The Spring Assessing Method of the Threat of Melolontha spp. grubs for Scots Pine Plantations
by
 , and
, and
Hanna Szmidla
1,* ,
,
Monika Małecka
1,
Miłosz Tkaczyk
1,
Grzegorz Tarwacki
1 and
Zbigniew Sierota
2 1
Department of Forest Protection, Forest Research Institute, Braci Leśnej 3, Sękocin Stary, 05-090 Raszyn, Poland
2
Department of Forestry and Forest Ecology, Faculty of Environmental Management and Agriculture, Warmia and Mazury University in Olsztyn, Pl. Łódzki 2, 10-727 Olsztyn, Poland
*
Author to whom correspondence should be addressed.
Forests 2019, 10(5), 399; https://0-doi-org.brum.beds.ac.uk/10.3390/f10050399
Submission received: 18 March 2019
/
Revised: 5 May 2019
/
Accepted: 7 May 2019
/
Published: 9 May 2019
(This article belongs to the Special Issue Impacts, Monitoring and Management of Forest Pests and Diseases)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Root-feeding Melolonthinae larvae are a forest pest species in Europe that can exert serious damage. In Poland, they are classified as the most dangerous pest on land dedicated to afforestation and the most serious threat to natural regeneration in the stands. This study was performed in three forest districts in east Poland (Lubartów, Marcule, and Wyszków forest districts) in mixed conifer forests, where the presence of Melolontha spp. grubs was evaluated in autumn and spring of 2012 to 2017, respectively. In spring 2012, 2014, and 2016, consecutively ‘small sawdust pits’ in rows between seedlings were prepared, and in adjacent inter-rows similar control pits without sawdust were marked. In spring and autumn of the following year, sawdust and soil from both types of pits were sieved and Melolonthinae larvae were counted and compared. More grubs were found in sawdust pits in spring than in autumn. In Marcule Forest District (FD) (2014), more grubs were found in inter-rows than in rows with seedlings, when compared to grubs detected using the traditional method of “autumn large pits assessment”, recommended by Polish forest rules. The Melolonthinae population size and location of grubs were related to the weather conditions in the evaluated periods, as well. We conclude that to properly assess the cockchafer threat, it is necessary to perform spring assessment and to search in inter-rows. This could be recommended for decisions on control.
1. Introduction
Root-feeding insects are key components of many terrestrial ecosystems. The detrimental effect of below ground insect herbivory in agricultural and forest ecosystems is widely appreciated, because root-damaging pests cause great economic losses [1,2]. Among the many insects that have substantial impacts on forest management in European countries, the genus of Melolontha spp. (Coleoptera: Scarabaeidae) are especially important. The cockchafer genus, Melolontha spp., is estimated to have been a pest on 200,000 hectares, mainly in Central Europe, over the past 20 years [3,4,5].
In Poland, the most important species are Melolontha melolontha L. (common or May cockchafer), and M. hippocastani Fabr. (forest cockchafer) [6,7,8]. These species occur at varied frequency throughout the country, but are commonly found in the central and southeastern parts of Poland. The area of occurrence of the two species considered as pests has increased in recent years, from 10.4 thousand hectares in 2016 to 18.3 thousand hectares in 2017 [9]. In fact, in certain regions in Poland, the high population density of Melolontha spp. grubs makes reforestation or afforestation impossible.
Melolontha spp. have a 3- to 5-year life cycle: In Central Europe, the life cycle is typically four years [10,11]. Melolontha melolontha L and Melolontha hippocastani Fabr. have very similar biology [12]. During the life cycle, the insect undergoes a complete transformation characterized by varying morphology and behavior at each life stage. The grubs (larvae) are more problematic in forests because they damage the roots of seedlings and trees. [10,13,14]. Dead roots of trees and plants are colonized and decomposed by fungi and bacteria, and released carbon dioxide serves to attract grubs to migrate to the root system and to feed on living roots [15,16]. The most intensive feeding periods for grubs take place after molting (autumn) and after vertical migration to the surface in the spring; it is during these periods that the greatest damage to tree roots occurs. Adult beetles feed in tree crowns, causing defoliation, reduced photosynthetic capacity, and weakness in the trees [17].
Until 2010, pesticides were used to effectively control both adults and larvae [14,18]. Two legal acts adopted in 2009 have had considerable influence on the extent and form of allowable pesticide use: (a) Directive 2009/128/EC of the European Parliament and of the Council on 21 October 2009 established a framework for community action to achieve sustainable use of pesticides; and (b) Regulation (EC) no. 1107/2009 of the European Parliament and of the Council on 21 October 2009 regulated the placement of plant protection products on the market and repealed Council Directives 79/117/EEC and 91/414/EEC. These acts limited chemical control of the cockchafer population, shifting management attention to natural (birds after soil ploughing, use of antifeedants, e.g., buckwheat) or biological (entomopathogenic fungi, parasitic nematodes) methods of control. The permitted insecticides are used as a last resort. In order to practically apply one of these non-chemical methods, it is necessary to know the current threat of the area designated for afforestation or renewal, risk assessment, and decision making.
Globally, field methods used in the monitoring of root pests are highly diverse. To monitor the threat represented by the cockchafer and its larval stage in Australia, 5 to 10 soil cores of a 10 cm depth were taken before pupation [19]. In Germany, a 4 × 1 m2 Goettinger frame was used to search for cockchafer larvae in soil [20]. In Austria, Pernfuss et al. [21] dug three square holes per plot (20 × 20 cm wide and 20 cm deep) and sampled garden chafer (Phyllopertha horticola) larvae from these locations before and after control treatment. In Poland, the threat from Melolontha spp. larvae to young plantations is assessed in autumn, one year before tree planting. According to the Polish Instruction of Forest Protection [22], this threat assessment is performed between 15 September and 30 October by counting the number of grubs in the soil in at least six large pits (1 × 0.5 × 0.5 m) placed in each hectare of afforested or restored area. The age of the grubs is determined by the width of the head capsule—one-year up to 2.5 mm, two-year up to 4.0 mm, and three-year and older above 4.0 mm [22]. The number of grubs in one pit exceeding five one-year grubs, four two-year grubs, or three three-year and older grubs specifies the need for protective treatment.
An alternative method has been described where the Melolontha spp. threat is assessed in spring in small soil pits (0.2 × 0.2 × 0.3 m) that are easier to excavate and have been filled with fresh pine sawdust [23]. Sawdust is a nutritional base for colonization by local populations of bacteria and fungi involved in cellulose and lignin degradation [24,25,26,27]. This degradation produces: (1) Many antibiotics, inhibitors, and biopesticides, therefore shaping some microbial properties of soils and acting as belowground arthropod attractants [28]; and (2) carbon dioxide, released from the decayed cellulose of sawdust, as well other volatile compounds [16], exert an aggregative impact on the activity of larvae of Melolontha spp. [15,16,29,30].
The hypothesis of this study was that the ‘small sawdust pits’ method allows better determination of the real spring threat of Melolontha spp. to seedlings and better indicates the location of grubs in the plantation, compared to the traditional large pit method used in Poland. Additionally, the impact of weather conditions on a local Melolonthinae population was assessed. The results of this work could be used to modify the traditional method of hazard assessment from grubs in afforestation.
2. Materials and Methods
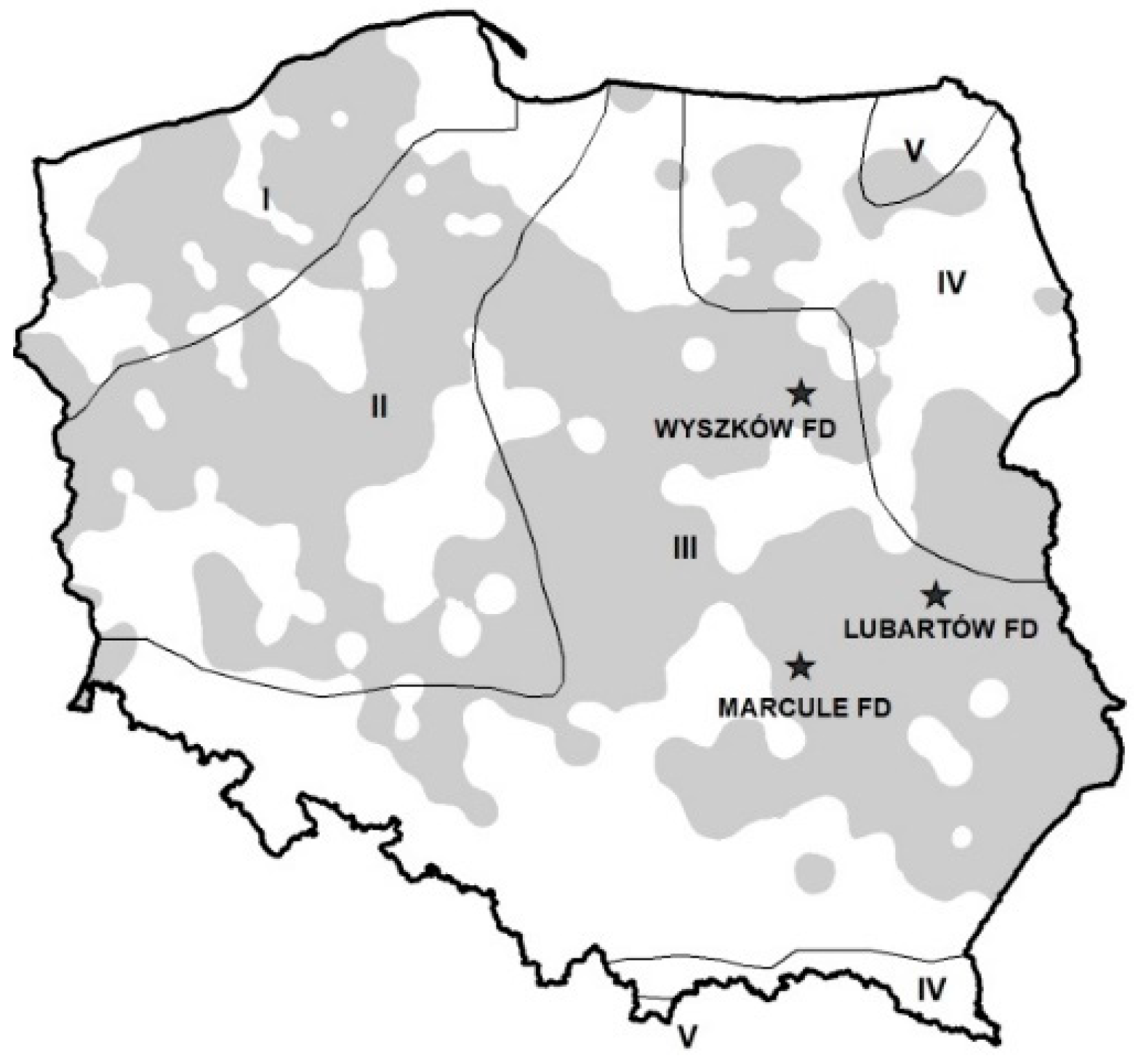
Experimental plots were located in three forest districts (FD): Lubartów (51°23′16.8″ N; 22°37′32.4″ E), Marcule (51°3′58″ N; 21°17′12″ E), and Wyszków (52°43′15″ N; 21°39′03″ E). The field studies were conducted in Lubartów FD in 2012 to 2013, in Marcule FD in 2014 to 2015, and in Wyszków FD in 2016 to 2017. Before planting, the area was an 80 to 100 years old Scots pine in fresh mixed coniferous Oxalis-Myrtillus type (OMT) stands, harvested two years before. The area was partially covered with grass and Myrtillus, and soil was prepared with a LPZ-75 plough. All evaluated forest districts are situated in the Middle European Plain Province of the Middle Europa Megaregion [31], with similar winter temperatures (IIIrd soil freezing class, according to European Act (PN)-EN-12831) (Figure 1), and annual precipitation classified by Bojarczuk et al. [32] as the continental climatic zone.
One year before the investigations, all experimental areas were considered to have a strong threat from cockchafer larvae to intended Scots pine plantations. The threat was based on the high mean number of cockchafer larvae found in soil checked in the autumn of the year preceding afforestation; this evaluation used one “big soil pit”, 1 × 1 × 0.5 m (width/length/depth), for every 5 hectares, according to Polish rules (IOL 2012). The mean number of grubs was approximately 4 three-year grubs/pit, which meant a serious threat and could lead directly to the destruction of the stand [22,33].
In April 2012, 2014, and 2016, respectively, one-year-old seedlings originating from a local open nursery were planted manually, spaced 0.7 m between seedlings in a row and 1.2 m between rows, in each focal area. Within each forest district, three 15-m-long plots were randomly assigned. Each plot contained eight rows of seedlings. In May, 18 holes (0.2 × 0.2 × 0.3 m) were dug between seedlings in each experimental area. The holes, called “small pits”, were spaced evenly in every row (Figure 2). Each hole was filled with fresh, moist Scots pine sawdust (0.2 m deep) and covered with 10 cm local soil. Sawdust was sampled from each pit: (i) At the end of September (for an autumn rating), and (ii) at the end of May (for a spring rating, one year after planting). Samples were sieved through sieves with a mesh size of 2 mm. At the same time, 18 similar holes were dug in the interstices, located 50 cm away from the seedlings and two one side of the adjacent interrow, and the soil from these holes was similarly sieved (Figure 2).
In the Marcule FD in autumn 2014, we performed an additional experiment in three other plots, in two replications. The additional pits were dug in rows: (i) Between seedlings, and (ii) in the nearest inter-row, at a distance of 50 cm from seedlings, and sieved together in autumn with (iii) a similar pit assessment of the soil directly under neighboring seedlings (across these three treatments there were 108 pits altogether, resulting in sieving of a total of 3 × 1.3 m3 of soil).
Larvae remaining on the sieve were counted and compared. The total number of larvae in all small pits, per unit volume (0.012 m3 or 0.216 m3 per each area), was converted to an estimate of the number of larvae expected to occupy the volume of the big soil pits (0.5 m3), according to the Instruction of Forest Protection [22].
The values of monthly air temperature and precipitation in the forest districts for the period of 2011–2017 were obtained courteously from the Institute of Meteorology and Water Management (IMGW Warsaw). For the vegetation growth season (April–October), the values of the Selyaninov hydrothermal coefficient (HTC) were calculated according to the formula:
where P is the monthly sum of precipitation and Σ t is the sum of the average daily temperatures in the month, respectively, in the vegetation growth period, during which the temperature should be above 10 °C [34]. The Selyaninov HTC coefficient is often used in agriculture and horticulture as a synthetic index of the precipitation and temperature dependence in a given period, defining the weather in the range from hot drought (HTC < 1 means drought) to cold and excessive precipitation [35,36].
HTC = 10 × P/Σ t,
Results were analyzed in two ways. First, for every forest district, differences between the number of grubs in control pits and the number of grubs in sawdust pits were determined. In the second part of the analysis, the number of grubs was compared separately between the dates of the performance of small pitches (spring, autumn) for each of the experimental variants. In both cases, the non-parametric Mann-Whitney test was used to evaluate differences, since a Kolmogorov–Smirnov test found that the data deviated significantly from a normal distribution. To evaluate the correlation between the number of grubs and the hydrothermal coefficient, a simple linear regression was used. Analyses were performed in the Statistica v. 13.0 package (Dell Inc., Round Rock, TX, USA).
3. Results
In our research, the Melolontha grubs were counted together, regardless of the species and age, assuming that each individual is responsible for damage to the roots. On the areas tested, M. melolontha 3-years-old grubs dominated; M. hippocastanei were found singly.
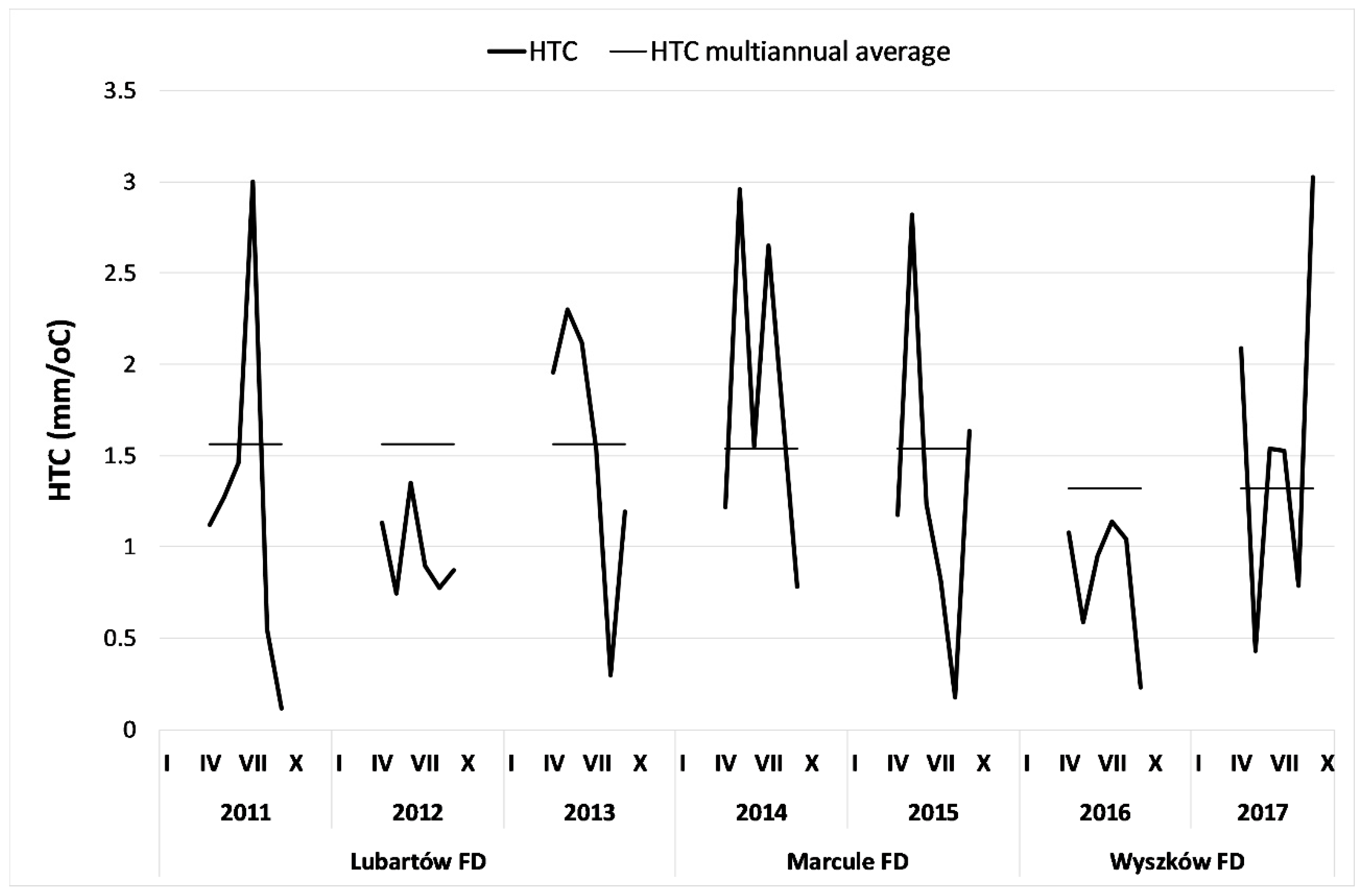
The weather conditions in the period 2011–2017 were highly variable in particular months. Total precipitation in autumn 2012 and spring 2013 was rather low, whereas in spring 2014 it was above the median perennial moisture. In August 2015, precipitation was dramatically low, and the air temperature very high. In spring 2016, weather conditions were rather unfavorable for vegetation (due to low rainfall and relatively high temperatures), and spring 2017 was dry and cold, whereas the autumn of 2017 was wet and warm.
The values of the hydrothermal coefficient (HTC) were generally lower than the multi-annual averages for these areas, especially in the particularly dry years of 2012 and 2016 (Figure 3). In 2014 and 2017, the rainfall was higher and the temperature was rather low, which resulted in more favorable conditions for Scots pine cultures.
The comparison of the number of grubs of all stages and species found in sawdust pits and sawdust-free pits within each time period, assessed separately for FD districts (Figure 4), showed the following relationships:
- In the Lubartów FD, significant differences between spring pits with sawdust and spring pits without sawdust were observed (p-value = 0.038).
- In the Marcule FD, grubs were significantly more common in control pits made in autumn than in pits made in spring (p-value = 0.002); in pits with sawdust, no differences between seasons were found.
- In the Wyszków FD, cockchafer grubs were significantly more common in spring 2017 in small pits with sawdust than in pits without sawdust (p-value = 0.031).
These results suggest that it is important to perform spring assessment in sawdust pits in all forest districts. In Lubartów FD, the number of cockchafer grubs found in sawdust pits in spring was 4 times greater than in autumn of the preceding year, and in Wyszków FD the number was more than 13 times greater. The results showed that the grubs were gathering only in interrows and were not found both in pits with or without sawdust and under seedlings. The total average number of grubs in interrows between pits with sawdust was 13 for plots (5 when standardized to 0.5 m2 [22]), and 3 (1) in interrows between pits without sawdust. For comparison, there were 2 grubs found in the large pit according to the traditional IOL method [22] (personal information from FD).
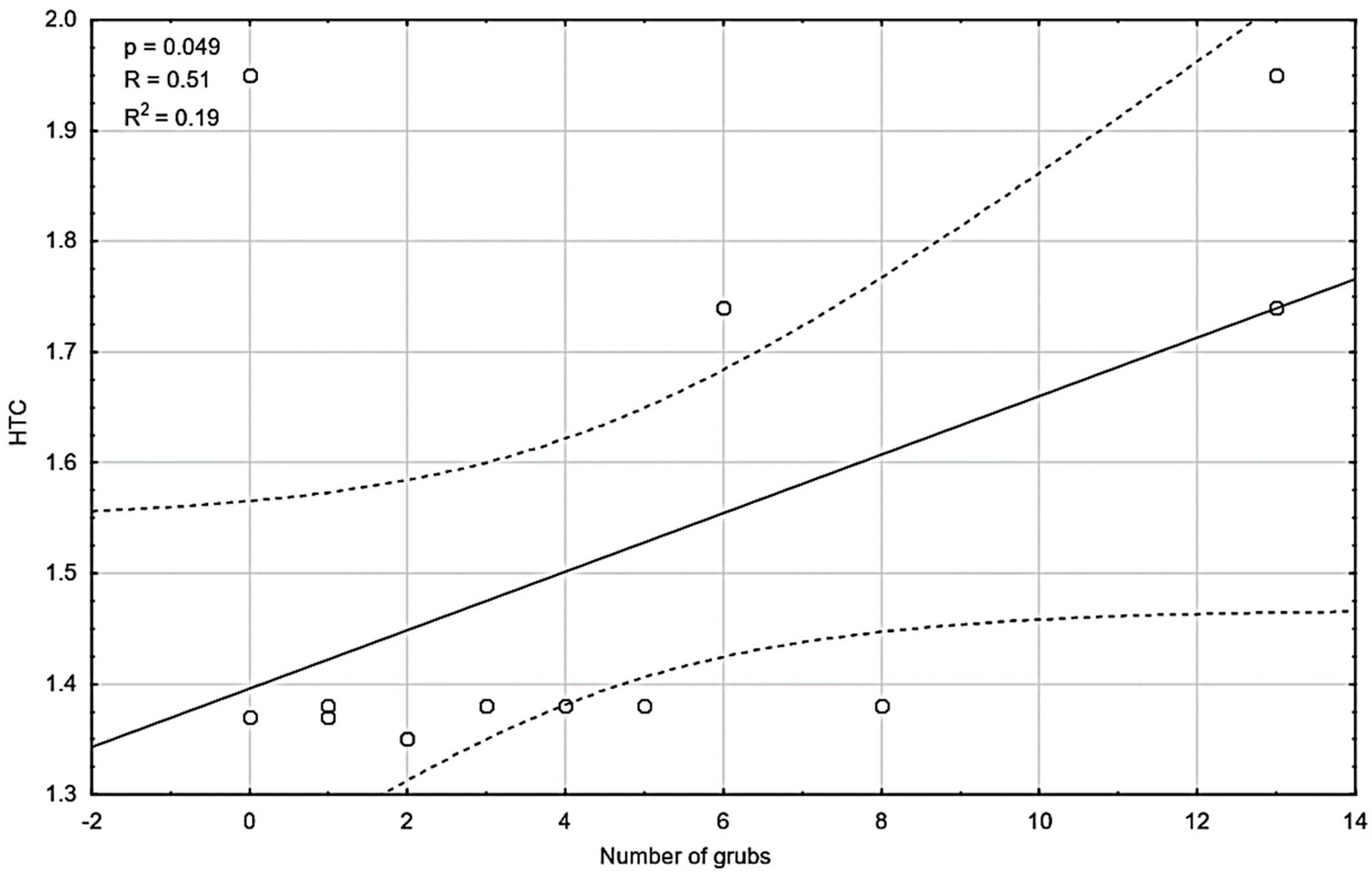
There was a significant relationship (p-value = 0.049) between the occurrence of grubs in the pits and the hydrothermal coefficient. The value of the correlation coefficient was R = 0.51, which indicates the moderate strength of this relationship (Figure 5). The higher value of the hydrothermal coefficient was associated with a higher number of grubs observed in the pits, as result of the better development of plants and their higher attraction.
4. Discussion
Soil is a particularly heterogeneous environment, and so the range of abiotic conditions experienced by root herbivores (e.g., larvae of Melolonthae) can be both diverse and complex. Melolontha spp. are typical eurytopic organisms; their life cycle is dependent on soil moisture and temperature [17]. Barnett and Johnson [37] described that temperature and soil moisture are responsible for movements and changes in the physiological processes of root herbivores. When the temperature is low, metabolic rates decrease, and, as a consequence, larvae are predicted to have decreased movement compared to optimal temperatures [38].
With low rainfall, the grubs in this study showed a tendency to gather in small sawdust pits, where there was probably higher humidity. Similar results were obtained by Lees [39], who found that larvae of the Coleopteran genus, Agriotes, responded in two ways to soil moisture. First, they increased activity and migrated out of dry sand into wet sand, but if soil moisture conditions changed too rapidly, they became trapped and burrowing activity ceased.
In our experiment, we found a significant relationship between the occurrence of grubs in the pits and the hydrothermal coefficient. The higher value of the hydrothermal coefficient was related to a higher number of grubs observed in the pits. This can be explained by the fact that when temperature is low, enzymes react slowly, and, as a consequence, larvae are predicted to display decreased movement compared to times of optimal temperature [38]. An increase of the water content in the soil causes a reduction of oxygen and a significant increase of carbon dioxide levels [37]. This could reduce the movement of most insects not adapted to reduced oxygen contents. In an indirect way, soil moisture can affect the migration of root pests caused by the search for a host plant. Increased soil moisture affects the amount of gaseous molecules, like those released from plant roots and decaying matter, because it can change diffusion rates when compared to standard atmospheric air [40]. Carbon dioxide has been shown to be a major player when it comes to the plant host location by pests [41], and the larvae of Melolontha and many other soil insects are attracted to CO2 [16,29,42]. Carbon dioxide dissolves more quickly in water, which could make it more difficult for pests to move towards the plant roots when soils are saturated.
During this research, we found aggregation of larvae of Melolontha in rows close to seedlings when conditions were unfavorable for growth (during low precipitation in spring 2013 and 2017). These conditions affect the amount and quality of food available for pests and also the plant metabolites released into the rhizosphere under stress. For example, Schenk and Jackson [43] found that the absolute rooting depth of plants in moisture-limited environments has a tendency to increase with increasing levels of precipitation. However, at the same time, some plant species have been shown to increase the size of their rooting systems when soils become drier [44]. Jupp and Newman [45] found that some plant systems increased lateral root growth from 300% to 500% when soil is dry. In contrast, dry soil conditions increase the Scots pine’s susceptibility to soil acidification, and this could significantly reduce fine root growth and increase root mortality [46]. Released in the process of decomposition of dead roots, carbon dioxide may additionally lure the grubs [16].
Opinion is divided on the impact of drought on plant nutrition. Drought may increase the uptake of nutrients by plant root systems through the aggregation of organic and inorganic materials by capillary forces [44]. On the other hand, drought could increase the concentrations of terpenes and resin acids in gymnosperms [47,48]. Terpenes are the major class of compounds to which 17% of insects respond; compounds from this class act as a chemoattractant to insect species, especially those of the order, Coleoptera, and act as potent long-distance signals for foraging insects [49,50,51].
The results obtained here show that the inspection of pits in autumn of the year of cultivation did not offer satisfactory information about the real threat represented by Melolontha spp. to plant cultures in the year following the cultivation. Autumn inspection in the year of cultivation also did not corresponded with results of primary threat assessment in the autumn before spring cultivation. In all cases, inspection performed in the spring of the next year found that cockchafer larvae preferred pits with sawdust over control pits with forest soil and roots. The short period of this research, and the unexpected variability of the weather—exemplified by a dry 2015 followed by a wet 2016—did not allow us to generalize these results beyond the focal experiment. The great number of pits in which there were no grubs suggests that cockchafer larvae occur in clusters. The hypothesis that spring threat assessments and assessment in inter-rows are necessary in order to accurately predict the threat level from grubs was supported. This research showed that a greater number of grubs is correlated with a higher value of the HTC indicator. Sawdust in the pits keeps moisture for a longer time, which is more attractive for grubs in the dry season. The spring assessment period seems to be important due to the migration of grubs toward both developing tree roots with forming mycorrhiza and old, dead roots with residues that give off compounds and indirectly inform grubs of available food [50].
In general, the use of spring risk assessment seems valuable, as indicated by earlier results of mycological [52], bacterial [27], and other sylvicultural research [23,53] in soils subjected to sawdust. Together, these studies confirm that there is increased activity of grubs in these soils after the introduction of sawdust. In addition, small soil pits checked in spring seem to offer more precision for assessment of the severity of the threat of grubs to seedlings, when compared with the more labor-intensive and expensive method prescribed by Polish regulations [22].
5. Conclusions
- A significant relationship between the number of Melolontha spp grubs and the hydrothermal coefficient was found.
- It seems that the pits with sawdust checked in spring allowed the number of grubs to be determined more precisely compared to the assessment performed in autumn.
Author Contributions
M.M. and Z.S. generated the data. All authors analyzed and discussed the data. The manuscript was written by H.S., Z.S. and M.M. Statistical analysis using the Statistica v. 13.0 package by: M.T. The general conception of the project was provided by Z.S.
Funding
This work was supported by the State Forest Holding in Poland through Project No. 500 426. (contract number OR.271.3.4.2015).
Acknowledgments
The authors thank the forest staff of the Lubartów, Marcule, and Wyszków forest districts for valuable assistance during fieldwork. The authors would like to thank the anonymous reviewers for their valuable comments and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hunter, M.D. Root herbivory in forest ecosystems. In Root Feeders, an Ecosystem Perspective; Johnson, S.N., Murray, P.J., Eds.; CAB Biosciences: Ascot, UK, 2008; pp. 68–95. [Google Scholar]
- Johnson, S.N.; Benefer, C.M.; Frew, A.; Griffiths, B.S.; Hartley, S.E.; Karley, A.J.; Rasmann, S.; Schumann, M.; Sonnemann, I.; Robert, C.A.M. New frontiers in belowground ecology for plant protection from root-feeding insects. Appl. Soil Ecol. 2016, 108, 96–107. [Google Scholar] [CrossRef] [Green Version]
- Keller, S.; Brenner, H. Development of the Melolontha populations in the canton Thurgau, eastern Switzerland, over the last 30 years. IOBC/WPRS Bull. 2005, 28, 31. Available online: http://iobc-wprs.org/pub/bulletins/iobc-wprs_bulletin_2005_28_02.pdf#page=46 (accessed on 18 December 2018).
- Oltean, I.; Varga, M.; Gliga, S.; Florian, T.; Bunescu, H.; Bodis, I.; Covaci, A. Monitoring Melolontha melolontha L. species in 2007, in the nursery from UP IV Bătrâna OS Toplița, Harghita forest district. Bull. UASVM CN. 2010, 67, 525. [Google Scholar]
- Švestka, M. Changes in the abundance of Melolontha hippocastani Fabr. and Melolontha melolontha (L.) (Coleoptera: Scarabeidae) in the Czech Republic in the period. J. For. Sci. 2010, 56, 417–428. [Google Scholar] [CrossRef]
- Švestka, M. Ecological conditions influencing the localization of egg-laying by females of the cockchafer (Melolontha hippocastani F.). J. For. Sci. 2007, 53, 16–24. [Google Scholar] [CrossRef]
- Woreta, D.; Sukovata, L. Effect of food on development of the Melolonta hippocastani F. beetles (Coleoptera, Melolonthidae). For. Res. Pap. 2010, 71, 195–199. [Google Scholar] [CrossRef]
- Wagenhoff, E.; Blum, R.; Delb, H. Spring phenology of cockchafers, Melolontha spp. (Coleoptera: Scarabaeidae), in forests of south-western Germany: Results of a 3-year survey on adult emergence, swarming flights, and oogenesis from 2009 to 2011. J. For. Sci. 2014, 60, 154–165. [Google Scholar] [CrossRef]
- Sukovata, L. Szkodniki korzeni drzew i krzewów leśnych [Pests of the roots of trees and shrubs]. In Krótkoterminowa prognoza występowania ważniejszych szkodników i chorób infekcyjnych drzew leśnych w Polsce w 2018 roku [Short-term forecast of the occurrence of major pests and infectious diseases of forest trees in 2018]; Jabłoński Tomasz, Ed.; Forest Research Institute, Analysis and Reports: Sękocin Stary, Poland, 2018; pp. 24–27. Available online: https://www.bdl.lasy.gov.pl/portal/Media/Default/Publikacje/prognoza_2018.pdf (accessed on 1 December 2018).
- Švestka, M. Distribution of tribes of cockchafers of the genus Melolontha in forests of the Czech Republic and the dependence of their swarming on temperature. J. For. Sci. 2006, 52, 520–530. [Google Scholar] [CrossRef]
- Sierpinska, A. Observations on ecology of common cockchafer (Melolontha melolontha L.) and forest cockchafer (Melolontha hippocastani Fabr.)—Based on the research conducted in forest district Piotrkow in 2007. Prog. Plant Prot. 2008, 3, 956–965. [Google Scholar]
- Sukovata, L.; Jaworski, T.; Karolewski, P.; Kolk, A. The performance of Melolontha grubs on the roots of various plant species. Turkish J. Agric. For. 2015, 39, 107–116. [Google Scholar] [CrossRef]
- Blum, M.S. Fundamentals of Insects Physiology; Wiley: New York, NY, USA, 1985; ISBN 0471054682. [Google Scholar]
- Malinowski, H. Current problems of forest protection connected with the control of cockchafers (Melolontha spp.). Prog. Plant Prot. w Ochr. Roślin 2007, 47, 314–322. [Google Scholar]
- Galbreath, R.A. Orientation of grass grub Costelytra zealandica (Coleoptera: Scarabaeidae) to a carbon dioxide source. N. Z. Entomol. 1988, 11, 6–7. [Google Scholar] [CrossRef]
- Weissteiner, S.; Huetteroth, W.; Kollmann, M.; Weißbecker, B.; Romani, R.; Schachtner, J.; Schütz, S. Cockchafer larvae smell host root scents in soil. PLoS ONE 2012, 7, e45827. [Google Scholar] [CrossRef] [PubMed]
- Sierpiński, Z. Ważniejsze owady-szkodniki korzeni drzew i krzewów leśnych; Państwowe Wydawnictwo Rolnicze i Leśne: Warszawa, Poland, 1975. [Google Scholar]
- Ansari, M.A.; Shah, F.A.; Tirry, L.; Moens, M. Field trials against Hoplia philanthus (Coleoptera: Scarabaeidae) with a combination of an entomopathogenic nematode and the fungus Metarhizium anisopliae CLO 53. Biol. Control 2006, 39, 453–459. [Google Scholar] [CrossRef]
- Bailey, P.T. Pests of Field Crops and Pastures: Identification and Control; CSIRO publishing: Collingwood, Australia, 2007; ISBN 0643099425. [Google Scholar]
- Benker, U.; Leuprecht, B. Field experience in the control of common cockchafer in the Bavarian region Spessart. IOBC/WPRS Bull. 2005, 28, 21–24. [Google Scholar]
- Pernfuss, B.; Zelger, R.; Kron-Morelli, R.; Strasser, H. Control of the garden chafer Phyllopertha horticola with GranMet-P, a new product made of Metarhizium anisopliae. IOBC/WPRS Bull. 2005, 28, 47–50. [Google Scholar]
- Instrukcja ochrony lasu [Instruction of Forest Protection]; CILP: Warsaw, Poland, 2012; Available online: https://www.lasy.gov.pl/pl/publikacje/copy_of_gospodarka-lesna/ochrona_lasu/instrukcja-ochrony-lasu/instrukcja-ochrony-lasu-tom-i/view and https://www.lasy.gov.pl/pl/publikacje/copy_of_gospodarka-lesna/ochrona_lasu/instrukcja-ochrony-lasu/instrukcja-ochrony-lasu-tom-ii/view; (accessed on 1 December 2018).
- Małecka, M.; Sierota, Z.; Tarwacki, G. Effect of sawdust addition into one−year−old Scots pine plantation on number of Melolontha grubs. Sylwan 2014, 158, 604–613. [Google Scholar]
- Bååth, E.; Söderström, B. Degradation of macromolecules by microfungi isolated from different podzolic soil horizons. Can. J. Bot. 1980, 58, 422–425. [Google Scholar] [CrossRef]
- Svarstad, H.; Bugge, H.C.; Dhillion, S.S. From Norway to Novartis: Cyclosporin from Tolypocladium inflatum in an open access bioprospecting regime. Biodivers. Conserv. 2000, 9, 1521–1541. [Google Scholar] [CrossRef]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi, 2nd ed.; IHW Verlag: Eching, Germany, 2007. [Google Scholar]
- Kubiak, K.; Tkaczyk, M.; Małecka, M.; Sierota, Z. Pine sawdust as stimulator of the microbial community in post-arable afforested soil. Arch. Agron. Soil Sci. 2017, 63, 427–441. [Google Scholar] [CrossRef]
- Wenke, K.; Kai, M.; Piechulla, B. Belowground volatiles facilitate interactions between plant roots and soil organisms. Planta 2010, 231, 499–506. [Google Scholar] [CrossRef]
- Johnson, S.N.; Gregory, P.J. Chemically-mediated host-plant location and selection by root-feeding insects. Physiol. Entomol. 2006, 31, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Weissteiner, S.; Schütz, S. Are different volatile pattern influencing host plant choice of belowground living insects. Mitt. Dtsch. Ges. Allg. Angew. Ent. 2006, 15, 51–55. [Google Scholar]
- Kondracki, J. Geografia Regionalna Polski [Physico-geographical regionalization of Poland]. Państwowe Wydaw. Nauk. Warszawa 2002, 441. [Google Scholar]
- Bojarczuk, T.; Bugala, W.; Chylarecki, H. Zrejonizowany dobór drzew i krzewów do uprawy w Polsce. Arbor. Kórnickie 1980. [Google Scholar]
- Niemczyk, M.; Karwañski, M.; Grzybowska, U. Effect of environmental factors on occurrence of cockchafers (Melolontha spp.) in forest stands. Baltic For. 2017, 23, 334–341. [Google Scholar]
- Żmudzka, E. The climatic background of agricultural production in Poland (1951–2000). Misc. Geogr. 2004, 11, 127–137. [Google Scholar] [CrossRef]
- Evarte-Bundere, G.; Evarts-Bunders, P. Using of the hydrothermal coefficient (HTC) for interpretation of distribution of non-native tree species in Latvia on example of cultivated species of genus Tilia. Acta Biol. Univ. Daugavp 2012, 12, 135–148. [Google Scholar]
- Riaznova, A.A.; Voropay, N.N.; Okladnikov, I.G.; Gordov, E.P. Development of computational module of regional aridity for web-GIS “Climate”. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Tomsk, Russia, 11–16 July 2016; IOP Publishing: Bristol, UK, 2016; p. 012032. [Google Scholar]
- Barnett, K.; Johnson, S.N. Living in the soil matrix: Abiotic factors affecting root herbivores. In Advances in Insect Physiology; Johnson, S.N., Hiltpold, I., Turlings, T.C.J., Eds.; Elsevier: Oxford, UK, 2013; Volume 45, pp. 1–52. ISBN 0065-2806. [Google Scholar]
- Edwards, A.C.; Cresser, M.S. Freezing and its effect on chemical and biological properties of soil. In Advances in Soil Science; Springer: Berlin, Germany, 1992; pp. 59–79. [Google Scholar]
- Lees, A.D. On the behaviour of wireworms of the genus Agriotes Esch. (Coleoptera, Elateridae): I. Reactions to humidity. J. Exp. Biol. 1943, 20, 43–53. [Google Scholar]
- Payne, D.; Gregory, P.J. The Soil Atmosphere. Russell’s Soil Cond. Plant Growth; Longman: Harlow, UK, 1988; pp. 298–314. [Google Scholar]
- Klingler, J. On the orientation of plant nematodes and of some other soil animals. Nematologica 1965, 11, 4–18. [Google Scholar] [CrossRef]
- Bernklau, E.J.; Bjostad, L.B. Reinvestigation of host location by western corn rootworm larvae (Coleoptera: Chrysomelidae): CO2 is the only volatile attractant. J. Econ. Entomol. 1998, 91, 1331–1340. [Google Scholar] [CrossRef]
- Schenk, H.J.; Jackson, R.B. Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. J. Ecol. 2002, 90, 480–494. [Google Scholar] [CrossRef] [Green Version]
- Gregory, P.J. Roots, rhizosphere and soil: The route to a better understanding of soil science? Eur. J. Soil Sci. 2006, 57, 2–12. [Google Scholar] [CrossRef]
- Jupp, A.P.; Newman, E.I. Morphological and anatomical effects of severe drought on the roots of Lolium perenne L. New Phytol. 1987, 105, 393–402. [Google Scholar] [CrossRef]
- Vanguelova, E.I.; Nortcliff, S.; Moffat, A.J.; Kennedy, F. Morphology, biomass and nutrient status of fine roots of Scots pine (Pinus sylvestris) as influenced by seasonal fluctuations in soil moisture and soil solution chemistry. Plant Soil 2005, 270, 233–247. [Google Scholar] [CrossRef]
- Tang, C.-S.; Cai, W.-F.; Kohl, K.; Nishimoto, R.K. Plant Stress and Allelopathy; American Chemical Society: Washington, DC, USA, 1995. [Google Scholar]
- Turtola, S.; Manninen, A.-M.; Rikala, R.; Kainulainen, P. Drought stress alters the concentration of wood terpenoids in Scots pine and Norway spruce seedlings. J. Chem. Ecol. 2003, 29, 1981–1995. [Google Scholar] [CrossRef]
- Dicke, M. Local and systemic production of volatile herbivore-induced terpenoids: Their role in plant-carnivore mutualism. J. Plant Physiol. 1994, 143, 465–472. [Google Scholar] [CrossRef]
- Eilers, E.J.; Talarico, G.; Hansson, B.S.; Hilker, M.; Reinecke, A. Sensing the underground—Ultrastructure and function of sensory organs in root-feeding Melolontha melolontha (Coleoptera: Scarabaeinae) larvae. PLoS ONE 2012, 7, e41357. [Google Scholar] [CrossRef]
- Hiltpold, I.; Bernklau, E.; Bjostad, L.B.; Alvarez, N.; Miller-Struttmann, N.E.; Lundgren, J.G.; Hibbard, B.E. Nature, evolution and characterisation of rhizospheric chemical exudates affecting root herbivores. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 45, pp. 97–157. ISBN 0065-2806. [Google Scholar]
- Kwaśna, H.; Sierota, Z.; Bateman, G.L. Fungal communities in fallow soil before and after amending with pine sawdust. Appl. Soil Ecol. 2000, 14, 177–182. [Google Scholar] [CrossRef]
- Szmidla, H.; Tkaczyk, M.; Monika, M.; Sierota, Z. Assessing the number of Melolonthinae larvae in the sawdust traps in young Scots pine (Pinus sylvestris L.) plantations. Sylwan 2018, 162, 590–597. [Google Scholar]
Figure 1.
Area of occurrence of Melolonthinae in Poland in 2017 (grey), borders of climatic zones (lines), and locality of experimental plots (stars) in tested forest districts (FD).
Figure 1.
Area of occurrence of Melolonthinae in Poland in 2017 (grey), borders of climatic zones (lines), and locality of experimental plots (stars) in tested forest districts (FD).

Figure 2.
Location of small pits with sawdust (a) between seedlings in the row, and control pit without sawdust (b) in the interrow.
Figure 2.
Location of small pits with sawdust (a) between seedlings in the row, and control pit without sawdust (b) in the interrow.

Figure 3.
Monthly values of the hydrothermal coefficient (HTC) for research areas (source data: Institute of Meteorology and Water Management Bulletin).
Figure 3.
Monthly values of the hydrothermal coefficient (HTC) for research areas (source data: Institute of Meteorology and Water Management Bulletin).

Figure 4.
Comparison of the numbers of cockchafers found in pits of different treatments between evaluated periods at particular forest districts (FD).
Figure 4.
Comparison of the numbers of cockchafers found in pits of different treatments between evaluated periods at particular forest districts (FD).

Figure 5.
Relationship between the hydrothermal coefficient (HTC) and the total number of grubs in pits.
Figure 5.
Relationship between the hydrothermal coefficient (HTC) and the total number of grubs in pits.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Szmidla, H.; Małecka, M.; Tkaczyk, M.; Tarwacki, G.; Sierota, Z. The Spring Assessing Method of the Threat of Melolontha spp. grubs for Scots Pine Plantations. Forests 2019, 10, 399. https://0-doi-org.brum.beds.ac.uk/10.3390/f10050399
AMA Style
Szmidla H, Małecka M, Tkaczyk M, Tarwacki G, Sierota Z. The Spring Assessing Method of the Threat of Melolontha spp. grubs for Scots Pine Plantations. Forests. 2019; 10(5):399. https://0-doi-org.brum.beds.ac.uk/10.3390/f10050399
Chicago/Turabian StyleSzmidla, Hanna, Monika Małecka, Miłosz Tkaczyk, Grzegorz Tarwacki, and Zbigniew Sierota. 2019. "The Spring Assessing Method of the Threat of Melolontha spp. grubs for Scots Pine Plantations" Forests 10, no. 5: 399. https://0-doi-org.brum.beds.ac.uk/10.3390/f10050399
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.