Timber and Branch Volume Prediction: Effects of Stand and Site Characteristics on Dendromass and Timber-To-Branch Volume Ratio of Norway Spruce in Managed Forests



Abstract
:1. Introduction
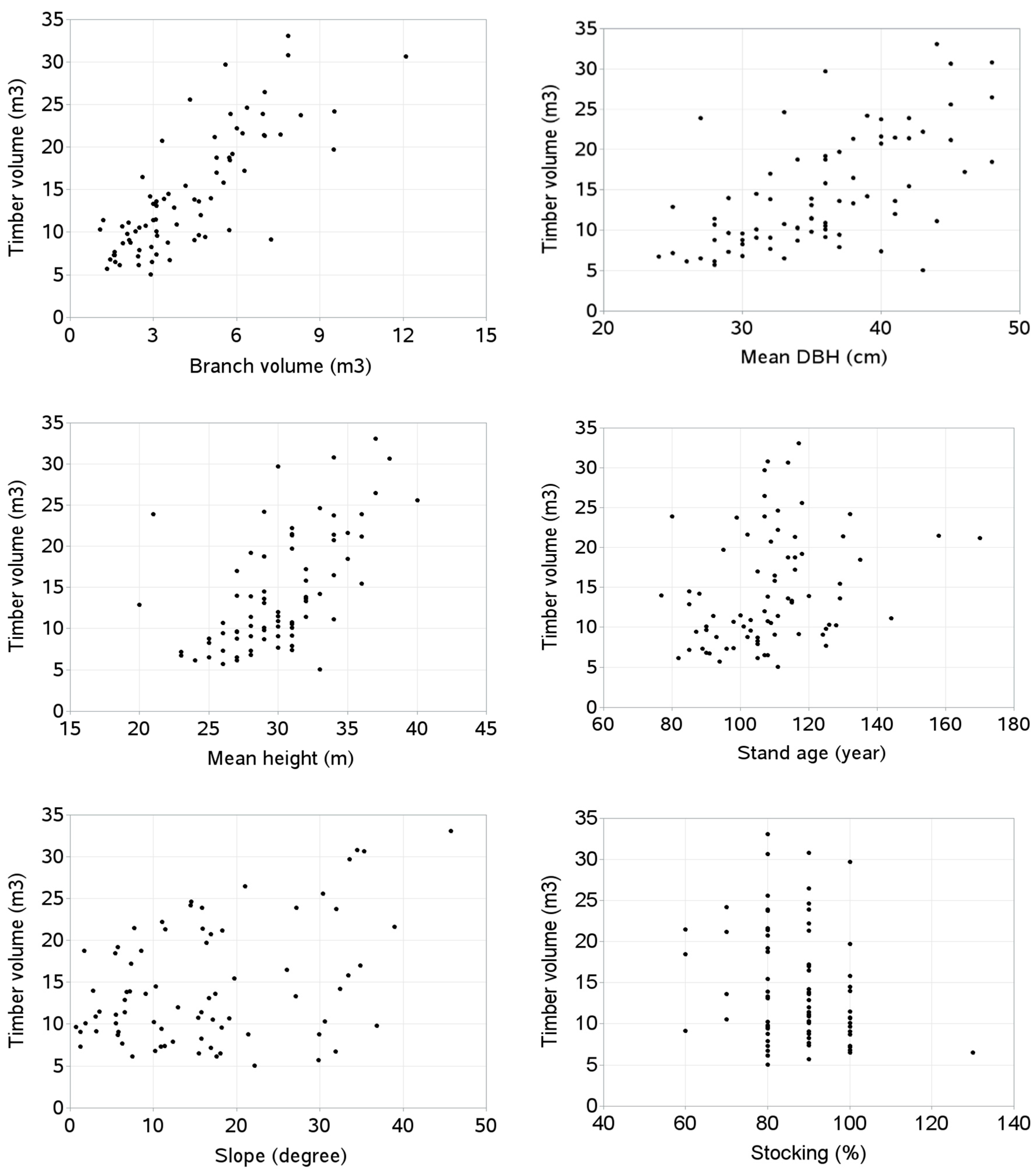
- What stand and site characteristics are correlated with timber volume?
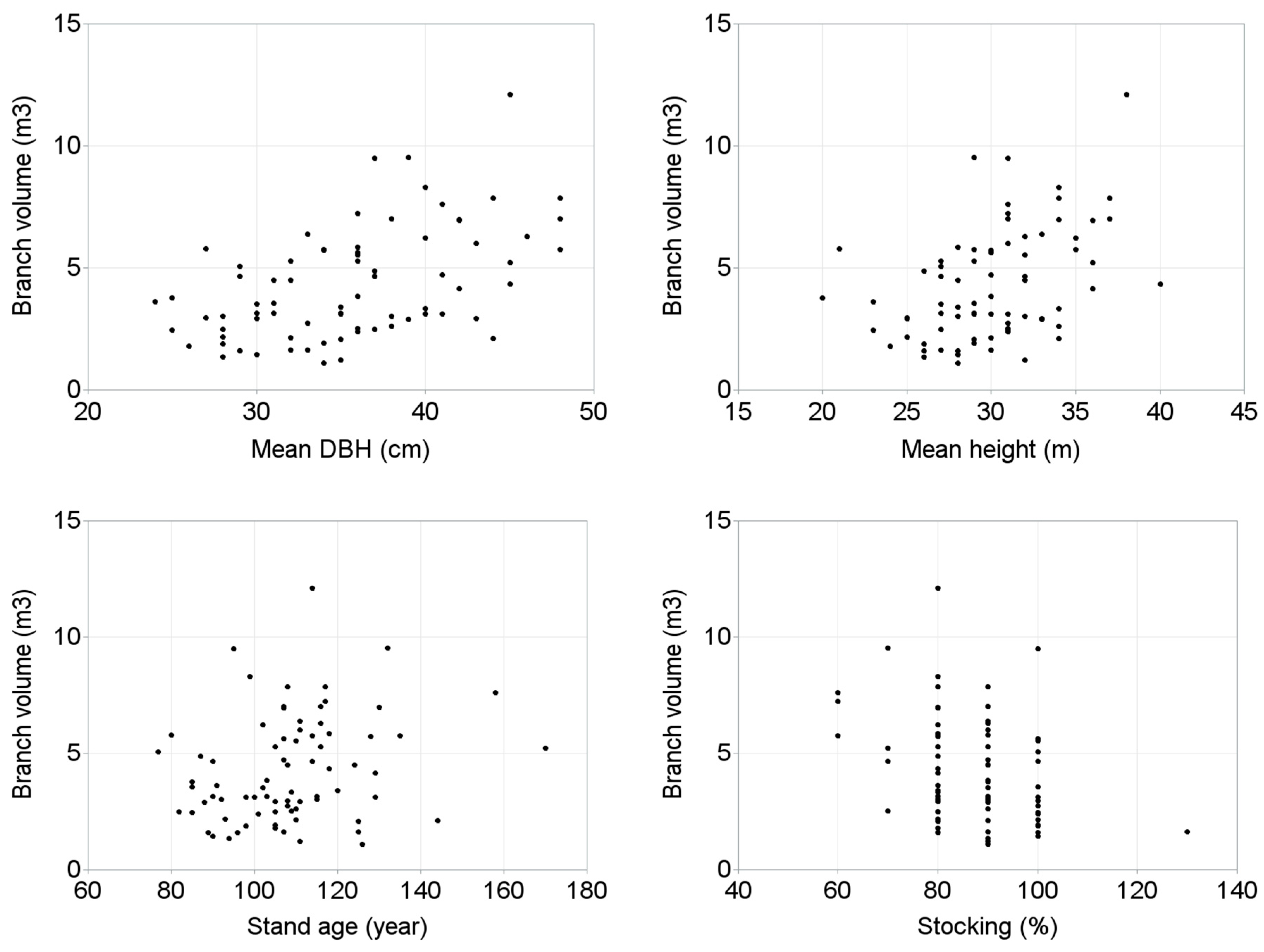
- What stand and site characteristics are correlated with branch volume?
- What is the ratio of timber volume to branch volume and how it is related to stand and site characteristics?
- Is it possible to predict the timber and branch volumes from dendrometric measurements and site characteristics?
2. Materials and Methods
Study Area
3. Sampling and Measurements
4. Data Analysis
| Site Quality Class | SQC1 | SQC2 |
| Rich | 0 | 0 |
| Medium | 1 | 0 |
| Poor | 0 | 1 |
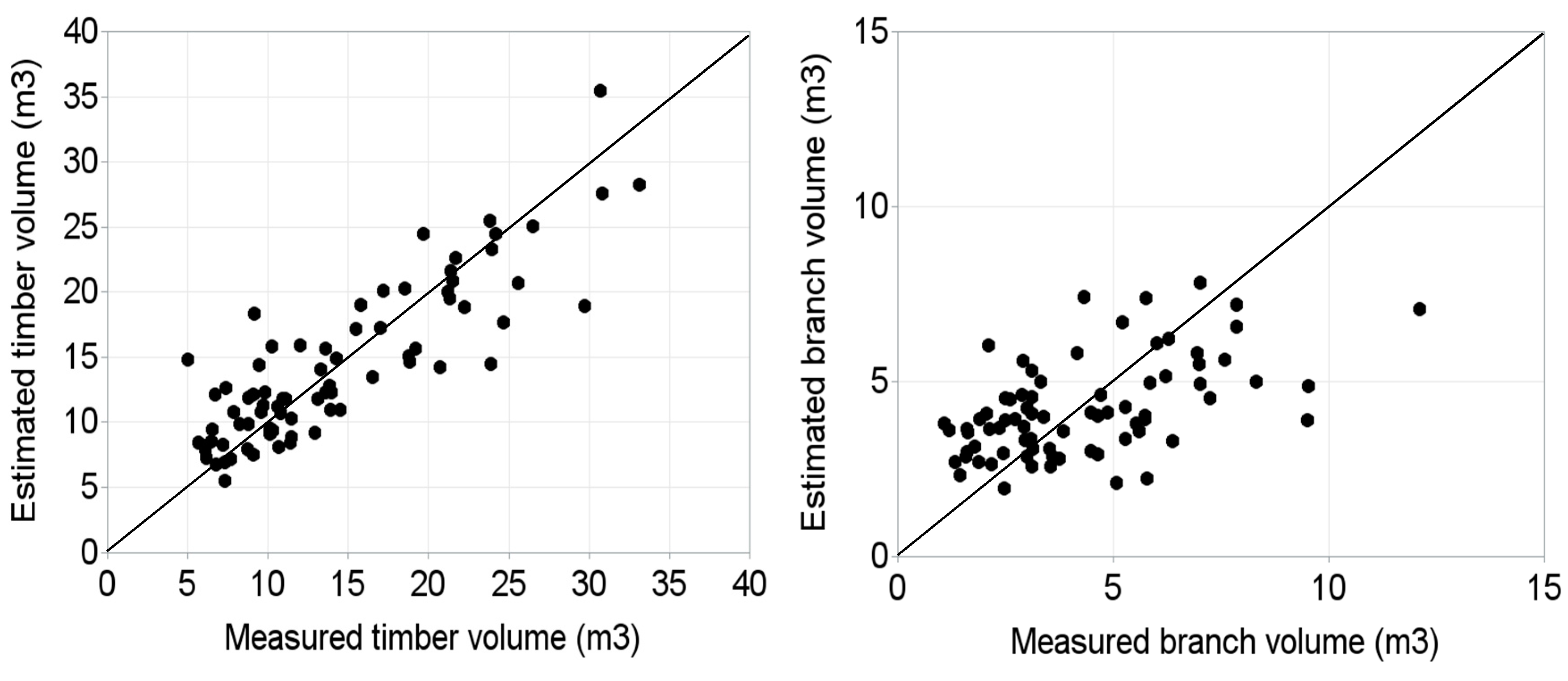
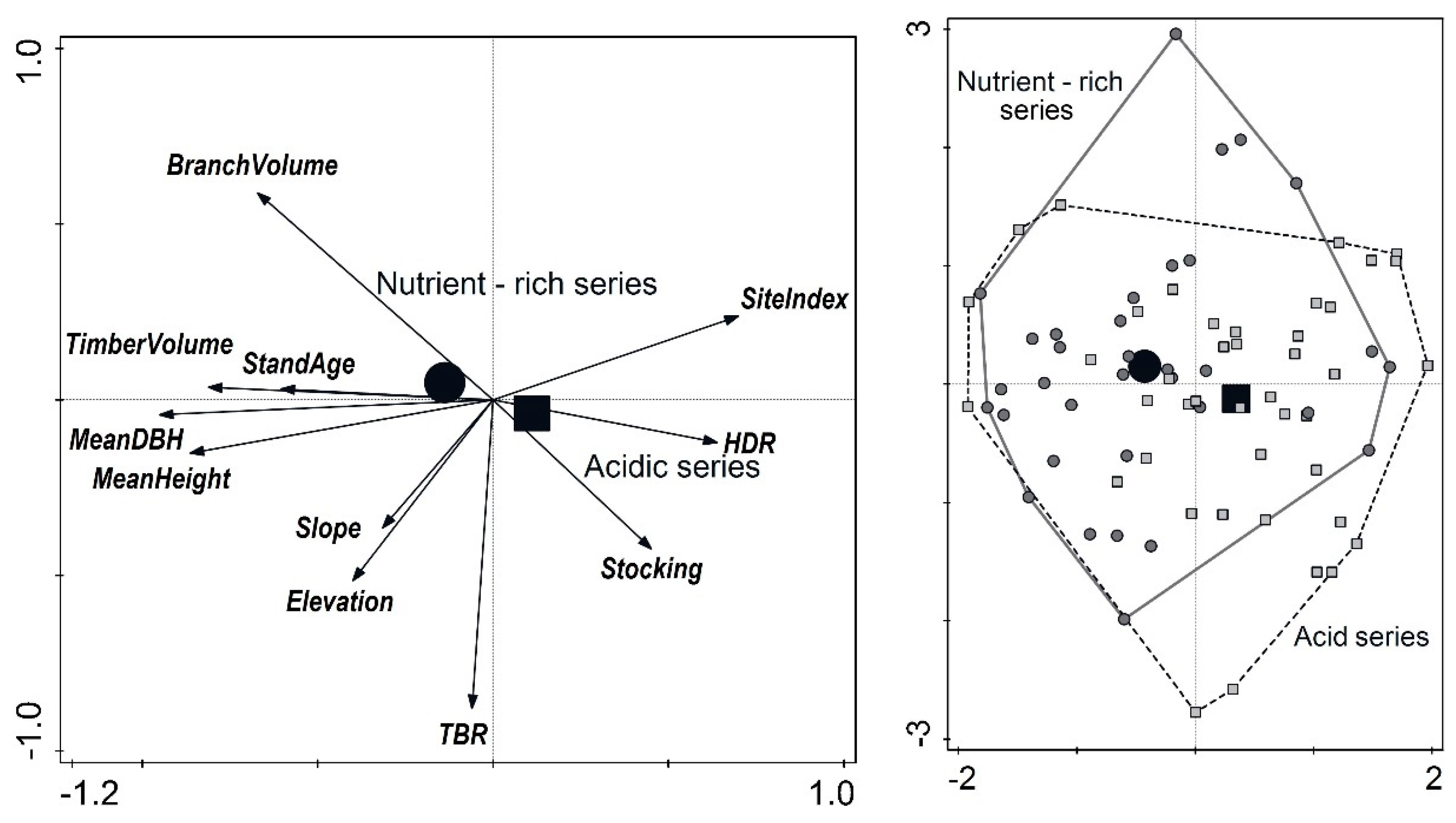
5. Results
6. Discussion
7. Conclusions
- Models for prediction of timber and branch volumes of Norway spruce were developed.
- Model described larger proportion of timber volume than branch volume.
- Factors affecting the timber-to-branch volume ratio (TBR) were analyzed and TBR was found significantly correlated with elevation and branch biomass.
- Timber volume increased with increasing slope and TBR increased with increasing elevation.
- Branch volume increased with decreasing site quality class and stand stocking.
Author Contributions
Funding
Conflicts of Interest
References
- Dymond, C.C.; Titus, B.D.; Stinson, G.; Kurz, W.A. Future quantities and spatial distribution of harvesting residue and dead wood from natural disturbances in Canada. For. Ecol. Manag. 2010, 260, 181–192. [Google Scholar] [CrossRef]
- Zhang, F.; Johnson, D.M.; Sutherland, J.W. A GIS-based method for identifying the optimal location for a facility to convert forest biomass to biofuel. Biomass Bioenergy 2011. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.-J.; Nabuurs, G.-J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Babst, F.; Poulter, B.; Trouet, V.; Tan, K.; Neuwirth, B.; Wilson, R.; Carrer, M.; Grabner, M.; Tegel, W.; Levanic, T.; et al. Site- and species-specific responses of forest growth to climate across the European continent. Glob. Ecol. Biogeogr. 2013, 22, 706–717. [Google Scholar] [CrossRef]
- Spalding-Fecher, R.; Winkler, H.; Mwakasonda, S. Energy and the World Summit on Sustainable Development: What next? Energy Policy 2005, 33, 99–112. [Google Scholar] [CrossRef]
- Al-Mansour, F.; Zuwala, J. An evaluation of biomass co-firing in Europe. Biomass Bioenergy 2010, 34, 620–629. [Google Scholar] [CrossRef]
- Chudikova, P.; Tausova, M.; Erdelyiova, K.; Taus, P. Potential of dendromass in the Slovak Republic and its actual exploitation in thermic economy. Acta Montan. Slovaca 2010, 15, 139–145. [Google Scholar]
- Schulte, A. Dendromass: Raw material of the future-trends and interdependencies. Forstarchiv 2007, 78, 59–64. [Google Scholar]
- Welfle, A.; Gilbert, P.; Thornley, P. Increasing biomass resource availability through supply chain analysis. Biomass Bioenergy 2014, 70, 249–266. [Google Scholar] [CrossRef]
- Murach, D.; Kindermann, C.; Hirschl, B.; Aretz, A.; Schneider, B.U.; Grünewald, H.; Schultze, B.; Quinkenstein, A.; Bilke, G.; Muchin, A.; et al. Dendromass, raw material of the future—background and first results of the research project DENDROM. Forstarchiv 2007, 78, 88–94. [Google Scholar]
- Bond-Lamberty, B.; Wang, C.; Gower, S.T. Net primary production and net ecosystem production of a boreal black spruce wildfire chronosequence. Glob. Chang. Biol. 2004, 10, 473–487. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Konôpka, B.; Pajtík, J.; Marušák, R. Canopy closure altered biomass allocation in young spruce stand. J. For. Sci. 2016, 61, 62–71. [Google Scholar]
- Johansson, T. Biomass Production of Norway Spruce (Picea abies (L.) Karst.) Growing on Abandoned Farmland. Silva Fenn. 1999, 33. [Google Scholar] [CrossRef]
- West, P.W. Tree and Forest Measurement; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-540-95965-6. [Google Scholar]
- Repola, J. Biomass Equations for Scots Pine and Norway Spruce in Finland. Silva Fenn. 2009, 43. [Google Scholar] [CrossRef]
- Skovsgaard, J.P.; Bald, C.; Nord-Larsen, T. Functions for biomass and basic density of stem, crown and root system of Norway spruce (Picea abies (L.) Karst.) in Denmark. Scand. J. For. Res. 2011, 26, 3–20. [Google Scholar] [CrossRef]
- Williams, H.; Messier, C.; Kneeshaw, D.D. Effects of light availability and sapling size on the growth and crown morphology of understory Douglas-fir and lodgepole pine. Can. J. For. Res. 1999, 29, 222–231. [Google Scholar] [CrossRef]
- King, J.S.; Giardina, C.P.; Pregitzer, K.S.; Friend, A.L. Biomass partitioning in red pine (Pinus resinosa) along a chronosequence in the Upper Peninsula of Michigan. Can. J. For. Res. 2007, 37, 93–102. [Google Scholar] [CrossRef]
- Schall, P.; Lödige, C.; Beck, M.; Ammer, C. Biomass allocation to roots and shoots is more sensitive to shade and drought in European beech than in Norway spruce seedlings. For. Ecol. Manag. 2012, 266, 246–253. [Google Scholar] [CrossRef]
- Dowell, R.C.; Gibbins, D.; Rhoads, J.L.; Pallardy, S. Biomass production physiology and soil carbon dynamics in short-rotation-grown Populus deltoides and P. deltoides\times P. nigra hybrids. For. Ecol. Manag. 2009, 257, 134–142. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, X.; Zhang, X.; Booth, T.; Lu, F. Root:shoot ratios across China’s forests: Forest type and climatic effects. For. Ecol. Manag. 2012, 269, 19–25. [Google Scholar] [CrossRef]
- Gower, S.T.; Reich, P.B.; Son, Y. Canopy dynamics and aboveground production of five tree species with different leaf longevities. Tree Physiol. 1993, 12, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Funct. Plant Biol. 2000, 27, 1191. [Google Scholar] [CrossRef]
- Kantola, A.; Mäkelä, A. Development of biomass proportions in Norway spruce (Picea abies [L.] Karst.). Trees Struct. Funct. 2006, 20, 111–121. [Google Scholar] [CrossRef]
- Kantola, A.; Mäkelä, A. Crown development in Norway spruce [ Picea abies (L.) Karst.]. Trees 2004, 18. [Google Scholar] [CrossRef]
- Laasasenaho, J. Taper curve and volume functions for pine, spruce and birch [Pinus sylvestris, Picea abies, Betula pendula, Betula pubescens]. Commun. Instituti For. Fenn. 1982. [Google Scholar]
- Hakkila, P. Wood density survey and dry weight tables for pine, spruce and birch stems in Finland. Metsäntutkimuslaitoksen Julk. 1979, 96, 59. [Google Scholar]
- Helmisaari, H.-S.; Makkonen, K.; Kellomäki, S.; Valtonen, E.; Mälkönen, E. Below- and above-ground biomass, production and nitrogen use in Scots pine stands in eastern Finland. For. Ecol. Manag. 2002, 165, 317–326. [Google Scholar] [CrossRef]
- Yuste, J.C.; Konopka, B.; Janssens, I.A.; Coenen, K.; Xiao, C.W.; Ceulemans, R. Contrasting net primary productivity and carbon distribution between neighboring stands of Quercus robur and Pinus sylvestris. Tree Physiol. 2005, 25, 701–712. [Google Scholar] [CrossRef]
- Wirth, C.; Schumacher, J.; Schulze, E.-D. Generic biomass functions for Norway spruce in Central Europe--a meta-analysis approach toward prediction and uncertainty estimation. Tree Physiol. 2004, 24, 121–139. [Google Scholar] [CrossRef]
- Avery, T.E.; Burkhart, H.E. Forest Measurements, Fifth ed.; Waveland Press: Long Grove, IL, USA, 2015; ISBN 9781478629740. [Google Scholar]
- Benson, R.E.; Johnston, C.M. Logging Residues under Different Stand and Harvesting Conditions, Rocky Mountains; Intermountain Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture: Ogden, UT, USA, 1976.
- Sačkov, I.; Natov, P.; Messingerová, V. On-line Calculator of Volume and Price extraction of above the Ground Dendromass from Forest Stand. Lesn. Časopis 2008, 54, 75–85. [Google Scholar]
- Benson, R.E.; Schlieter, J.A. Logging Residues in Principal Forest Types of the Northern Rocky Mountains; Intermountain Forest and Range Experihent Station, U.S. Department of Agriculture Forest Service: Ogden, UT, USA, 1980.
- Schmidt-Vogt, H. The Spruce [Picea]. In Geographical Distribution, Morphology, Ecology, and Forest Communities; Paul Parey: Hamburg, Germany, 1977. [Google Scholar]
- Litton, C.M.; Raich, J.W.; Ryan, M.G. Carbon allocation in forest ecosystems. Glob. Chang. Biol. 2007, 13, 2089–2109. [Google Scholar] [CrossRef]
- Brunner, I.; Bakker, M.R.; Björk, R.G.; Hirano, Y.; Lukac, M.; Aranda, X.; Børja, I.; Eldhuset, T.D.; Helmisaari, H.S.; Jourdan, C.; et al. Fine-root turnover rates of European forests revisited: An analysis of data from sequential coring and ingrowth cores. Plant Soil 2013, 362, 357–372. [Google Scholar] [CrossRef]
- Cienciala, E.; Černý, M.; Tatarinov, F.; Apltauer, J.; Exnerová, Z. Biomass functions applicable to Scots pine. Trees 2006, 20, 483–495. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Bellos, P.; Hochbichler, E.; Lexer, M.J. Testing generalized allometries in allocation modeling within an individual-based simulation framework. Trees 2010, 24, 139–150. [Google Scholar] [CrossRef]
- Pajtík, J.; Konôpka, B.; Lukac, M. Biomass functions and expansion factors in young Norway spruce (Picea abies [L.] Karst) trees. For. Ecol. Manag. 2008, 256, 1096–1103. [Google Scholar] [CrossRef]
- Dutca, I.; Abrudan, I.; Stancioiu, P.; Viorel, B. Biomass conversion and expansion factors for young Norway spruce (Picea abies (L.) Karst.) trees planted on non-forest lands in Eastern Carpathians. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38. [Google Scholar]
- Konôpka, B.; Pajtík, J.; Moravčík, M.; Lukac, M. Biomass partitioning and growth efficiency in four naturally regenerated forest tree species. Basic Appl. Ecol. 2010, 11, 234–243. [Google Scholar] [CrossRef]
- Marklund, L.G. Biomass Functions for Norway Spruce (Picea abies (L.) Karst.) in Sweden [Biomass Determination, Dry Weight]; Department of Forest Survey, Swedish University of Agricultural Sciences: Umeå, Sweden, 1987. [Google Scholar]
- Marklund, L.G. Biomass Functions for Pine, Norway Spruce, and Birch in Sweden; Department for Forest Survey Report 45; Swedish University of Agricultural Sciences: Umeå, Sweden, 1988. [Google Scholar]
- Petráš, R.; Pajtík, J. Sústava česko-slovenských objemových tabuliek drevín [Volume equations for tree species in Czechoslovakia]. Lesn. časopis 1991, 37, 49–56. [Google Scholar]
- Ter-Mikaelian, M.T.; Korzukhin, M.D. Biomass equations for sixty-five North American tree species. For. Ecol. Manag. 1997, 97, 1–24. [Google Scholar] [CrossRef]
- Lehtonen, A.; Cienciala, E.; Tatarinov, F.; Mäkipää, R. Uncertainty estimation of biomass expansion factors for Norway spruce in the Czech Republic Uncertainty estimation of biomass expansion factors for Norway spruce in the Uncertainty estimation of biomass expansion factors for Norway spruce in the Czech Republic. Ann. For. Sci 2007, 64, 133–140. [Google Scholar]
- Saidur, R.; Abdelaziz, E.A.; Demirbas, A.; Hossain, M.S.; Mekhilef, S. A review on biomass as a fuel for boilers. Renew. Sustain. Energy Rev. 2011, 15, 2262–2289. [Google Scholar] [CrossRef]
- Štorek, V.; Štícha, V.; Skoupý, A.; Kondělka, P.; Mikulenčák, J. Stanovování množství zbytkové dendromasy na konkrétní pracoviště—těžební prvek v porostní skupině: Certifikovaná metodika; Česká zemědělská univerzita v Praze (Determining the amount of residual dendromass in a specific logging place in the part of a stand); Foresta SF, a.s.: Vsetín, Czech Republic, 2017; ISBN 8074581047. [Google Scholar]
- Tolasz, R.; Míková, T.; Valeriánová, A.; Voženílek, V. Climate atlas of Czechia. Czech Hydrometeorological Institute; Czech Hydrometeorological Institute Prague: Olomouc, Czech Republic, 2007. [Google Scholar]
- Kozák, J.; Němeček, J. Atlas půd České Republiky; MZe ČR ve spolupráci s ČZU: Prague, Czech Republic, 2009; ISBN 9788021318823. [Google Scholar]
- Viewegh, K.; Kusbach, J.V.A.; Mikeska, M. Czech forest ecosystem classification. J. For. Sci. 2003, 49, 85–93. [Google Scholar] [CrossRef]
- Černý, M.; Pařez, J. Tabulky a polynomy pro výpočet objemu kulatiny bez kůry; Ministerstvo zemědělství ČR: Praha, Czech Republic, 1995.
- BioNorm II Prenormative Work on Sampling and Testing of Solid Biofuels for the Development of Quality Management; Publications Office of the European Union: Luxembourg, 2004.
- Požgaj, A.; Chovanec, D.; Kurjatko, S.; Babiak, M. Štruktúra a vlastnosti dreva; Príroda: Bratislava, Slovakia, 1993; ISBN 80-07-00600-1. [Google Scholar]
- Agency, E.E. EU-DEM v1.1. Available online: https://land.copernicus.eu/imagery-in-situ/eu-dem/eu-dem-v1.1?tab=mapview (accessed on 5 March 2018).
- Bates, D.; Watts, D. Nonlinear Regression. Analysis and its Applications; Wiley: New York, NY, USA, 1988. [Google Scholar]
- Sharma, R.P.; Brunner, A.; Eid, T. Site index prediction from site and climate variables for Norway spruce and Scots pine in Norway. Scand. J. For. Res. 2012, 27, 619–636. [Google Scholar] [CrossRef]
- Sharma, R.; Vacek, Z.; Vacek, S. Generalized Nonlinear Mixed-Effects Individual Tree Crown Ratio Models for Norway Spruce and European Beech. Forests 2018, 9, 555. [Google Scholar] [CrossRef]
- Montgomery, D.; Peck, E.A.; Vining, G. Introduction to Linear Regression Analysis; Wiley: New York, NY, USA, 2001. [Google Scholar]
- SAS Institute Inc. SAS/ETS1 9.1.3 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using Canoco 5; Cambridge University Press: Cambridge, UK, 2014; ISBN 9781107694408. [Google Scholar]
- Spiecker, H. Growth Trends in European Forests Studies from 12 Countries; Springer Science & Business Media: Berlin, Germany, 1996; ISBN 978-3-540-61460-9. [Google Scholar]
- Mäkinen, H.; Nöjd, P.; Kahle, H.-P.; Neumann, U.; Tveite, B.; Mielikäinen, K.; Röhle, H.; Spiecker, H. Radial growth variation of Norway spruce (Picea abies (L.) Karst.) across latitudinal and altitudinal gradients in central and northern Europe. For. Ecol. Manag. 2002, 171, 243–259. [Google Scholar] [CrossRef]
- Mäkinen, H.; Nöjd, P.; Kahle, H.-P.; Neumann, U.; Tveite, B.; Mielikäinen, K.; Röhle, H.; Spiecker, H. Large-scale climatic variability and radial increment variation of Picea abies (L.) Karst. in central and northern Europe. Trees 2003, 17, 173–184. [Google Scholar]
- Dittmar, C.; Eißing, T.; Rothe, A. Elevation-specific tree-ring chronologies of Norway spruce and Silver fir in Southern Germany. Dendrochronologia 2012, 30, 73–83. [Google Scholar] [CrossRef]
- Leal, S.; Melvin, T.M.; Grabner, M.; Wimmer, R.; Briffa, K.R. Tree-ring growth variability in the Austrian Alps: The influence of site, altitude, tree species and climate. Boreas 2007, 36, 426–440. [Google Scholar] [CrossRef]
- Hartl, C.; Zang, C.; Dittmar, C.; Esper, J.; Göttlein, A.; Rothe, A. Vulnerability of Norway spruce to climate change in mountain forests of the European Alps. Clim. Res. 2014, 60, 119–132. [Google Scholar] [CrossRef]
- Hartl-Meier, C.; Dittmar, C.; Zang, C.; Rothe, A. Mountain forest growth response to climate change in the Northern Limestone Alps. Trees 2014, 28, 819–829. [Google Scholar] [CrossRef]
- Wilson, R.J.S.; Wilson, R.J.S.; Hopfmueller, M. Dendrochronological investigations of Norway spruce along an elevational transect in the Bavarian Forest, Germany. Dendrochronologia 2001, 1, 67–79. [Google Scholar]
- Čejková, A.; Kolář, T. Extreme radial growth reaction of norway spruce along an altitudinal gradient in the šumava mountains. Geochronometria 2009, 33, 41–47. [Google Scholar] [CrossRef]
- Bednarz, Z.; Jaroszeewicz, B.; Ptak, J.; Szwagrzyk, J. Dendrochronology of Norway Norway spruce (Picea abies (L. Karst) in the Babia Gora National Park, Poland. Dendrochronologia 1999, 16, 45–55. [Google Scholar]
- Kaczka, R.J.; Czajka, B.; Łajczak, A. The tree-ring growth responses to climate in the timberline ecotone of Babia Góra Mountain. Geogr. Pol. 2015, 88, 163–176. [Google Scholar] [CrossRef]
- Savva, Y.; Oleksyn, J.; Reich, P.; Tjoelker, M.; Vaganov, E.A.; Modrzynski, J. Interannual growth response of Norway spruce to climate along an altitudinal gradient in the Tatra Mountains, Poland. Trees 2006, 20, 735–746. [Google Scholar] [CrossRef]
- Büntgen, U.; Frank, D.C.; Kaczka, R.J.; Verstege, A.; Zwijacz-Kozica, T.; Esper, J. Growth responses to climate in a multi-species tree-ring network in the Western Carpathian Tatra Mountains, Poland and Slovakia. Tree Physiol. 2007, 27, 689–702. [Google Scholar] [CrossRef]
- Kaczka, R.J.; Büntgen, U. Spatial autocorrelation and growth/climate response of a high elevation norway spruce network along the Carpathian arc. TRACE 2006, 6, 103–112. [Google Scholar]
- Sidor, C.G.; Popa, I.; Vlad, R.; Cherubini, P. Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carpathians (Romania). Trees 2015, 29, 985–997. [Google Scholar] [CrossRef]
- Ponocná, T.; Spyt, B.; Kaczka, R.; Büntgen, U.; Treml, V. Growth trends and climate responses of Norway spruce along elevational gradients in East-Central Europe. Trees 2016, 30, 1633–1646. [Google Scholar] [CrossRef]
- Vacek, S.; Chroust, L.; Souček, J. Wood production analysis of autochthonous Norway norway spruce forest stand. Lesnictví 1994, 10, 457–469. [Google Scholar]
- Kuuluvainen, T. Relationships between crown projected area and components of above-ground biomass in Norway spruce trees in even-aged stands: Empirical results and their interpretation. For. Ecol. Manag. 1991, 40, 243–260. [Google Scholar] [CrossRef]
- Vacek, S.; Hejcmanová, P.; Hejcman, M. Vegetative reproduction of Picea abies by artificial layering at the ecotone of the alpine timberline in the Giant (Krkonoše) Mountains, Czech Republic. For. Ecol. Manag. 2012, 263, 199–207. [Google Scholar] [CrossRef]
- Vacek, S. Morfologická proměnlivost autochtonních smrkových populací v Krkonoších. (Morphological variability of autochthonous Norway norway spruce populations in the Krkonoše Mts). Lesnictví 1983, 29, 265–284. [Google Scholar]
- Schuster, R.; Oberhuber, W. Drought sensitivity of three co-occurring conifers within a dry inner Alpine environment. Trees (Berl. West) 2013, 27, 61–69. [Google Scholar] [CrossRef]
- Hlásny, T.; Barcza, Z.; Fabrika, M.; Balázs, B.; Churkina, G.; Pajtík, J.; Turčáni, M. Climate change impacts on growth and carbon balance of forests in Central Europe. Clim. Res. 2011, 47, 219–236. [Google Scholar] [CrossRef]
- King, G.M.; Gugerli, F.; Fonti, P.; Frank, D.C. Tree growth response along an elevational gradient: Climate or genetics? Oecologia 2013, 173, 1587–1600. [Google Scholar] [CrossRef]
- Biondi, F.; Qeadan, F. A Theory-Driven Approach to Tree-Ring Standardization: Defining the Biological Trend from Expected Basal Area Increment. Tree-Ring Res. 2008, 64, 81–96. [Google Scholar] [CrossRef]
- Büntgen, U.; Tegel, W.; Heussner, K.; Hofmann, J.; Kontic, R.; Kyncl, T.; Cook, E. Effects of sample size in dendroclimatology. Clim. Res. 2012, 53, 263–269. [Google Scholar] [CrossRef]
- Babst, F.; Bouriaud, O.; Alexander, R.; Trouet, V.; Frank, D. Toward consistent measurements of carbon accumulation: A multi-site assessment of biomass and basal area increment across Europe. Dendrochronologia 2014, 32, 153–161. [Google Scholar] [CrossRef]
- Čihák, T.; Vejpustková, M. Parametrování alometrických rovnic pro kvantifikaci nadzemní biomasy smrku smrkového (Picea abies (L.) H. Karst.) V České republice. J. For. Sci. 2018, 64, 108–117. [Google Scholar]
- Ilomäki, S.; Nikinmaa, E.; Mäkelä, A. Crown rise due to competition drives biomass allocation in silver birch. Can. J. For. Res. 2003, 33, 2395–2404. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G. Crown allometry and growing space efficiency of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica L.) in pure and mixed stands. Plant Biol. 2005, 7, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Schröter, M.; Härdtle, W.; von Oheimb, G. Crown plasticity and neighborhood interactions of European beech (Fagus sylvatica L.) in an old-growth forest. Eur. J. For. Res. 2012, 131, 787–798. [Google Scholar] [CrossRef]
- Bulušek, D.; Vacek, Z.; Vacek, S.; Král, J.; Bílek, L.; Králíček, I. Spatial pattern of relict beech (Fagus sylvatica L.) forests in the Sudetes of the Czech Republic and Poland. J. For. Sci. 2016, 62, 293–305. [Google Scholar] [CrossRef]
- Assman, E. The Principles of Forest Yield Study; Pergamon Press: Oxford, UK, 1970. [Google Scholar]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Individual tree crown width models for Norway spruce and European beech in Czech Republic. For. Ecol. Manag. 2016, 366, 208–220. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Mean | Standard Deviation | Minimum | Maximum |
|---|---|---|---|---|
| Timber volume (m3) | 14.43 | 6.98 | 5.03 | 33.11 |
| Branch volume (m3) | 4.27 | 2.27 | 1.08 | 12.09 |
| Timber-to-branch volume ratio | 3.72 | 1.43 | 1.27 | 9.57 |
| Branch weight (kg) | 2.75 | 1.53 | 0.67 | 8.70 |
| Mean DBH (cm) | 35.49 | 6.06 | 24.00 | 48.00 |
| Mean height (m) | 30.01 | 3.88 | 20.00 | 40.00 |
| Elevation (m) | 726.8 | 172.5 | 380 | 1040 |
| Slope (degree) | 16.17 | 10.94 | 0.72 | 45.69 |
| Stand age (year) | 16.17 | 10.94 | 0.72 | 45.70 |
| Stand stocking (%) | 86 | 11.19 | 60 | 130 |
| Timber Volume | Branch Volume | Mean DBH | Mean Height | Elevation | Slope | Stand Age | Aspect | Stocking | |
|---|---|---|---|---|---|---|---|---|---|
| Branch volume | 0.792 (0.0001) | ||||||||
| Mean DBH | 0.618 (0.0001) | 0.5152 (0.0001) | |||||||
| Mean height | 0.579 (0.0001) | 0.426 (0.0001) | 0.8554 (0.0001) | ||||||
| Elevation | 0.145 (0.2129) | −0.0158 (0.8922) | 0.3459 (0.0022) | 0.173 (0.1350) | |||||
| Slope | 0.392 (0.0005) | 0.1979 (0.0866) | 0.2269 (0.0487) | 0.2925 (0.0103) | 0.2046 (0.0762) | ||||
| Stand age | 0.285 (0.0124) | 0.2357 (0.0403) | 0.5765 (0.0001) | 0.4931 (0.0001) | 0.2691 (0.0188) | 0.0187 (0.8724) | |||
| Aspect | 0.172 (0.1382) | −0.193 (0.0948) | −0.0748 (0.5205) | −0.132 (0.2580) | 0.199 (0.0844) | −0.025 (0.8275) | −0.018 (0.8761) | ||
| Stocking | −0.23432 (0.0416) | −0.33419 (0.0032) | −0.35116 (0.0019) | −0.28117 (0.0139) | −0.00098 (0.9933) | −0.07151 (0.5393) | −0.45303 (<0.0001) | 0.12383 (0.2865) | |
| Timber-to-branch volume ratio (TBR) | 0.06686 (0.5661) | −0.44449 (0.0001) | 0.04532 (0.6975) | 0.14596 (0.2083) | 0.29713 (0.0091) | 0.21281 (0.0649) | 0.11116 (0.3391) | −0.03423 (0.7691) | 0.1709 (0.1398) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štícha, V.; Sharma, R.P.; Vacek, Z.; Vacek, S.; Nuhlíček, O. Timber and Branch Volume Prediction: Effects of Stand and Site Characteristics on Dendromass and Timber-To-Branch Volume Ratio of Norway Spruce in Managed Forests. Forests 2019, 10, 144. https://0-doi-org.brum.beds.ac.uk/10.3390/f10020144
Štícha V, Sharma RP, Vacek Z, Vacek S, Nuhlíček O. Timber and Branch Volume Prediction: Effects of Stand and Site Characteristics on Dendromass and Timber-To-Branch Volume Ratio of Norway Spruce in Managed Forests. Forests. 2019; 10(2):144. https://0-doi-org.brum.beds.ac.uk/10.3390/f10020144
Chicago/Turabian StyleŠtícha, Václav, Ram P. Sharma, Zdeněk Vacek, Stanislav Vacek, and Ondřej Nuhlíček. 2019. "Timber and Branch Volume Prediction: Effects of Stand and Site Characteristics on Dendromass and Timber-To-Branch Volume Ratio of Norway Spruce in Managed Forests" Forests 10, no. 2: 144. https://0-doi-org.brum.beds.ac.uk/10.3390/f10020144