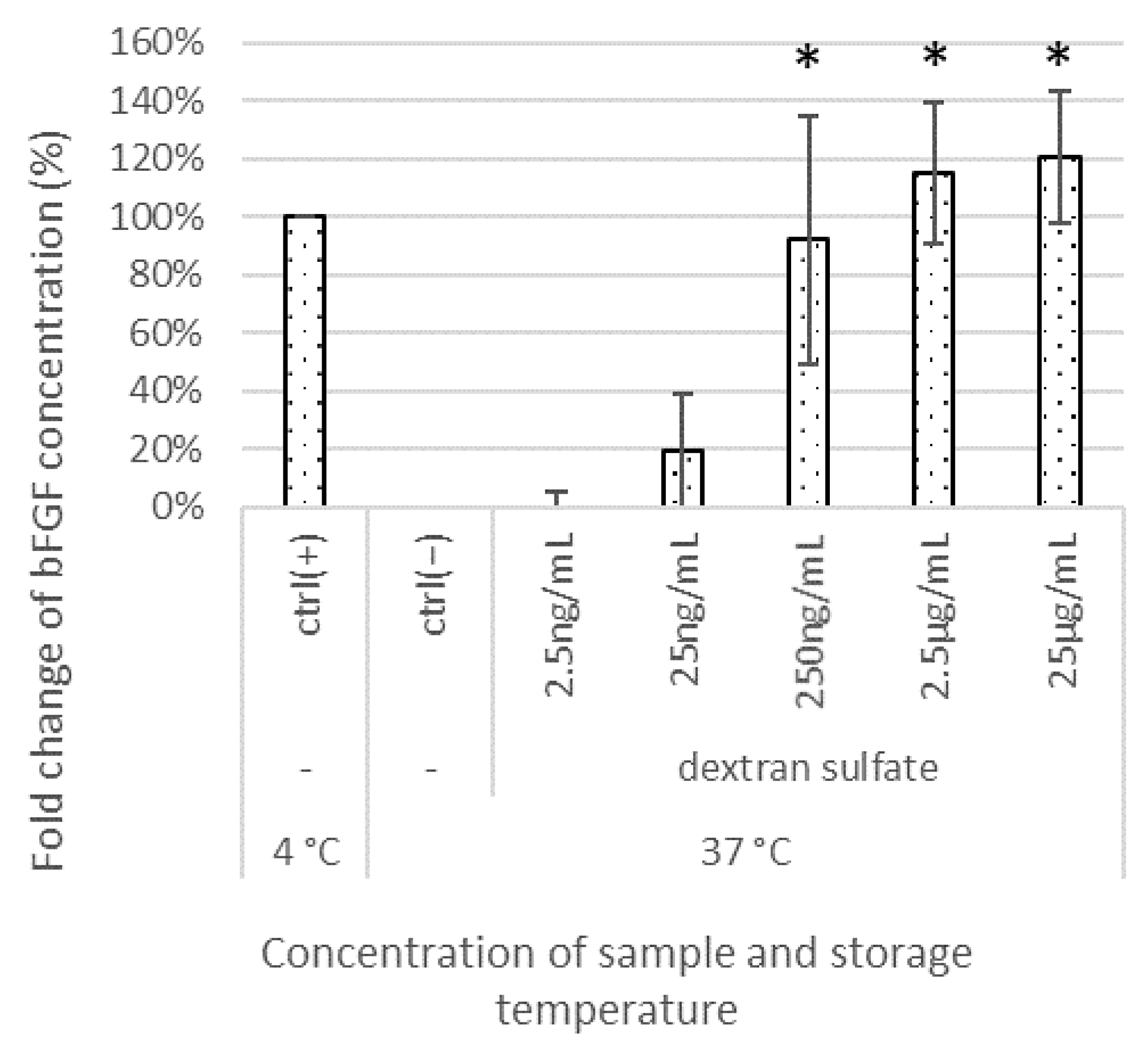
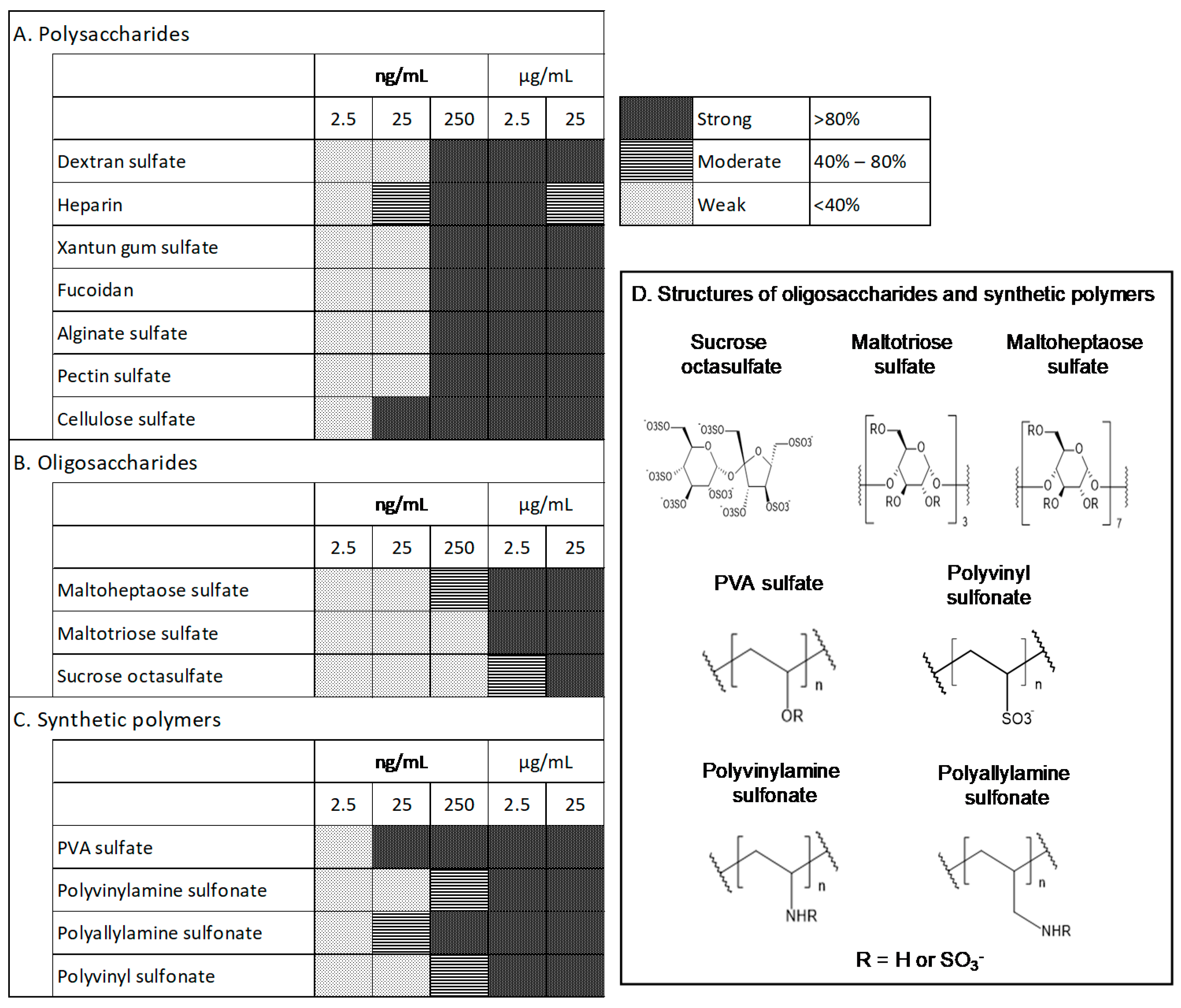
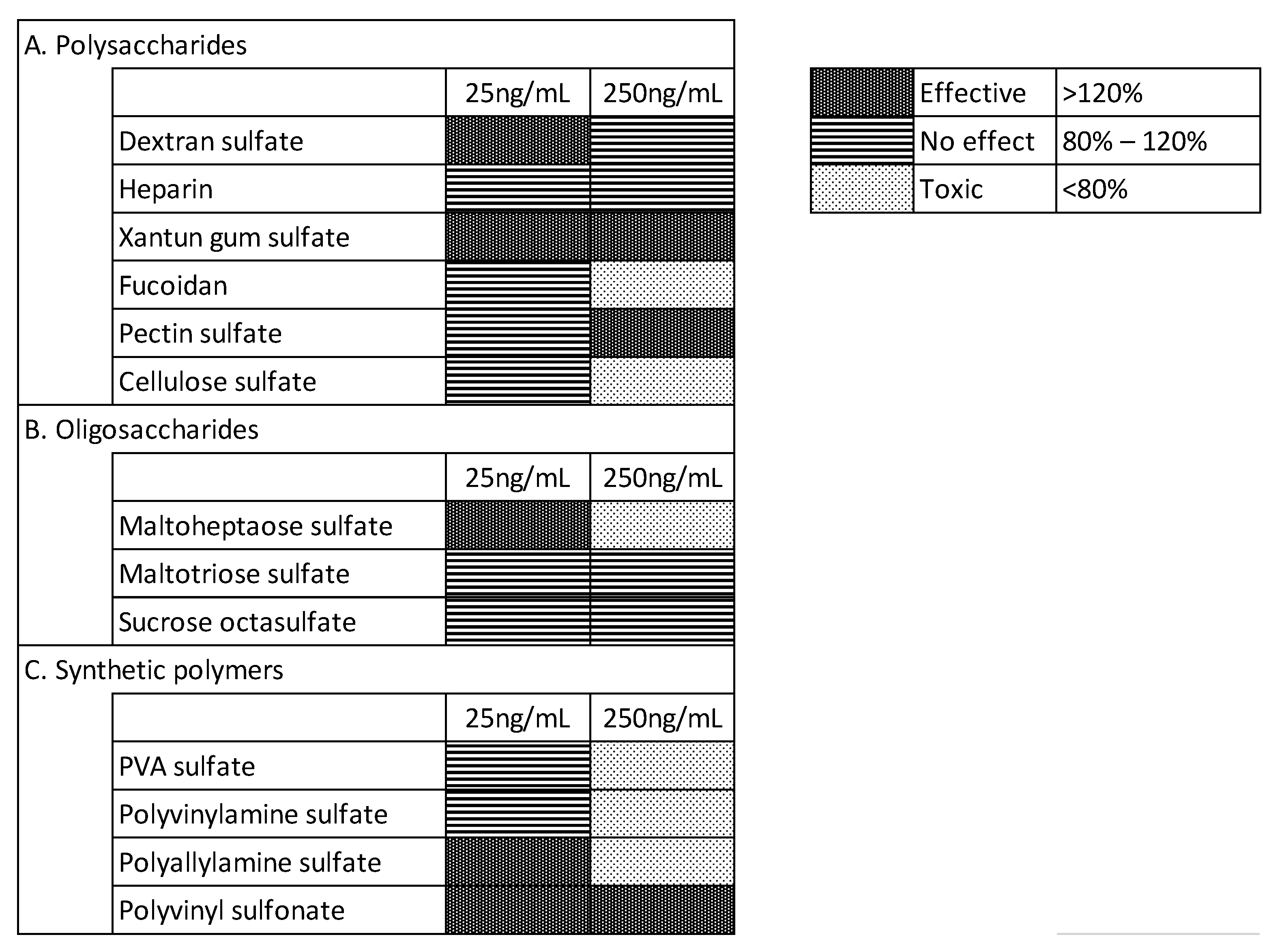
3.1. Effects of Test Materials on bFGF Stabilization
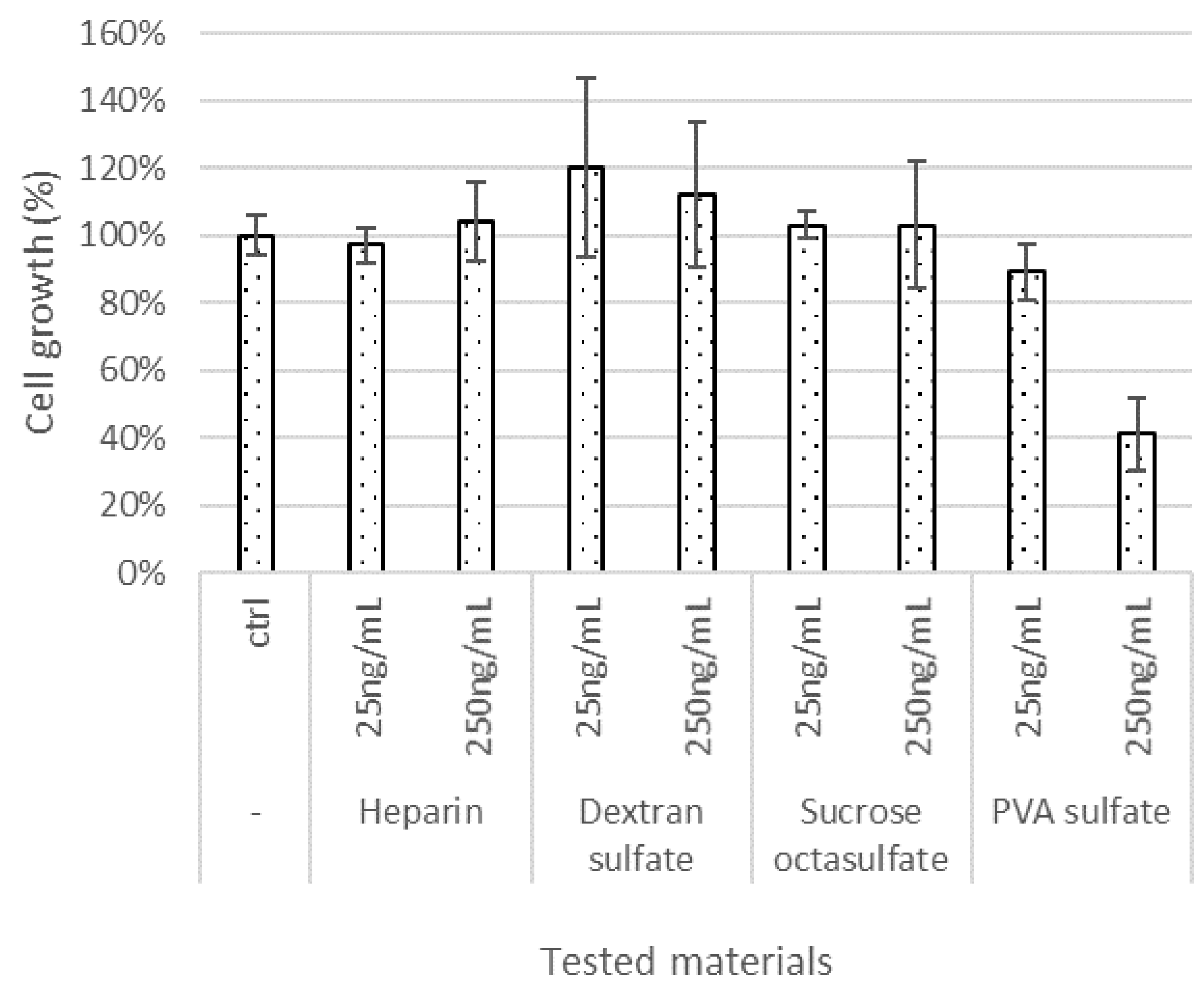
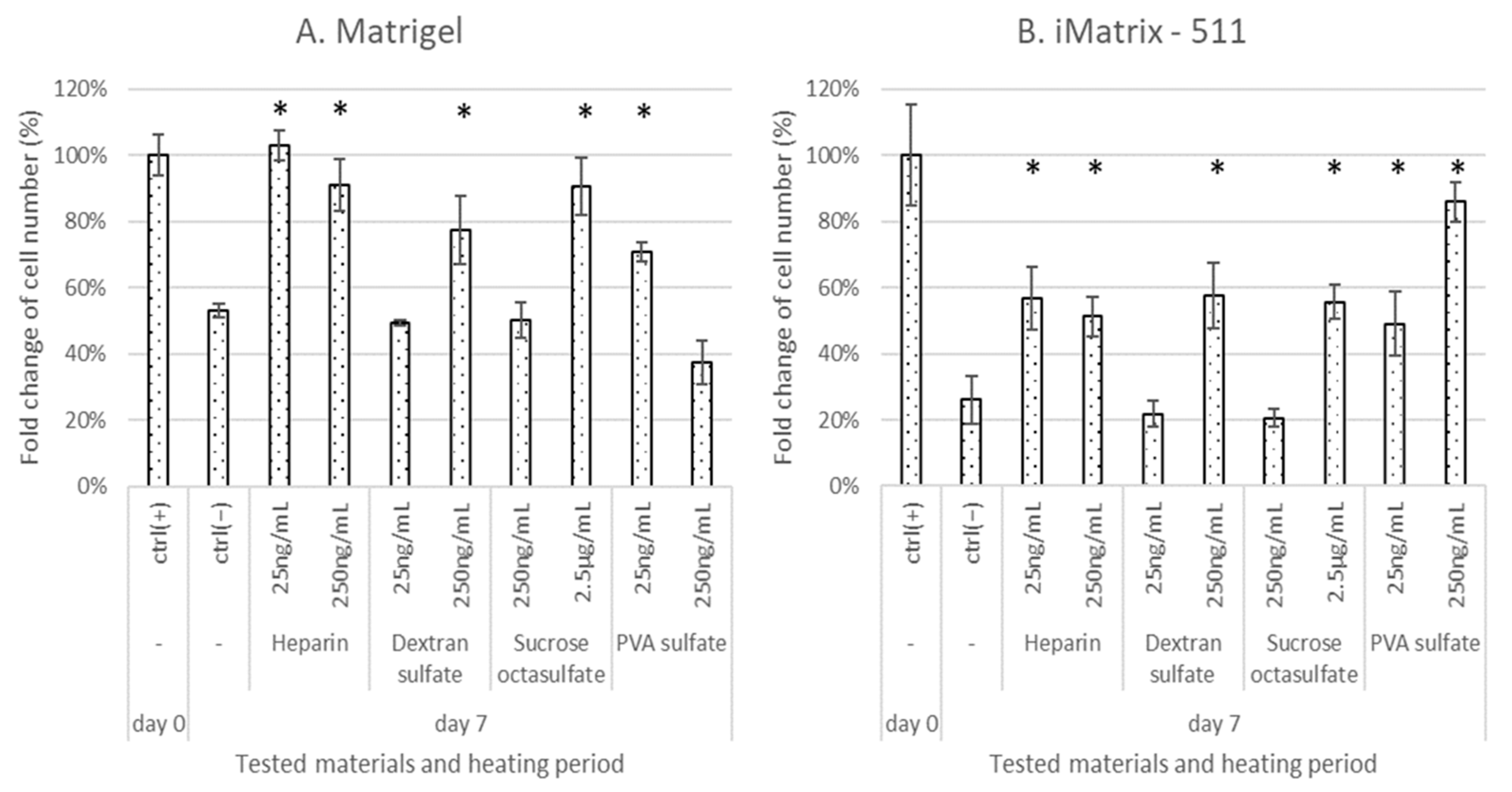
3.2. Effect of Test Materials on Cell Proliferation
3.3. Effects of the Test Materials on Medium Performance after Storage at 37 °C for 1 Week
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okita, K.; Ichisaka, T.; Yamanaka, S. Generation of germline-competent induced pluripotent stem cells. Nature 2007, 448, 313–317. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, J.; Woltjen, K.; Kaji, K. New strategies to generate induced pluripotent stem cells. Curr. Opin. Biotechnol. 2009, 20, 516–521. [Google Scholar] [CrossRef]
- González, F.; Boué, S.; Belmonte, J. Methods for making induced pluripotent stem cells: Reprogramming à la carte. Nat. Rev. Genet. 2011, 12, 231–242. [Google Scholar] [CrossRef]
- Haridhasapavalan, K.; Borgohain, M.; Dey, C.; Saha, B.; Narayan, G.; Kumar, S.; Thummer, R. An insight into non-integrative gene delivery approaches to generate transgene-free induced pluripotent stem cells. Gene 2019, 686, 146–159. [Google Scholar] [CrossRef]
- Mochiduki, Y.; Okita, K. Methods for iPS cell generation for basic research and clinical applications. Biotechnol. J. 2012, 7, 789–797. [Google Scholar] [CrossRef]
- Lavon, N.; Zimerman, M.; Itskovitz-Eldor, J. Scalable expansion of pluripotent stem cells. In Engineering and Application of Pluripotent Stem Cells: Advances in Biochemical Engineering/Biotechnology; Martin, U., Zweigerdt, R., Gruh, I., Eds.; Springer: Cham, Switzerland, 2018; Volume 163, pp. 23–27. [Google Scholar] [CrossRef]
- Nakagawa, M.; Taniguchi, Y.; Senda, S.; Takizawa, N.; Ichisaka, T.; Asano, K.; Morizane, A.; Doi, D.; Takahashi, J.; Nishizawa, M.; et al. A novel efficient feeder-free culture system for the derivation of human induced pluripotent stem cells. Sci. Rep. 2014, 4, 3594. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Rosler, E.; Jiang, J.; Lebkowksi, J.S.; Gold, J.D.; O’Sullivan, C.; Delavan-Boorsma, K.; Mok, M.; Bronstein, A.; Carpenter, M.K.; et al. Basic fibroblast growth factor supports undifferentiated human embryonic stem cell growth without conditioned medium. Stem Cells 2009, 23, 315–323. [Google Scholar] [CrossRef]
- Dvorak, P.; Dvorakova, D.; Koskova, S.; Vodinska, M.; Najvirtova, M.; Krekac, D.; Hampl, A. Expression and potential role of fibroblast growth factor 2 and its receptors in human embryonic stem cells. Stem Cells 2005, 23, 1200–1211. [Google Scholar] [CrossRef]
- Avery, S.; Inniss, K.; Moore, H. The regulation of self-renewal in human embryonic stem cells. Stem Cells Dev. 2006, 15, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Ding, V.M.Y.; Boersema, P.J.; Foong, L.Y.; Preisinger, C.; Koh, G.; Natarajan, S.; Lee, D.-Y.; Noekhorst, J.; Snei, B.; Lemeer, S.; et al. Tyrosine phosphorylation profiling in FGF-2 stimulated human embryonic stem cells. PLoS ONE 2011, 6, e17538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlodavsky, I.; Korner, G.; Ishai-Michaeli, R.; Bashkin, P.; Bar-Shavit, R.; Fuks, Z. Extracellular matrix-resident growth factors and enzymes: Possible involvement in tumor metastasis and angiogenesis. Cancer Metastasis Rev. 1990, 9, 203–226. [Google Scholar] [CrossRef] [PubMed]
- Green, P.J.; Walsh, F.S.; Doherty, P. Promiscuity of fibroblast growth factor receptors. Bioessays 1996, 18, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Shahrokh, Z.; Wang, Y.J.; Stratton, P.R.; Eberlein, G. Approaches to analysis of aggregates and demonstrating mass balance in pharmaceutical protein (basic fibroblast growth factor) formulations. J. Pharm. Sci. 1994, 83, 1645–1650. [Google Scholar] [CrossRef]
- Xiang, L.; Ishii, T.; Hosoda, K.; Kamiya, A.; Enomoto, M.; Nameki, N.; Inoue, Y.; Kubota, K.; Kohno, T.; Wakamatsu, K. Interaction of anti-aggregation agent dimethylethylammonium propane sulfonate with acidic fibroblast growth factor. J. Magn. Reson. 2008, 194, 147–151. [Google Scholar] [CrossRef]
- Sun, C.; Liu, M.; Sun, P.; Yang, M.; Yates, E.A.; Gui, Z.; Fernig, D.G. Sulfated polysaccharides interact with fibroblast growth factors and protect from denaturation. FEBS Open Bio 2019, 9, 1477–1487. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.-L.; Che, P.-P.; ZhuGe, Q.-Y.; Jin, B.-H.; Shen, B.-X.; Xiao, J.; Zhao, Y.-Z. Liposomes with silk fibroin hydrogel core to stabilize bFGF and promote the wound healing of mice with deep second-degree scald. Adv. Healthc. Mater. 2017, 6, 1700344. [Google Scholar] [CrossRef]
- Yu, X.; Biedrzycki, A.H.; Khalil, A.S.; Hess, D.; Umhoefer, J.M.; Markel, M.D.; Murphy, W.L. Nanostructured mineral coatings stabilize proteins for therapeutic delivery. Adv. Mater. 2017, 29, 1701255. [Google Scholar] [CrossRef]
- Masuzawa, Y.; Kitazawa, M. Synthetic polymers as xeno-free materials for stabilizing basic fibroblast growth factor in human mesenchymal stem cell cultures. Biochem. Biophys. Rep. 2021, 25, 100929. [Google Scholar] [CrossRef]
- Onuma, Y.; Higuchi, K.; Aiki, Y.; Shu, Y.; Asada, M.; Asahima, M.; Suzuki, M.; Imamura, T.; Ito, Y. A stable chimeric fibroblast growth factor (FGF) can successfully replace basic FGF in human pluripotent stem cell culture. PLoS ONE 2015, 10, e0118931. [Google Scholar] [CrossRef]
- Han, U.; Park, H.H.; Kim, Y.J.; Park, T.H.; Park, J.H.; Hong, J. Efficient encapsulation and sustained release of basic fibroblast growth factor in nanofilm: Extension of the feeding cycle of human induced pluripotent stem cell culture. ACS Appl. Mater. Interf. 2017, 9, 25087–25097. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, S.; Kajiwara, T.; Mizumoto, H. Expansion and differentiation of human iPS cells in a three-dimensional culture using hollow fibers and separation of the specific population by magnetic-activated cell sorting. J. Biosci. Bioeng. 2019, 128, 480–486. [Google Scholar] [CrossRef] [PubMed]
- McGrath, M.; Tam, E.; Sladkova, M.; AlManaie, A.; Zimmer, M.; de Peppo, G.M. GMP-compatible and xeno-free cultivation of mesenchymal progenitors derived from human-induced pluripotent stem cells. Stem Cell Res. Ther. 2019, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Parr, A.M.; Walsh, P.J.; Truong, V.; Dutton, J.R. cGMP-compliant expansion of human iPSC cultures as adherent monolayers. In Induced Pluripotent Stem (iPS) Cells: Methods in Molecular Biology; Humana Press: New York, NY, USA, 2015; Volume 1357, pp. 221–229. [Google Scholar] [CrossRef]
- Koike, H.; Kubota, K.; Sekine, K.; Takebe, T.; Rie, O.; Zheng, Y.-W.; Ueno, Y.; Tanigawa, N.; Taniguchi, H. Establishment of automated culture system for murine induced pluripotent stem cells. BMC Biotechnol. 2012, 12, 81. [Google Scholar] [CrossRef] [Green Version]
- Rohani, L.; Johnson, A.A.; Naghsh, P.; Rancourt, D.E.; Ulrich, H.; Holland, H. Concise review: Molecular cytogenetics and quality control: Clinical guardians for pluripotent stem cells. Stem Cells Transl. Med. 2018, 7, 867–875. [Google Scholar] [CrossRef]
- Kuo, H.-H.; Gao, X.; DeKeyser, J.M.; Fetterman, K.A.; Pinheiro, E.A.; Weddle, C.J.; Fonoudi, H.; Orman, M.V.; Romero-Tejeda, M.; Jouni, M.; et al. Negligible-cost and weekend-free chemically defined human iPSC culture. Stem Cell Rep. 2020, 14, 256–270. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.K.; Na, J.; Jackson, J.P.; Okamoto, T.; Jones, M.; Baker, D.; Hata, R.-I.; Moore, H.D.; Sato, J.D.; Andrews, P.W. Heparin promotes the growth of human embryonic stem cells in a defined serum-free medium. Proc. Natl. Acad. Sci. USA 2008, 105, 13409–13414. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Kim, M.H.; Agung, E.; Senda, S.; Kino-oka, M. Effect of migratory behaviors on human induced pluripotent stem cell colony formation on different extracellular matrix proteins. Regen. Ther. 2019, 10, 27–35. [Google Scholar] [CrossRef]
- Lennon, D.P.; Haynesworth, S.E.; Bruder, S.P.; Jaiswal, N.; Caplan, A.I. Human and animal mesenchymal progenitor cells from bone marrow: Identification of serum for optimal selection and proliferation. In Vitro Cell. Dev. Biol. Anim. 1996, 32, 602–611. [Google Scholar] [CrossRef]
- Chen, G.; Gulbranson, D.R.; Hou, Z.; Bolin, J.M.; Ruotti, V.; Probasco, M.D.; Smuga-Otto, K.; Howden, S.E.; Diol, N.R.; Propson, N.E.; et al. Chemically defined conditions for human iPSC derivation and culture. Nat. Methods 2011, 8, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geoffrey, L.F. Albumin and mammalian cell culture: Implications for biotechnology applications. Cytotechnology 2010, 62, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Uygun, B.E.; Stojsih, S.E.; Matthew, H.W.T. Effects of immobilized glycosaminoglycans on the proliferation and differentiation of mesenchymal stem cells. Tissue Eng. Part A 2009, 15, 3499–3512. [Google Scholar] [CrossRef] [PubMed]
- Singh, U.; Quintanilla, R.; Grecian, S.; Gee, K.; Rao, M.; Lakshmipathy, U. Novel Live Alkaline Phosphatase Substrate for Identification of Pluripotent Stem Cells. Stem. Cell Rev. 2012, 8, 1021–1029. [Google Scholar] [CrossRef] [Green Version]
- Nagasaka, R.; Matsumoto, M.; Okada, M.; Sasaki, H.; Kanie, K.; Kii, K.; Uozumi, T.; Kiyota, Y.; Honda, H.; Kato, R. Visualization of morphological categories of colonies for monitoring of effect on induced pluripotent stem cell culture status. Regen. Ther. 2017, 6, 41–51. [Google Scholar] [CrossRef]
- Miyazaki, T.; Futaki, S.; Suemori, H.; Taniguchi, Y.; Yamada, M.; Kawasaki, M.; Hayashi, M.; Kumegai, H.; Nakatsuji, K.; Sekiguchi, K.; et al. Laminin E8 fragments support efficient adhesion and expansion of dissociated human pluripotent stem cells. Nat. Commun. 2012, 3, 1236. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.S.; Postovit, L.M.; Lajoie, G.A. Matrigel: A complex protein mixture required for optimal growth of cell culture. Proteomics 2010, 10, 1886–1890. [Google Scholar] [CrossRef]
- Doğan, A.; Yalvaç, M.E.; Şahin, F.; Kabanov, A.V.; Palotás, A.; Rizvanov, A.A. Differentiation of human stem cells is promoted by amphiphilic pluronic block copolymers. Int. J. Nanomed. 2012, 7, 4849–4860. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masuzawa, Y.; Kitazawa, M. Xeno-Free Materials for Stabilizing Basic Fibroblast Growth Factor and Enhancing Cell Proliferation in Human Pluripotent Stem Cell Cultures. Materials 2022, 15, 3687. https://0-doi-org.brum.beds.ac.uk/10.3390/ma15103687
Masuzawa Y, Kitazawa M. Xeno-Free Materials for Stabilizing Basic Fibroblast Growth Factor and Enhancing Cell Proliferation in Human Pluripotent Stem Cell Cultures. Materials. 2022; 15(10):3687. https://0-doi-org.brum.beds.ac.uk/10.3390/ma15103687
Chicago/Turabian StyleMasuzawa, Yoko, and Manabu Kitazawa. 2022. "Xeno-Free Materials for Stabilizing Basic Fibroblast Growth Factor and Enhancing Cell Proliferation in Human Pluripotent Stem Cell Cultures" Materials 15, no. 10: 3687. https://0-doi-org.brum.beds.ac.uk/10.3390/ma15103687