The Colour of Tropical Woods Influenced by Brown Rot


Technical University in Zvolen, Faculty of Wood Sciences and Technology, T. G. Masaryka 24, Zvolen, SK 96001, Slovakia
*
Author to whom correspondence should be addressed.
Forests 2019, 10(4), 322; https://0-doi-org.brum.beds.ac.uk/10.3390/f10040322
Submission received: 26 February 2019
/
Revised: 31 March 2019
/
Accepted: 2 April 2019
/
Published: 10 April 2019
(This article belongs to the Special Issue Wood Properties and Processing)
Abstract
:Interesting aesthetic properties of tropical woods, like surface texture and colour, are rarely impaired due to weathering, rotting and other degradation processes. This study analyses the colour of 21 tropical woods before and after six weeks of intentional attack by the brown-rot fungus Coniophora puteana. The CIEL*a*b* colour system was applied for measuring the lightness, redness and yellowness, and from these parameters the hue tone angle and colour saturation were calculated. Lighter tropical woods tended to appear a less red and a more yellow, and had a greater hue tone angle. However, for the original woods was not found dependence between the lightness and colour saturation. Tropical woods at attack by C. puteana lost a weight from 0.08% to 6.48%. The lightest and moderately light species—like okoumé, iroko, ovengol and sapelli—significantly darkened, while the darkest species—wengé and ipé—significantly lightened. The majority of tropical woods obtained a brighter shade of yellow, typically wengé, okoumé and blue gum, while some of them also a brighter shade of green, typically sapelli, padouk and macaranduba. C. puteana specifically affected the hue tone angle and colour saturation of tested tropical woods, but without an apparent changing the tendency of these colour parameters to lightness. The total colour difference of tested tropical woods significantly increased in connection with changes of their lightness (ΔE*ab = 5.92 − 0.50·ΔL*; R2 = 0.37), but it was not influenced by the red and yellow tint changes, and weight losses.
1. Introduction
The surface appearance of wood is often evaluated by examining its texture, roughness, and colour [1,2,3]. The colour of an individual wood species is predetermined by the type and amount of extractives, by the surface roughness and moisture, and by the direction of light irradiation [4].
The CIE 1976 L*a*b* colour system classifies the temperate and tropical wood species into the positive octant with the lightness (L*) from 20 to 90, the redness index (+ a*) from 0 to 20, and the yellowness index (+ b*) from 10 to 30 [1,5]. The CIE 1976 L*a*b* colour system also allows visualisation of the cylindrical parameters of wood, the colour saturation—chromaticity (C*ab) and the hue tone angle (hab) [6]. The tropical wood species occupy a much greater portion of the colour space in comparison with temperate (for example European) species [7,8].
Within a defined wood species, the colour variations can be influenced by more factors, mainly by its chemical and anatomical structure [9] and specific genetic parameters [10], and also by environmental conditions at growth [11,12], atmospheric effects at exposure in exteriors or interiors [13,14], and biodeterioration processes [15,16,17].
Wooden products having a higher moisture content—usually above 20%–30%—are no rarely damaged by biodeterioration processes in presence of wood decaying fungi, staining fungi, moulds, or bacteria.
Generally, brown-rot fungi cause firstly yellowing and gradually browning of woods in a connection with decomposition of whiter hemicelluloses and cellulose, while darker lignin is less evidently damaged [18,19]. Wood extractives, which give to different wood species a characteristic colour, are specifically resistant to individual species of brown-rot fungi [20,21,22]. Therefore, the accidental or deliberate exposures of wood products to brown-rot fungi can lead to typical changes in their colour and aesthetic parameters.
The rotting of damp wooden products is common not only outdoors, but also inside of buildings. In interiors, the rotting of wood is first of all caused by brown-rot fungi [23,24,25]. The genus Coniophora comprises about 20 species frequently occurring in buildings. Gabriel and Švec [26] listed the species abundance of seven indoor wood decay basidiomycetes reported in Europe, when the brown-rot fungi Serpula lacrymans and Coniophora puteana (C. puteana) were most frequent. C. puteana causes—already in the early period of wood attack—a disruption of linkages between hemicelluloses and lignin, decomposition of polysaccharides, while lignin is oxidative modified and partly damaged [27]. Rot of wood with C. puteana is not usually homogenous and it’s specific parts can be more damaged [15].
In the literature, it is also noted that wood-inhabiting fungi are able add colour to wood due to a pigment residues left by fungi in wood (called as spalting) [17,28,29,30,31,32,33,34]. Spalting occurs in growing tree in form of zone lines formation or pigmentation. Also bleaching is marked as kinds of wood spalting [17,29,31]. Bleaching is caused by the breakdown of coloured lignin from the wood cell wall, generally by fungi classified as white-rotting, which results in a lightening of the natural wood colour and structural integrity of the wood. A lightening in colour can also be due to a build-up of white mycelium [18,20,35]. Wood with zone lines (thin and winding lines of dark melanin) are formed due to inter- or intra- fungal antagonism for example a pairing of the white-rot fungi Trametes versicolor/Bjerkadera adusta or by solitary isolates of Xylaria polymorpha ascomycete that causes soft rot [31]. Also, the brown-rot fungus Fistulina hepatica stains oaks and some other woods from light gold, yellow brown to reddish brown shades [36,37]. Pigment-type spalting fungi are a select group of soft-rotting ascomycetes with extracellular pigments production into wood, for example fungi from genera Chlorocibolia, Ceratocystis, Ophiostoma, Scytalidium and others [32,33]. Pigment penetration into wood depends on moisture content, the digestive capabilities of the fungus, the permeability of the wood structure, differences between heartwood and sapwood, and the type of pigment produced [29]. Mold fungi, such as Trichoderma spp., are not considered to be spalting fungi, as their hyphae do not colonize the wood internally and they do not produce the enzymes necessary to digest the wood cell wall components.
From several tropical woods are manufactured various products for interiors, for example, furniture, flooring, stairs, windows, doors, claddings or structural elements—using usually massive, glued timbers, veneers or plywood [38]. For more of these products, both strength and aesthetic are important. The interesting aesthetic and specific colours of the individual tropical wood species, existing in the natural state as well as in the primarily fungal-pigmented state, can be changed or even worsened at additional rotting processes. In interiors, the degree and range of rotting is influenced mainly by: (1) the natural durability of wood to decaying fungi (e.g., [39] classifies woods into five classes of durability), and (2) the enough moisture of wood above 20%, depending on presence of condensed, capillary, plumbing or rain water.
The aim of this study was to determine the colour changes of 21 tropical woods when exposed to the brown-rot fungus C. puteana, which can cause an important deterioration of wood products in the interiors of buildings.
2. Materials and Methods
2.1. Woods and Specimens
Twenty-one tropical woods, in a form of naturally dried and conditioned boards with a moisture content of 13 ± 2.5%, was bought from the trading company JAF Holz, Ltd., Slovakia (Table 1).
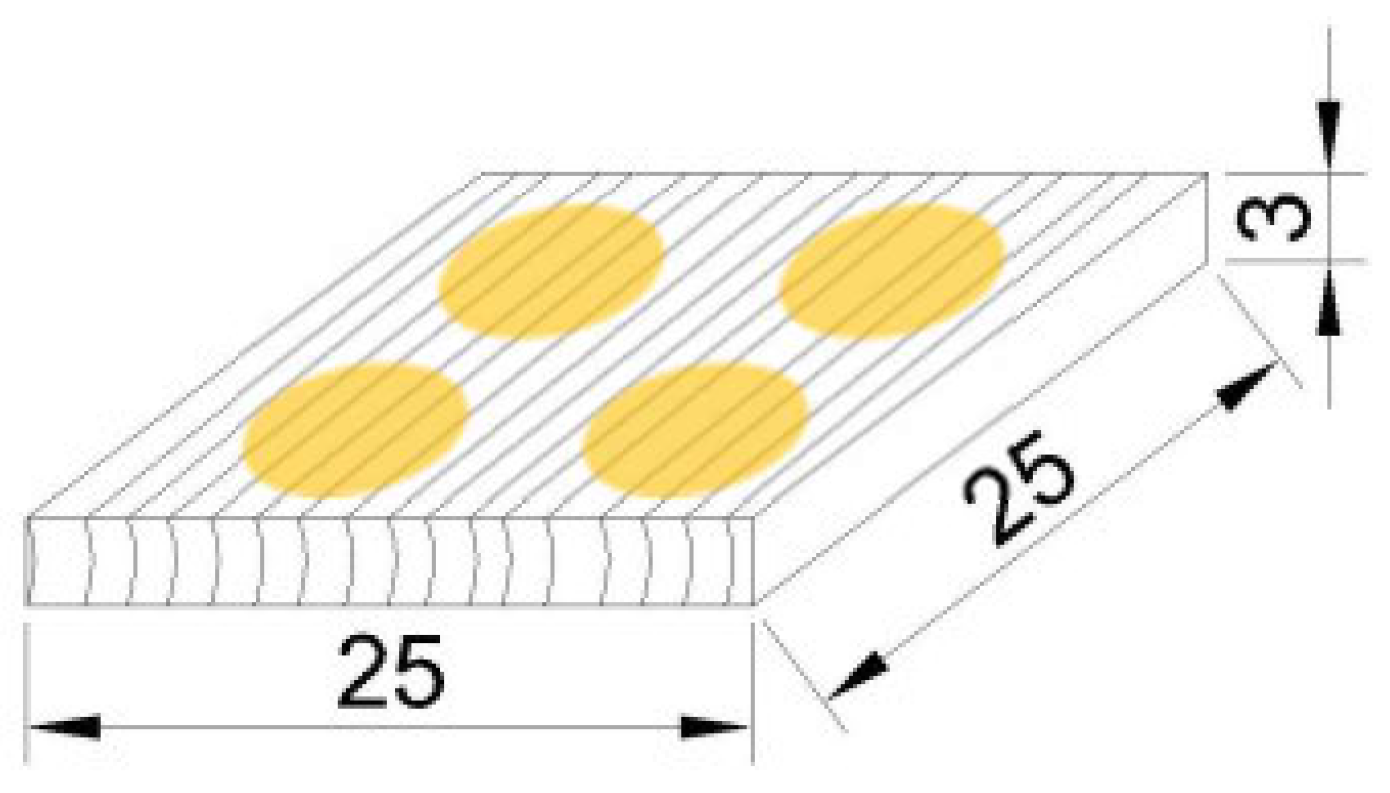
From the heart-zone of each wood species were prepared and tested four specimens 25 mm × 25 mm × 3 mm (longitudinal × radial × tangential)—without biological damage, knots or other defects. Before the fungal attack, the top surfaces of specimens were sanded along fibres using 240-grit sandpaper. Subsequently, the specimens were conditioned on a moisture content of 12 ± 1%, weighted with an accuracy of 0.001 g, sterilized with 30 W germicidal lamp (Chirana, Slovakia) at a temperature of 22 ± 2 °C per 20 min for each side, and finally their top surfaces submitted in sterilized room to colour analyses (point 2.3).
After fungal attack (point 2.2), the specimens were carefully cleaned from surface fungal mycelia, slowly dried in a laboratory, conditioned on a moisture content of 12 ± 1%, weighted with an accuracy of 0.001 g, the top surfaces sanded along fibres with 240-grit sandpaper, and finally the top surfaces again submitted to colour analyses (point 2.3).
At sanding of the top surfaces of tested specimens, performed before and after fungal attack, in both cases the thickness of specimens declined about approximately 10 micrometers.
2.2. Fungal Attack of Woods
The specimens of tropical woods were exposed to the brown-rot fungus Coniophora puteana (Schumacher ex Freist) Karsten, strain BAM Ebw. 15 (Bundesanttalt für Materialforshung und—prüfung, Berlin) in glass Petri dishes with a diameter of 100 mm. Two replicates of the same wood species were placed into one dish on plastic mats under which a fungal mycelium was already grown on an autoclave sterilized and solidified 3–4 mm thick layer of 4.5 wt.% malt agar medium (HiMedia, Ltd., India). Fungal attacks lasted 6 weeks at a temperature of 24 ± 2 °C and a relative humidity of 90 ± 5%.
The weight loss Δm (%) of specimens was calculated from their weights in conditioned state before and after fungal attack. This method may have caused some inaccuracies in Δm (due to sorption hysteresis and differential water sorption by healthy and rotten woods), but it was preferred to the method evaluating specimens in absolute dry state when colour change could be manifested at drying temperature of 103 °C.
2.3. Colour Analyses of Woods
The colour analyses were performed for each specimen before and after fungal attack on the sanded top surfaces (point 2.1) in the same four places (Figure 1). The colour measurements were performed with the Color Reader CR-10 (Konica Minolta, Japan), having a CIE 10° standard observer, CIE standard illuminate D65, sensor head with a diameter of 8 mm (i.e., the measuring area was 50 mm2), and a detector with 6 silicon photocells.
The colourimetric parameters of each specimen were analysed according to the CIE 1976 L*a*b* colour system. A larger value of L*, a*, or b* means a lighter, redder, or yellower colour, respectively.
Based on the L*, a*, and b* colour coordinates, following the colour saturation—chromacity C*ab and the hue tone angle hab were calculated according to the [42] by Equations (1) and (2):
From the relative colour changes ΔL∗, Δa∗, and Δb∗, namely differences between colour coordinates of the fungal-attacked and the original wood specimens, the total colour difference ΔE*ab was calculated by Equation (3) [42]:
Selected colourimetric parameters of tropical woods determined in the original state (a*, b*, C*ab, hab), as well as in the fungal-attacked state (a*F, b*F, C*ab F, hab F), were finally analysed in relation to their lightness (L∗ or L∗F) by linear correlations using Equation (4):
Equation (4) was used as well as for searching relations between the ΔE*ab ∗ and the relative colour changes ΔL∗, Δa∗, Δb∗, and the weight loss Δm, respectively.
Conversion of average value of the colourimetric coordinates L*, a* and b* of the fungal-attacked and the original wood specimens was generated their colour (Pantone) swatch.
The t-test statistically analysed the colour changes of individual tropical woods due to the brown-rot fungus C. puteana.
3. Results and Discussion
3.1. Colour of Original Tropical Woods
Table 2 documents colour characteristics of 21 tropical woods in the original state—a visualisation (colour and surface structure), and the colourimetric parameters L*, a*, b*, C*ab and hab.
The studied tropical woods differed mainly in the lightness L*, which ranged between 34.88 (“very dark” wengé), 43.78 (“dark” padouk), 63.58 (“light” iroko) and 75.36 (“very light” okoumé). Achieved results are in an accordance with similar works disserting specific colour and texture characteristics of selected tropical woods [7,40,43].
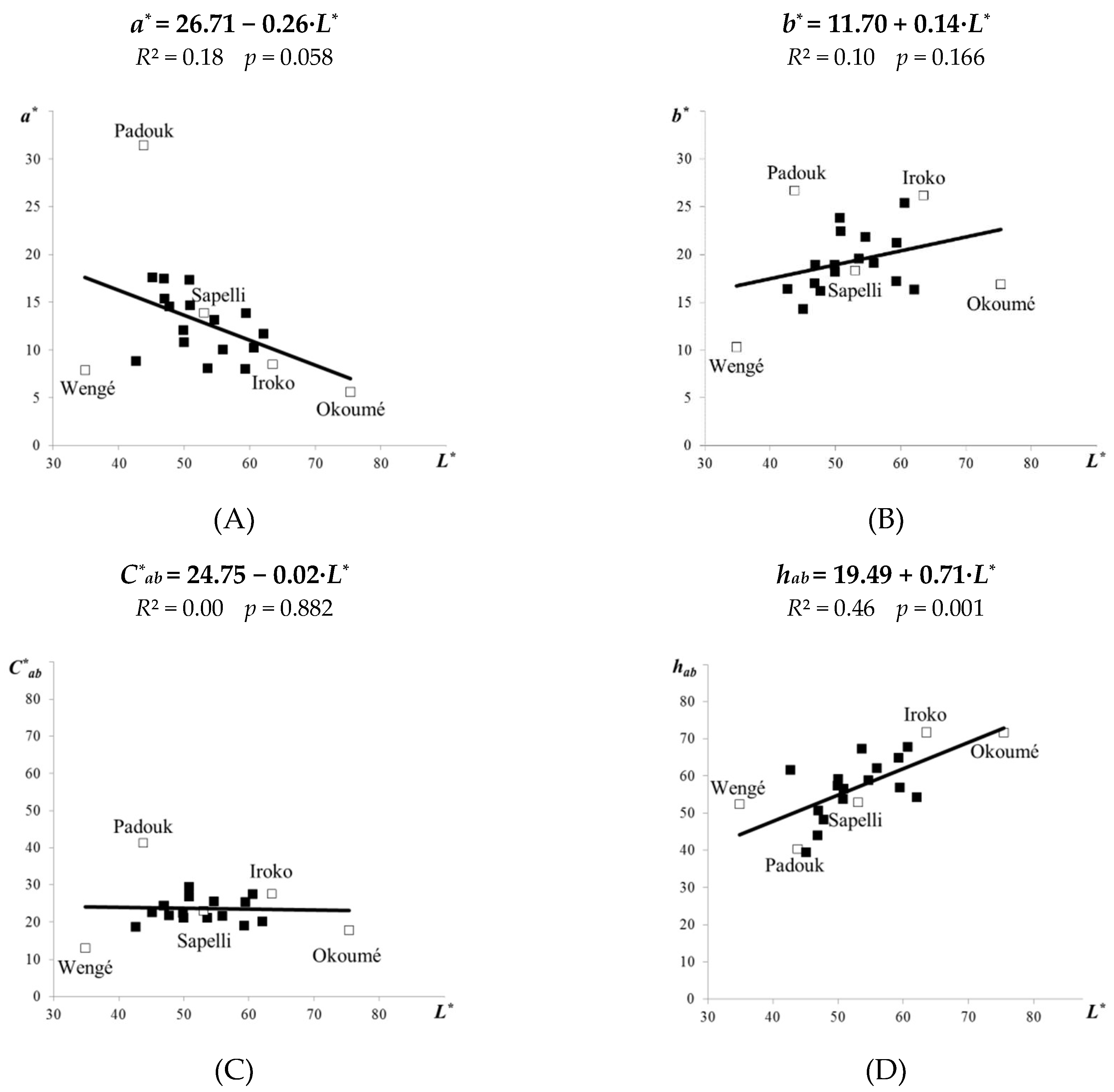
All 21 tropical woods had the colour parameters a* and b* in a positive sphere of distribution. The redness index (+a*) ranged from 5.61 (okoumé) to 31.42 (padouk), and the yellowness index (+b*) from 10.33 (wengé) to 26.72 (padouk). This result suggests that woods coloured a brighter red or yellow, such as padouk, may belong also to darker species. When the values of a* and b* were for 21 tropical wood species evaluated in comparison to their lightness L*, different tendencies of linear correlation were found. Value a* had a negative correlation against the L* (Figure 2A; R2 = 0.18), while the b* had a positive correlation against the L* (Figure 2B; R2 = 0.10). Nishino et al. [43] determined similar dependences between a* or b* and L* for 97 wood species from French Guiana.
The colour saturation C*ab indicates the distance from the chromatic point (a* = 0, b* = 0) on the CIE 1976 L*a*b* colour space. For tropical woods the values of colour saturation ranged from 13.02 (wengé—characterized by a little intensive red-yellow shade) to 41.26 (padouk—characterized by a strong intensive red-yellow shade). Graphical analysis of the C*ab against the L* for 21 tropical woods is present in Figure 2C. The C*ab exhibited only a minimal decrease at higher values of lightness L* with zero coefficient of determination (R2 = 0.00), i.e., was not found significant correlation.
The hue tone angle hab ranges for wood between 0° and 90° (the first quadrate), where 0° represents the red colour and 90° represents the yellow colour. The yellow shade prevailed to the red shade for a vast majority of tested tropical woods, typically for iroko and okoumé. On the contrary, for padouk, having the most red and yellow striking shades, a slightly more dominant was the red shade. A reasonable positive linear correlation between the hue tone angle hab and the lightness L* of individual tropical woods is presented in Figure 2D (R2 = 0.46).
This result suggests that tropical woods with a dominant yellow shade are usually lighter. Németh [44] also found a linear correlation between the lightness L* and the hue angle hab examining the colour co-ordinates of different temperate wood species.
3.2. Colour of Tropical Woods Exposed to the Fungus Coniophora puteana
Woods attacked by the brown-rot fungi gradually acquire deeper shades of brown of yellow [15,22,27,37,45]. However, specific colour changes during action of the brown-rot fungi can also occur, depending on the wood species [37,46], the history of wood ageing before fungal attack [15], the degree and uniformity of decay [47], or the specific enzymes, Fenton and other low-molecular degradation systems, and pigments produced by fungal mycelia [45].
Weight losses of 21 tropical woods attacked for 6 weeks by the brown-rot fungus C. puteana ranged from 0.08% to 6.48% (Table 3). The differences of the lightness ∆L* and other colourimetric parameters ∆a*, ∆b*, ∆C*ab, ∆hab and ΔE*ab, determined as a difference between the fungal-attacked and the original tropical wood, are documented in Table 3. Visualization, together with the pantone swatches, of the top surfaces of selected tropical woods before and after their exposure to C. puteana is shown in Figure 3.
After exposition to the brown-rot fungus C. puteana, the top surfaces of the darkest tropical woods wengé and ipé (Table 2) developed significantly lighter shades with the lightness increase ΔL* +10.46 and +2.47 (Table 3). This “unexpected” result can be explained by a washout or deterioration of dark extractives present in darker wood species during the mycological test performed in humid environment in Petri dishes. On the contrary, the most noticeable darkening was observed in the top surfaces of the lightest and medium light tropical woods sapelli, okoumé, iroko and ovengol with the ΔL* from −21.54 to −11.08 (Table 3). This “expected” result can be explained by degradation of white cellulose and hemicelluloses in presence of hydrolases and Fenton agent produced by C. puteana [45]. Previous studies have also reported a darkening of lighter European woods (beech and pine) due to decay processes [15,16].
A pronounced shade of the red due to C. puteana obtained only wengé, with Δa* +4.98. A lighter shade of red obtained okoumé, zebrano, iroko, and blue gum, with Δa* from +0.31 to +2.53 (Table 3). Conversely, the other tropical woods developed a greener shade, with Δa* from −0.22 to −7.76, the most markedly sapelli, padouk and macaranduba, with Δa* ≥ −5.68 (Table 3). The significant greening of padouk may be justified by its intensive red shade in the original state (Table 2).
The majority of tropical woods attacked by C. puteana obtained a more yellow shade, typically wengé, okoumé and blue gum, with Δb* from +4.42 to +10.05. However, three tropical woods—padouk, sapelli, and yellow balau—showed a significant tendency to become bluer, with Δb* from −3.08 to −8.74 (Table 3).
After fungal attack, a significant positive change in the colour saturation ΔC*ab had wengé +11.16, okoumé +5.93, and blue gum +5.15 (Table 3). It is an interesting knowledge, because the original wengé was the darkest species and okoumé the lightest one (Table 2). On the contrary, an evident negative change in ΔC*ab had sapelli, padouk, and yellow balau, with ΔC*ab from −4.04 to −10.84 (Table 3). However in a summary, the colour saturation of the tested original or fungal-attacked tropical woods had no significance to their lightness (Figure 2C, Figure 4C).
The positive differences in the hue tone angle Δhab indicate that the wood surfaces changed due to the brown-rot fungus more towards yellowish as to reddish. The largest positive change in Δhab was observed for maçaranduba, bubinga, dark red meranti, and sapelli in range from +13.46 to +7.21 (Table 3). A significantly negative Δhab, connected with more evident redness as yellowing, obtained padouk −5.49, and iroko −4.30. Statistically insignificant redness occurred for blue gum, ovengol, and zebrano, with Δhab from −0.99 to −0.09 (Table 3).
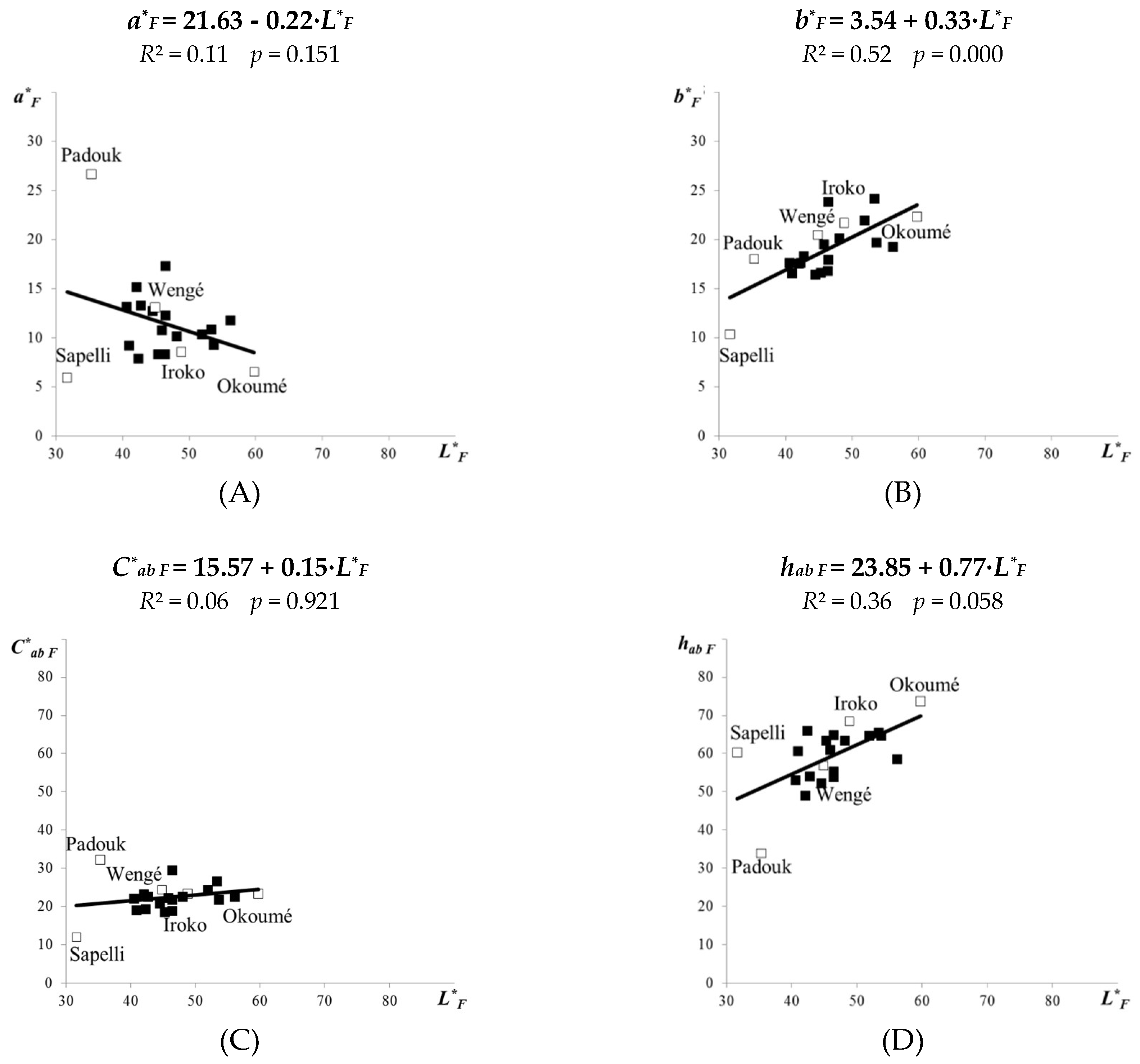
The linear correlations between the colourimetric parameters a*F, C*ab F or hab F versus the lightness L*F for the 21 fungal-attacked tropical woods (Figure 4A,C,D) remained almost the same as were determined for the 21 original tropical woods (Figure 2A,C,D).
In the fungal-attacked tropical woods, again no evident relationships were found between the lightness and redness (R2 = 0.11), or the lightness and colour saturation (R2 = 0.06). Indirectly, it can be stated that fungal attack did not have an apparent effect on these relationships. However, due to C. puteana the relationship between lightness and hue tone angle evidently decreased (R2 = 0.36 for hab F—Figure 4D; while previously R2 = 0.46 for hab—Figure 2D).
Conversely, the b*F coordinate, which indicates yellowing, grew more evidently with L*F for the 21 fungal-attacked tropical woods. It is evident from comparing the slope trend B and the coefficient of determination R2 in Figure 2B (b* = 11.70 + 0.14 · L*; B = 0.14; R2 = 0.10) with the same parameters in Figure 4B (b*F = 3.54 + 0.33 · L*F; B = 0.33; R2 = 0.52). As the slope trend B increased more evidently only for the yellow colour coordinate b*, from 0.14 to 0.33, there indirectly was confirmed that at brown rot some lighter tropical woods (such as okoumé, or dark red meranti) obtain a more yellow shade.
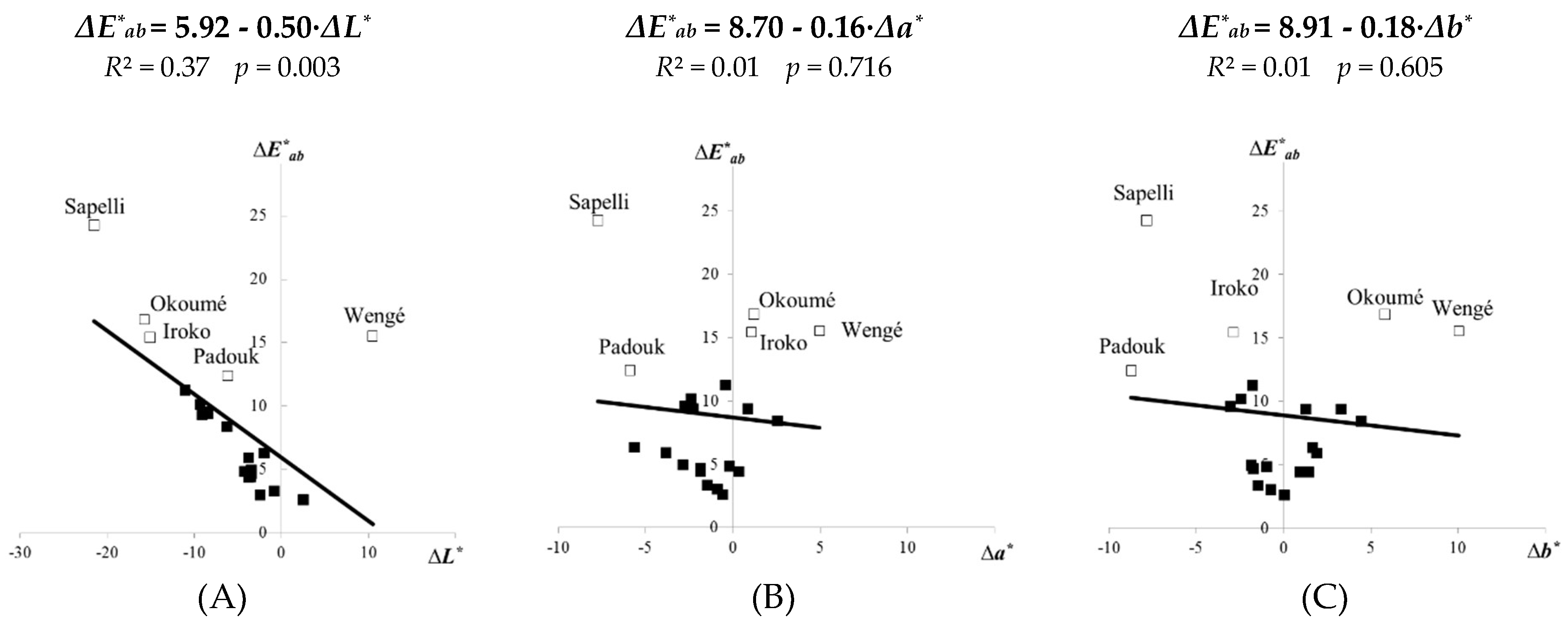
The total colour differences ΔE∗ab of the fungal-attacked tropical woods were significantly similar to their lightness changes ΔL∗ (Figure 5A), however, values of ΔE∗ab were greater as ΔL∗ in an accordance with Equation (3) (Table 3). Usually, the lightest and medium light tropical woods okoumé, sapelli, iroko and ovengol had the highest ΔE∗ab values from 11.28 to 24.26. This result is in an accordance with other works dealing with the durability and colour changes of tropical woods due to biological deterioration [48,49]. However, it was simultaneously observed that after attack by C. puteana the top surfaces of wengé and padouk (darker tropical woods—Table 2) also had high values of ΔE∗ab 15.80 and 12.41 (Table 3, Figure 5A). This result can be explained by washout of dark extractives presented in darker wood species during the mycological test, similarly mentioned for the lightness changes. On the contrary, effects of the Δa∗ and Δb∗ values of all tropical species on the ΔE∗ab value were not significant (Figure 5B,C).
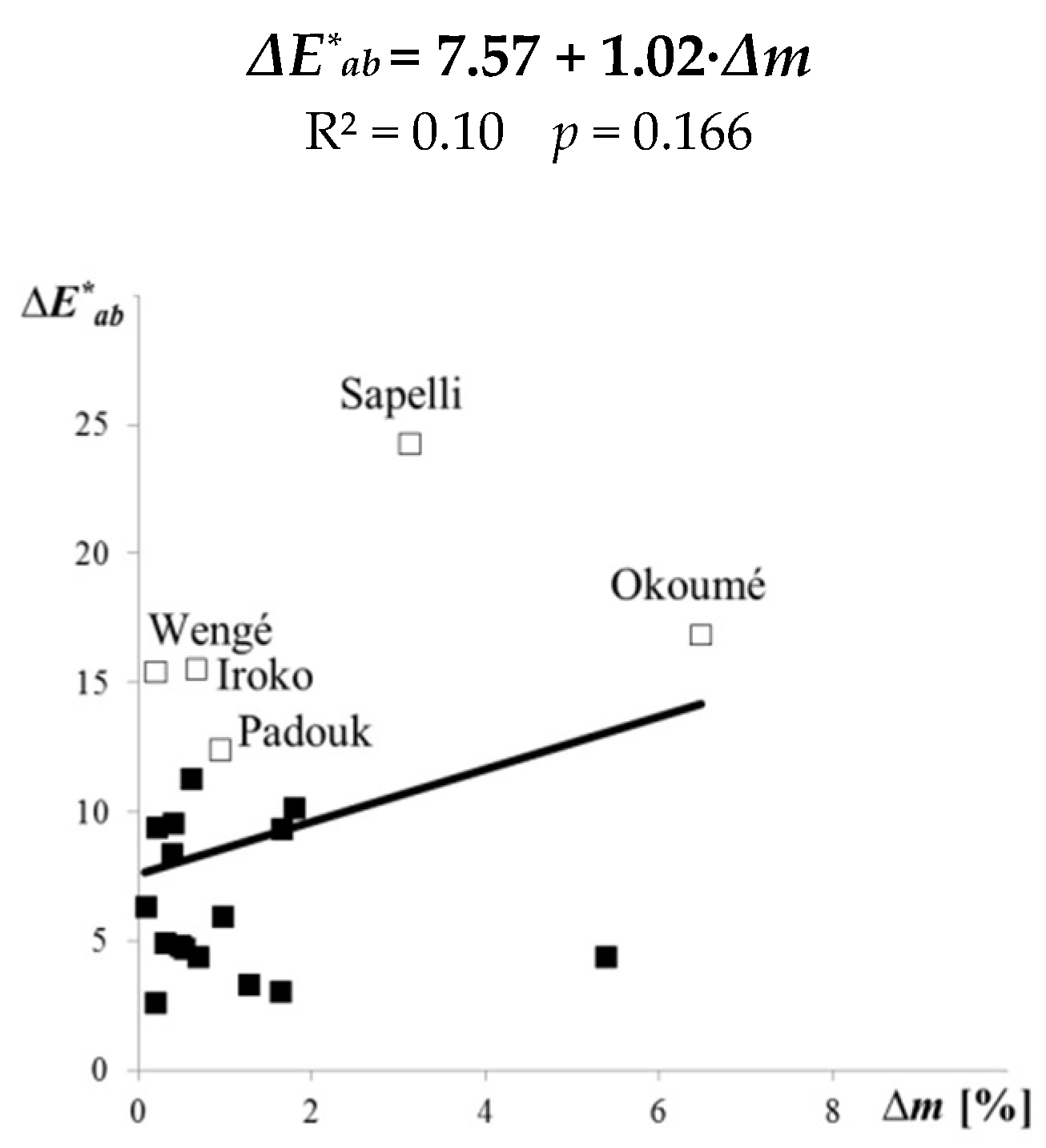
Several studies have shown that there are some relationships between the colour parameters of wood and its decay resistance [15,16,46,47,48,49,50]. The relationship between the colour and weight loss of decayed wood is based on the type and amount of wood extractives, which have effect on the colour, and on its decay resistance. Such relationships could be encouraging but not always sufficient for predicting decay resistance of tested woods. Specifically, from our experiment it is evident that for 21 tropical woods no dependency was found between the total colour difference ΔE∗ab and the decay resistance determined as weight loss Δm (Figure 6; R2 = 0.10).
Generally, the intensity and specificity of colour changes in woods attacked by brown rot—when browning can be connected with yellowing or bluing and also with reddening or greening—are influenced not only by the enzymatic and pigment specification of the individual brown-rot fungus, but also by the wood species (e.g., presence of specific extracts), the degree of its decay, and the environmental factors. Therefore, some of tested tropical woods at exposition to C. puteana had a more yellowish shade while other ones a more reddish shade, when all the colour changes depended probably on their molecular structure and natural durability. In future experiments, we would like to analyse these factors in more detail.
Colour changes of tropical woods exposed in interiors of buildings are unwanted. Brown-rot fungi decrease their strength and can worsen their colour and aesthetic. On the contrary, some benefits of rotting can be interesting for claddings and other decorative products, but in this situation the intentional decay has to be performed before they are installed. By processing spalted wood artists can create decorative material, such as fine art mosaics, furniture, and dishes, which have been used for centuries [51] and also hold an interesting niche modern market in decorative veneers or other decorative materials in North America and Europe [17].
4. Conclusions
Several tropical woods have positive surface characteristics, such as interesting texture and colour, and high natural durability, which is an essential requisite for wooden constructions exposed to the environment with a high risk of biodeterioration by fungi and insects. For architects, changes in their original colour at biodeterioration processes are important. The colour analyses of 21 tropical woods before and after intentional attack by the brown rot-fungus C. puteana led to the following conclusions:
- For the original tropical woods, the a* coordinate (redding) declined with increase of the lightness L*, while the b* (yellowing) and h*ab (hue ton angle) coordinates grew with the lightness. No significance was found between C*ab (colour saturation) and L*.
- For the fungal-attacked tropical woods, the linear correlations between the colour coordinates a*, C*ab or hab and the lightness L* remained almost the same as for the original tropical woods, and only the b* coordinate grew more evidently in relation to L*.
- The total discoloration ΔE∗ab values were highest for the top surfaces of the lighter species (sapelli, okoumé, iroko) and the darkest species (wengé), when the ΔE∗ab were justified by the marked change of the lightness ΔL*.
- Significant changes in the lightness and colouration of the fungal-attacked tropical woods indicated these colour changes could be caused not only by the biodegradation of polysaccharides, but also by biodegradation or leaching of some extractives during laboratory mycological tests.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “conceptualization, Z.V. and L.R.; methodology, Z.V. and L.R.; software, Z.V. and L.R.; validation, Z.V. and L.R.; formal analysis, Z.V. and L.R.; investigation, Z.V. and L.R.; resources, Z.V. and L.R.; data curation, Z.V. and L.R.; writing—original draft preparation, Z.V. and L.R.; writing—review and editing, Z.V. and L.R.; visualization, Z.V.; supervision, L.R..; project administration, L.R.
Acknowledgments
This work was supported by the Slovak Research and Development Agency under the contract No. APVV-17-0583, and the VEGA project 1/0729/18.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Janin, G.; Gonçalez, J.C.; Ananías, R.A.; Charrier, B.; Silva, G.F.D.; Dilem, A. Aesthetics appreciation of wood colour and patterns by colorimetry. Part 1. Colorimetry theory for the CIE Lab system. Maderas Cienc. Tecnol. 2001, 3, 14. [Google Scholar]
- Tolvaj, L.; Persze, L.; Lang, E. Correlation between hue angle and lightness of wood species grown in Hungary. Wood Res. 2013, 58, 141–145. [Google Scholar]
- Slabejová, G.; Šmidriaková, M.; Fekiač, J. Gloss of transparent coating on beech wood surface. Acta Fac. Xylologiae Zvolen 2016, 58, 37–44. [Google Scholar]
- Hon, D.N.-S.; Minemura, N. Color and discoloration. In Wood and Cellulosic Chemistry, 2nd ed.; Hon, D.N.-S., Shiraishi, N., Eds.; CRC Press: New York, USA, 2000; pp. 385–442. [Google Scholar]
- Babiak, M.; Kubovský, I.; Mamoňová, M. Color space of the selected domestic species. In Interaction of Wood with Various Forms of Energy, 1st ed.; Kurjatko, S., Kúdela, J., Eds.; Technical University in Zvolen: Zvolen, Slovakia, 2004; pp. 113–117. [Google Scholar]
- Katuščák, S.; Kucera, J. CIE orthogonal and cylindrical color parameters and the color sequences of the temperate wood species. Wood Res. 2000, 45, 9–21. [Google Scholar]
- da Silva, R.A.F.; Setter, C.; Mazette, S.S.; de Melo, R.R.; Stangerlin, D.M. Colorimetry of wood from thirty tropical species. Ciênc. Madeira 2017, 8, 36–41. [Google Scholar]
- Meints, T.; Teischinger, A.; Stingl, R.; Hansmann, C. Wood colour of central European wood species: CIE Lab characterisation and colour intensification. Eur. J. Wood Wood Prod. 2017, 75, 499–509. [Google Scholar] [CrossRef]
- Klement, I.; Vilkovská, T. Color characteristics of red false heartwood and mature wood of beech (Fagus sylvatica L.) determining by different colour saturation city coordinates. Sustainability 2019, 11, 690. [Google Scholar] [CrossRef]
- Mosedale, J.R.; Charrier, B.; Janin, G. Genetic control of wood colour, density and heartwood ellagitannin content of European oak (Quercus petraea and Quercus robur). Forestry 1996, 69, 111–124. [Google Scholar] [CrossRef]
- Phelps, J.E.; McGinnes, E.A.; Garret, H.E.; Cox, G.S. Growth quality evaluation of black walnut wood. II. Color analyses of veneer produced on different sites. Wood Fiber Sci. 1982, 15, 177–185. [Google Scholar]
- Derkyi, N.S.A.; Bailleres, H.; Chaix, G.; Thevenon, M.F. Colour variation in teak (Tectona grandis) wood from plantations across the ecological zones of Ghana. Ghana J. For. 2009, 25, 40–48. [Google Scholar] [CrossRef]
- Kržišnik, D.; Lesar, B.; Thaler, N.; Humar, M. Influence of natural and artificial weathering on the colour change of different wood and wood-based materials. Forests 2018, 9, 488. [Google Scholar] [CrossRef]
- Reinprecht, L.; Mamoňová, M.; Pánek, M.; Kačík, F. The impact of natural and artificial weathering on the visual, colour and structural changes of seven tropical woods. Eur. J. Wood Wood Prod. 2018, 76, 175–190. [Google Scholar] [CrossRef]
- Reinprecht, L.; Hulla, M. Colour changes in beech wood modified with essential oils due to fungal and ageing-fungal attacks with Coniophora puteana. Drewno 2015, 58, 37–48. [Google Scholar]
- Vidholdová, Z.; Slabejová, G.; Polomský, J. Colour changes of Scots pine wood due to action of the white-rot fungus Trametes versicolor. In Protecting Trees and Wood, 1st ed.; Hlaváč, P., Vidholdová, Z., Eds.; Technical University in Zvolen: Zvolen, Slovakia, 2016; pp. 61–66. [Google Scholar]
- Van Court, R.C.; Robinson, S.C. Stimulating Production of Pigment-Type Secondary Metabolites from Soft Rotting Wood Decay Fungi (“Spalting” Fungi). In Advances in Biochemical Engineering/Biotechnology, 1st ed.; Springer Nature: Basel, Switzerland, 2019; p. 16. [Google Scholar]
- Rayner, A.D.; Boddy, L. Fungal Decomposition of Wood. Its Biology and Ecology; Antony Rowe Ltd.: Chippenham, UK, 1997; p. 587. [Google Scholar]
- Pandey, K.K.; Pitman, A.J. FTIR studies of the changes in wood chemistry following decay by brown-rot and white-rot fungi. Int. Biodeter. Biodegr. 2003, 52, 151–160. [Google Scholar] [CrossRef]
- Zabel, R.A.; Morrell, J.J. Wood Microbiology: Decay and Its Prevention; Academic Press: London, UK, 2012; p. 476. [Google Scholar]
- Nascimento, M.S.; Santana, A.L.B.D.; Maranhão, C.A.; Oliveira, L.S.; Bieber, L. Phenolic extractives and natural resistance of wood. In Biodegradation-Life of Science; Chamy, R., Rosenkranz, F., Eds.; InTech: Rijeka, Croatia, 2013; pp. 349–370. [Google Scholar]
- Sablík, P.; Giagli, K.; Pařil, P.; Baar, J.; Rademacher, P. Impact of extractive chemical compounds from durable wood species on fungal decay after impregnation of nondurable wood species. Eur. J. Wood Wood Prod. 2016, 74, 231–236. [Google Scholar] [CrossRef]
- Schmidt, O. Indoor wood-decay basidiomycetes: Damage, causal fungi, physiology, identification and characterization, prevention and control. Mycol. Prog. 2007, 6, 261–279. [Google Scholar] [CrossRef]
- Frankl, J. Wood-damaging fungi in truss structures of baroque churches. J. Perform. Constr. Facil. 2015, 29, 04014138. [Google Scholar] [CrossRef]
- Hyde, K.D.; Al-Hatmi, A.M.S.; Andersen, B.; Boekhout, T.; Buzina, W.; Dawson, T.L.; Eastwood, D.C.; Jones, E.B.G.; Hoog, S.; Kang, Y.; et al. The world’s ten most feared fungi. Fungal Divers. 2018, 93, 161–194. [Google Scholar] [CrossRef]
- Gabriel, J.; Švec, K. Occurrence of indoor wood decay basidiomycetes in Europe. Fungal Biol. Rev. 2017, 31, 212–217. [Google Scholar] [CrossRef]
- Irbe, I.; Andersone, I.; Andersons, B.; Noldt, G.; Dizhbite, T.; Kurnosova, N.; Nuopponen, M.; Stewart, D. Characterisation of the initial degradation stage of Scots pine (Pinus sylvestris L.) sapwood after attack by brow-rot fungus Coniophora puteana. Biodegradation 2011, 22, 719–728. [Google Scholar] [CrossRef]
- Robinson, S.C.; Laks, P.E. Wood species and culture age affect zone line production of Xylaria polymorpha. Open Mycol. J. 2010, 4, 18–21. [Google Scholar] [CrossRef]
- Robinson, S.C.; Tudor, D.; Cooper, P.A. Wood preference of spalting fungi in urban hardwood species. Int. Biodeter. Biodegr. 2011, 65, 1145–1149. [Google Scholar] [CrossRef]
- Robinson, S.C.; Tudor, D.; Cooper, P.A. Feasibility of using red pigment producing fungi to stain wood for decorative applications. Can. J. For. Res. 2011, 41, 1722–1728. [Google Scholar] [CrossRef]
- Robinson, S.C. Developing fungal pigments for “painting” vascular plants. Appl. Microbiol. Biotechnol. 2012, 93, 1389–1394. [Google Scholar] [CrossRef]
- Robinson, S.C.; Tudor, D.; Cooper, P.A. Utilizing pigment-producing fungi to add commercial value to American beech (Fagus grandifolia). Appl. Microbiol. Biotechnol. 2012, 93, 1041–1048. [Google Scholar] [CrossRef]
- Beck, H.G.; Freitas, S.; Weber, G.; Robinson, S.C.; Morrell, J.J. Resistance of fungal derived pigments to ultraviolet light exposure. In International Research Group in Wood Protection; IRG/WP: St. George, UT, USA, 2014. [Google Scholar]
- Vega Gutierrez, S.; Robinson, S.C. Microscopic analysis of pigments extracted from spalting fungi. J. Fungi 2017, 3, 15. [Google Scholar] [CrossRef]
- Blanchette, R.A. Screening wood decayed by white rot fungi for preferential lignin degradation. Appl. Environ. Microbiol. 1984, 48, 647–653. [Google Scholar]
- Stalpers, J.A.; Vlug, I. Confistulina, the anamorphs of Fistulina hepatica. Can. J. Bot. 1983, 61, 1660–1666. [Google Scholar] [CrossRef]
- Hillis, W.E. Heartwood and Tree Exudates; Springer: Berlin/Heidelberg, Germany, 1987; p. 267. [Google Scholar]
- Coulson, J. Wood in Construction—How to Avoid Costly Mistakes; John Wiley & Sons Ltd.: Chichester, UK, 2012; p. 208. [Google Scholar]
- EN 350. Durability of Wood and Wood-Based Products. Testing and Classification of the Durability to Biological Agents of Wood and Wood-Based Materials; European Committee for Standardization: Brussels, Belgium, 2016. [Google Scholar]
- Wagenführ, R. Holzatlas; Fachbuchverlag Leipzig, Carl Hanser Verlag: Munchen, Germany, 2007; p. 816. [Google Scholar]
- URL 1. The IUCN Red List of Threatened Species. Version 2017-2. Available online: www.iucnredlist.org (accessed on 16 November 2017).
- CIE. Colorimetry—Part 4: CIE 1976 L*a*b Colour Space; CIE DS 014-4.3/E:2007; CIE Central Bureau: Vienna, Austria, 2007. [Google Scholar]
- Nishino, Y.; Janin, G.; Chanson, B.; Détienne, P.; Gril, J.; Thibaut, B. Colorimetry of wood specimens from French Guiana. J. Wood Sci. 1998, 44, 3–8. [Google Scholar] [CrossRef]
- Németh, K. The colour of wood in CIE Lab system. Az Erdészeti és Faipari Egyetem Tudományos Közleményei 1982, 2, 125–135. [Google Scholar]
- Eriksson, K.-E.L.; Blanchett, R.A.; Ander, P. Microbial and Enzymatic Degradation of Wood and Wood Components; Springer Series in Wood Science: Berlin/Heidelberg, Germany; New York, NY, USA, 1990; p. 407. [Google Scholar]
- Gierlinger, N.; Jacques, D.; Grabner, M.; Wimmer, R.; Schwanninger, M.; Rozenberg, P.; Pâques, L.E. Colour of larch heartwood and relationships to extractives and brown-rot decay resistance. Trees 2004, 18, 102–108. [Google Scholar] [CrossRef]
- Kokutse, A.D.; Stokes, A.; Baillères, H.; Kokou, K.; Baudasse, C. Decay resistance of Togolese teak (Tectona grandis Lf) heartwood and relationship with colour. Trees 2006, 20, 219–223. [Google Scholar] [CrossRef]
- Costa, M.D.A.; Costa, A.F.D.; Pastore, T.C.M.; Braga, J.W.B.; Gonçalez, J.C. Characterization of wood decay by rot fungi using colorimetry and infrared spectroscopy. Ciência Florestal 2011, 21, 567–577. [Google Scholar]
- Amusant, N.; Fournier, M.; Beauchene, J. Colour and decay resistance and its relationships in Eperua grandiflora. J. Ann. For. Sci. 2008, 65, 1–6. [Google Scholar] [CrossRef]
- Stangerlin, D.M.; Costa, A.F.D.; Gonçalez, J.C.; Pastore, T.C.M.; Garlet, A. Monitoring of biodeterioration of three Amazonian wood species by the colorimetry technique. Acta Amazon. 2013, 43, 429–438. [Google Scholar] [CrossRef]
- Blanchette, R.A.; Wilmering, A.M.; Baumeister, M. The use of green-stained wood caused by the fungus Chlorociboria in intarsia masterpieces from the 15th century. Holzforschung 1992, 46, 225–232. [Google Scholar] [CrossRef]
Figure 1.
The same four places on the top surface of specimen in which colour measurements were performed in its original and fungal-attacked state.
Figure 1.
The same four places on the top surface of specimen in which colour measurements were performed in its original and fungal-attacked state.

Figure 2.
Linear correlations between the lightness L* and the colour coordinates a*, b* (A, B), the colour saturation C*ab (C), and the hue tone angle hab (D)—for 21 original tropical wood species.
Figure 2.
Linear correlations between the lightness L* and the colour coordinates a*, b* (A, B), the colour saturation C*ab (C), and the hue tone angle hab (D)—for 21 original tropical wood species.

Figure 3.
Visualization and pantone swatches of the top surfaces of selected tropical woods before and after exposure to C. puteana.
Figure 3.
Visualization and pantone swatches of the top surfaces of selected tropical woods before and after exposure to C. puteana.

Figure 4.
Linear correlations between the lightness L* and the colour coordinates a*, b* (A, B), the colour saturation C*ab (C), and the hue tone angle hab (D)—for 21 tropical wood species attacked by the brown-rot fungus C. puteana.
Figure 4.
Linear correlations between the lightness L* and the colour coordinates a*, b* (A, B), the colour saturation C*ab (C), and the hue tone angle hab (D)—for 21 tropical wood species attacked by the brown-rot fungus C. puteana.

Figure 5.
Linear correlations between the total colour difference ΔE∗ab and changes of other colour parameter ΔL* (A), Δa* (B), Δb* (C)—for 21 tropical woods attacked with the brown-rot fungus C. puteana.
Figure 5.
Linear correlations between the total colour difference ΔE∗ab and changes of other colour parameter ΔL* (A), Δa* (B), Δb* (C)—for 21 tropical woods attacked with the brown-rot fungus C. puteana.

Figure 6.
Linear correlations between the total colour differences ΔE∗ab and the weight losses Δm of 21 tropical woods exposed 6 weeks to C. puteana.
Figure 6.
Linear correlations between the total colour differences ΔE∗ab and the weight losses Δm of 21 tropical woods exposed 6 weeks to C. puteana.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Tropical wood species used in the experiment.
| Family | Species | Species | Density at MC 12% (kg·m−3) | |
|---|---|---|---|---|
| Common Name 1) | Scientific Name | “Literature” [40] | “Experiment” | |
| Bignoniaceae | Ipé | Handroanthus serratifolius (Vahl) S.O.Grose 3) 4) | 960–1100 | 968 (26) |
| Burseraceae | Okoumé | Aucoumea klaineana Pierre 2) | 370–560 | 566 (27) |
| Cunoniaceae | Tineo | Weinmannia trichosperma Cav. | 570–650 | 646 (31) |
| Dipterocarpaceae | Dark red meranti | Shorea curtisii Dyer ex King 2) | 590–890 | 592 (10) |
| Yellow balau 5) | Shorea laevis Ridl. 2) | 900–1100 | 925 (55) | |
| Ebenaceae | Macassar ebony | Diospyros celebica Bakh. 2) | 1100–1200 | 1 013 (82) |
| Fabaceae | Doussié | Afzelia bipindensis Harms 2) | 750–950 | 889 (18) |
| Cerejeira | Amburana cearensis A. C. Sm. 2) | 550–650 | 651 (10) | |
| Bubinga | Guibourtia demeusei J. Léonard | 830–950 | 830 (13) | |
| Ovengol | Guibourtia ehie J. Léonard 2) | 700–910 | 755 (27) | |
| Merbau | Intsia bijuga O. Ktze. 2) | 830–900 | 837 (50) | |
| Santos rosewood | Machaerium scleroxylon Tul. 2) | 900–1000 | 904 (6) | |
| Zebrano | Microberlinia brazzavillensis Chev. 2) | 700–850 | 718 (23) | |
| Wengé | Millettia laurentii De Wild. 2) | 810–950 | 823 (37) | |
| Padouk | Pterocarpus soyauxii Taub. | 650–850 | 647 (37) | |
| Meliaceae | Sapelli | Entandrophragma cylindricum Sprague 2) | 510–750 | 631 (38) |
| Moraceae | Iroko | Milicia excelsa C. C. Berg 2) | 550–850 | 551 (16) |
| Myrtaceae | Karri | Eucalyptus diversicolor F. Muell. | 800–870 | 804 (30) |
| Blue gum | Eucalyptus globulus Labill. | 720–770 | 760 (59) | |
| Sapotaceae | Maçaranduba 6) | Manilkara bidenta A. Chev. | 900–1000 | 916 (19) |
| Makoré | Tieghemella heckelii Pierre 2) | 530–720 | 570 (25) | |
Notes: 1) by Association Technique Internationale des Bois Tropicaux (ATIBT) in France; 2) registered in The IUCN Red List of Threatened Species™ [41]; 3) name by EN 350 [39]; 4) previous name by [40] was Tabebuia serratifolia; 5) its other name is Bangkirai; 6) its other name is Massaranduba; Mean values of experimental densities are from 16 specimens (4 of those were used in this experiment) and standard deviations are in italic and parentheses.
Table 2.
Visualisations and colorimetric parameters of 21 tropical woods.
| Wood Species | Visual | Lightness L* | Redness +a* | Yellowness +b* | Colour Saturation C*ab | Hue Tone Angle hab |
|---|---|---|---|---|---|---|
| Ipé |  | 42.59 (0.79) | 8.87 (0.55) | 16.46 (1.01) | 18.70 (2.59) | 61.66 (1.22) |
| Okoumé |  | 75.36 (1.13) | 5.61 (0.61) | 16.89 (0.57) | 17.81 (0.65) | 71.66 (1.69) |
| Tineo |  | 47.69 (4.98) | 14.56 (1.81) | 16.23 (0.99) | 21.83 (1.77) | 48.26 (2.83) |
| Darkred meranti |  | 62.03 (1.52) | 11.76 (0.81) | 16.35 (0.64) | 20.15 (0.87) | 54.30 (1.56) |
| Yellow balau |  | 54.59 (0.92) | 13.22 (0.83) | 21.92 (0.88) | 25.60 (1.12) | 58.93 (1.02) |
| Macassar ebony |  | 59.40 (0.80) | 13.86 (0.68) | 21.29 (0.81) | 25.41 (0.91) | 56.93 (1.23) |
| Doussié |  | 60.57 (6.61) | 10.28 (1.89) | 25.42 (2.09) | 27.50 (1.81) | 67.93 (4.80) |
| Cerejeira |  | 49.93 (3.34) | 10.85 (1.52) | 18.19 (1.34) | 21.22 (1.59) | 59.24 (3.46) |
| Bubinga |  | 46.81 (1.23) | 17.50 (0.89) | 16.98 (1.17) | 24.40 (1.20) | 44.11 (2.00) |
| Ovengol |  | 53.56 (4.65) | 8.08 (1.07) | 19.63 (4.38) | 21.27 (4.33) | 67.36 (2.69) |
| Merbau |  | 46.92 (1.57) | 15.43 (0.53) | 18.97 (1.12) | 24.46 (1.09) | 50.83 (1.37) |
| Santos rosewood |  | 50.79 (3.39) | 14.68 (1.31) | 22.49 (3.13) | 26.88 (3.23) | 56.63 (2.44) |
| Zebrano |  | 55.85 (4.91) | 10.09 (1.06) | 19.18 (1.49) | 21.68 (1.63) | 62.24 (2.18) |
| Wengé |  | 34.88 (1.68) | 7.89 (0.64) | 10.33 (1.30) | 13.02 (1.24) | 52.42 (3.38) |
| Padouk |  | 43.78 (2.05) | 31.42 (1.61) | 26.72 (2.02) | 41.26 (2.32) | 40.34 (1.57) |
| Sapelli |  | 53.04 (1.69) | 13.88 (0.88) | 18.34 (0.72) | 23.01 (0.94) | 52.90 (1.62) |
| Iroko |  | 63.52 (6.78) | 8.49 (0.88) | 26.20 (5.69) | 27.57 (5.59) | 71.70 (2.54) |
| Karri |  | 50.72 (2.07) | 17.36 (1.13) | 23.84 (1.59) | 29.50 (1.83) | 53.92 (1.32) |
| Blue gum |  | 59.25 (3.37) | 8.05 (0.74) | 17.24 (1.23) | 19.03 (1.27) | 64.95 (1.98) |
| Maçaranduba |  | 45.06 (2.16) | 17.61 (2.39) | 14.34 (0.99) | 22.75 (2.23) | 39.45 (3.79) |
| Makoré |  | 49.85 (2.24) | 12.08 (0.88) | 18.95 (1.10) | 22.49 (1.15) | 57.47 (2.10) |
Notes: Mean values are from 16 measurements. Standard deviations are in italic and parentheses.
Table 3.
Weight loss and relative change of colourimetric parameters of 21 tropical woods caused by C. puteana.
Table 3.
Weight loss and relative change of colourimetric parameters of 21 tropical woods caused by C. puteana.
| Wood species | Δm | ΔL* | Δa* | Δb* | ΔC*ab | Δhab | ΔE*ab | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ipé | 0.20 (0.03) | 2.47 (1.32) | a | −0.59 (0.50) | c | 0.01 (0.31) | d | −0.26 (0.29) | d | 1.68 (1.56) | b | 2.63 (1.26) |
| Okoumé | 6.48 (2.98) | −15.74 (1.55) | a | 1.20 (0.23) | b | 5.80 (0.50) | a | 5.93 (0.49) | a | 1.70 (0.51) | d | 16.84 (1.34) |
| Tineo | 5.38 (3.71) | −3.59 (1.63) | d | −1.88 (1.12) | b | 1.41 (0.43) | c | −0.11 (0.92) | d | 6.03 (2.10) | a | 4.44 (1.68) |
| Dark red meranti | 0.22 (0.09) | −8.44 (0.96) | a | −2.29 (0.27) | a | 3.28 (1.04) | a | 1.71 (0.94) | a | 9.90 (1.70) | a | 9.43 (0.70) |
| Yellow balau | 0.40 (0.08) | −8.61 (1.06) | a | −2.77 (1.00) | a | −3.08 (0.95) | a | −4.04 (1.17) | a | 2.04 (1.77) | b | 9.59 (1.47) |
| Macassar ebony | 0.52 (0.30) | −3.48 (1.87) | a | −1.88 (0.31) | a | −1.76 (0.84) | a | −2.49 (0.86) | a | 1.57 (0.49) | a | 4.73 (0.81) |
| Doussié | 1.63 (1.51) | −2.51 (1.68) | d | −0.91 (0.76) | d | −0.76 (0.66) | d | −1.23 (0.84) | d | 1.36 (1.07) | d | 3.05 (1.60) |
| Cerejeira | 0.30 (0.15) | −3.51 (2.26) | b | −2.89 (1.57) | b | −1.86 (1.28) | b | −2.89 (1.67) | b | 5.73 (2.12) | c | 4.98 (2.93) |
| Bubinga | 0.96 (0.13) | −3.81 (1.24) | a | −3.87 (0.85) | a | 1.89 (0.47) | a | −1.11 (0.90) | c | 10.29 (1.10) | a | 5.96 (0.15) |
| Ovengol | 0.61 (1.02) | −11.08 (1.28) | a | −0.46 (0.48) | d | −1.81 (0.84) | a | −1.83 (0.87) | b | −0.84 (1.10) | d | 11.28 (1.24) |
| Merbau | 0.48 (0.32) | −4.29 (1.71) | a | −0.22 (1.71) | d | −1.01 (1.25) | d | −0.88 (1.95) | d | −1.16 (1.99) | d | 4.87 (1.78) |
| Santos rosewood | 1.28 (0.31) | −0.83 (0.66) | d | −1.49 (1.67) | c | −1.48 (2.55) | d | −2.49 (0.86) | d | 1.57 (0.49) | d | 3.36 (1.89) |
| Zebrano | 1.65 (0.62) | −9.14 (1.01) | a | 0.82 (1.04) | d | 1.26 (1.18) | d | 1.52 (1.46) | d | −0.09 (1.69) | d | 9.39 (1.08) |
| Wengé | 0.67 (0.35) | 10.46 (2.17) | a | 4.98 (0.72) | a | 10.05 (1.87) | a | 11.16 (1.50) | a | 4.69 (2.60) | b | 15.54 (1.55) |
| Padouk | 0.95 (1.69) | −6.18 (1.32) | a | −5.91 (0.96) | a | −8.74 (1.93) | a | −9.93 (1.46) | a | −5.49 (2.70) | a | 12.41 (1.41) |
| Sapelli | 3.12 (1.15) | −21.54 (0.92) | a | −7.76 (1.26) | a | −7.87 (1.52) | a | −10.84 (1.99) | a | 7.21 (1.11) | a | 24.26 (1.52) |
| Iroko | 0.20 (0.97) | −15.08 (3.76) | a | 1.07 (0.57) | c | −2.89 (1.41) | c | −2.37 (1.49) | d | −4.30 (0.71) | a | 15.44 (3.89) |
| Karri | 0.69 (0.47) | −3.83 (1.37) | a | 0.31 (1.23) | d | 0.89 (1.41) | d | 0.91 (1.81) | d | 0.54 (0.89) | d | 4.43 (1.17) |
| Blue gum | 0.38 (0.21) | −6.27 (2.54) | a | 2.53 (1.46) | a | 4.42 (0.88) | a | 5.15 (1.52) | a | −0.99 (1.35) | d | 8.42 (1.91) |
| Maçaranduba | 0.08 (0.05) | −1.99 (1.21) | b | −5.68 (0.40) | a | 1.66 (0.40) | b | −2.81 (0.38) | a | 13.46 (1.08) | a | 6.35 (0.64) |
| Makoré | 1.78 (1.84) | −9.43 (1.69) | a | −2.44 (1.37) | a | −2.45 (1.65) | b | −3.33 (2.09) | b | 2.35 (1.28) | c | 10.19 (2.10) |
Note: Mean values of colourimetric parameters are from 16 measurements, and of weight losses from 4 specimens. Standard deviations are in italics and parentheses. The t-test analysed the colour changes in relation to the original wood at the 99.9% significance level (a), the 99% significance level (b), the 95% significance level (c), and without an evident significant difference at p ≥ 0.05 (d).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vidholdová, Z.; Reinprecht, L. The Colour of Tropical Woods Influenced by Brown Rot. Forests 2019, 10, 322. https://0-doi-org.brum.beds.ac.uk/10.3390/f10040322
AMA Style
Vidholdová Z, Reinprecht L. The Colour of Tropical Woods Influenced by Brown Rot. Forests. 2019; 10(4):322. https://0-doi-org.brum.beds.ac.uk/10.3390/f10040322
Chicago/Turabian StyleVidholdová, Zuzana, and Ladislav Reinprecht. 2019. "The Colour of Tropical Woods Influenced by Brown Rot" Forests 10, no. 4: 322. https://0-doi-org.brum.beds.ac.uk/10.3390/f10040322
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.