Chemosensory Characteristics of Two Semanotus bifasciatus Populations


Key Laboratory of Forest Protection, Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China
*
Author to whom correspondence should be addressed.
Forests 2019, 10(8), 655; https://0-doi-org.brum.beds.ac.uk/10.3390/f10080655
Submission received: 9 July 2019
/
Revised: 1 August 2019
/
Accepted: 1 August 2019
/
Published: 2 August 2019
(This article belongs to the Special Issue Impacts, Monitoring and Management of Forest Pests and Diseases)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Semanotus bifasciatus (Motschulsky) (Cerambycidae: Coleoptera) is a major forest borer in China, and attractants provide a promising method for the control of this pest. Exploration of the chemosensory mechanisms of S. bifasciatus is important for the development of efficient attractants for this pest. However, little information is available about the olfactory mechanisms of S. bifasciatus. Previous research has indicated that the trapping effects of the same attractant are different between Beijing and Shandong populations of S. bifasciatus. To explore the reasons for this, next-generation sequencing was performed to analyze the antennal transcriptome of both sexes of the two S. bifasciatus populations, and the olfactory-related genes were identified. Furthermore, the expression levels and single nucleotide polymorphisms (SNPs) of the olfactory-related genes between the two populations were compared. We found that the expression levels of odorant binding proteins (OBPs), odorant receptors (ORs), and sensory neuron membrane proteins (SNMPs) in male S. bifasciatus of the Beijing population were obviously lower than those in the Shandong population, and most of the conserved SNPs in OBPs and ORs of the two populations showed more diversity in the Beijing population. Our work provides a foundation for future research of the molecular olfactory mechanisms and pest management of S. bifasciatus, as well as other longhorn beetles.
1. Introduction
Semanotus bifasciatus (Motschulsky) mainly damages Platycladus orientalis (L.) Franco, Sabina chinensis (L.) Antoine, and other species, and the damage it causes is very serious in some regions [1], which makes this pest a main forest borer species in China. In recent years, the protection of old cypress trees, forest resources, and the ecological environment has been threatened by S. bifasciatus in northern China. For example, 67.3% of planted seedlings were damaged by S. bifasciatus in Dalian, Liaoning province in 1988 [2]; about 90% of dead cypresses were infested by S. bifasciatus in the Lingyan Forest district of Taian, Shandong province in the period 2002–2006; and 97% of the introduced cypress were damaged by S. bifasciatus in Xiangshan Mountain park, Beijing in 1979. Thus, the control of S. bifasciatus is urgent and important for forest protection.
However, as a boring insect, S. bifasciatus remain concealed during egg and larval stages, so it is hard to control this pest. The adult form of S. bifasciatus appears in the forest for about half a month only, and at this stage, we can use treatments on them relatively easily. Thus, developing attractants for S. bifasciatus based on volatiles or pheromones has attracted the interest of many forest insect scientists, and some volatile lures, using volatiles from certain host plants, have been used to trap this pest [3,4]. The attracting effect of an attractant was tested at two different places—Beijing and Shandong, China—and the results indicated that the responses of two S. bifasciatus populations from these two places differed greatly (unpublished data). This phenomenon indicated that the olfactory sensibilities of the two S. bifasciatus populations were different, and this deserves further research. The results of such research may help us to develop accurate attractants for different pest populations.
However, the olfactory response mechanisms of S. bifasciatus are unclear, and only fragmented information has been available until now [5]. Thus, our work focuses on the olfactory variety of S. bifasciatus from different places.
In insects, environmental odors are mainly detected by olfactory sensory neurons (OSNs) located in sensilla on the antennae [6]. During transformation of chemical volatile signals to electrical nervous impulses, insects rely on at least six olfactory-related gene groups, including two binding protein families, three receptor families, and the sensory neuron membrane proteins (SNMPs) [7,8]. The three receptor families expressed in chemosensory neurons [9,10,11], include odorant receptors (ORs), ionotropic receptors (IR), and gustatory receptors (GRs) [12,13]. Insect ORs are seven-transmembrane domain proteins, and their topologies are reversed, comparing to vertebrate ORs [14,15]. ORs always appear as dimers, comprised by one specific OR [16] and one conserved gene, called the odorant receptor co-receptor (Orco) [17]. Most GRs are expressed in gustatory receptor neurons, which are involved in contact chemoreception [18]. IRs refer to receptors that are related to ionotropic glutamate receptors (iGluRs) [9]. In addition to the receptor genes, there are two types of binding proteins in olfactory systems that play critical roles in olfaction. Odorant binding proteins (OBP) are small, soluble proteins with six conserved cysteines [19,20]. OBPs (odorant binding proteins) have been proposed to deliver odor molecules to the receptors [19]. Chemosensory proteins (CSPs) are another type of small binding protein, which are more conserved, and they are characterized by two disulfide bridges formed by the four conserved cysteines [21].
Insects display a highly specific and sensitive olfactory sense and complex olfactory behavior, and their olfaction-related genes show large sequence diversity [22,23,24,25]. Therefore, identification of olfactory-related genes mainly relies on genomic data [26,27,28]. Recently, studies on antennal transcriptomes have led to the identification of olfactory-related genes in many insects [29,30,31,32,33,34], which demonstrated the power of transcriptomic data for olfactory gene identification. However, few studies have compared the olfactory recognition mechanisms between different populations of the same insect species.
In this study, to explore the olfactory variety of different S. bifasciatus populations, we sequenced the antennal transcriptomes of both sexes from two places, identified the olfactory-related genes of S. bifasciatus, and compared the expression quantity and single nucleotide polymorphisms (SNPs) between the two populations.
2. Materials and Methods
2.1. Insects for Antennal Transcriptome Sequencing
Sections of wood containing overwintering S. bifasciatus insects were collected from two sites, the Lingyan Forest district of Taian, Shandong province, and the Mentougou district of Beijing, in March 2018, and were placed in 26 °C ± 2 °C temperature with 50% ± 10% relative humidity and a 16 h light, 8 h dark photo-period. The wood cross-sections were sealed with wax. Eclosion was seen about two weeks later. After eclosion, antennae from 15 female and male S. bifasciatus from the two sites were cut off at the base, and the samples were frozen in liquid nitrogen immediately. Antenna from five S. bifasciatus were collected as one sample, and a total of three biological replicates were obtained.
2.2. RNA-Seq Library Preparation and Sequencing
We extracted the total RNA of S. bifasciatus antenna with TRIzol reagent (Invitrogen, Carlsbad, CA, USA), as previously described [8,35], and then treated them with RNase-free DNase I (TaKaRa, Dalian, Liaoning, China). The RNA purity was checked using the NanoPhotometer® spectrophotometer (IMPLEN, Westlake Village, CA, USA), concentration with Qubit® 2.0 Fluorometer (Life Technologies, Carlsbad, CA, USA), and integrity on the Bioanalyzer 2100 system (Agilent Technologies, Santa Clara, CA, USA), respectively.
A total amount of 1 μg RNA per sample was used to synthetize complementary DNA (cDNA) [36,37]. Then, sequencing libraries were prepared with an Illumina TruSeq™ RNA Sample Preparation Kit (Illumina, San Diego, CA, USA). An AMPure XP system (Beckman Coulter, Beverly, MA, USA) was used to select cDNA fragments (of 200 bp length, preferentially), and two-end adaptor ligated DNA fragments were enriched with polymerase chain reaction (PCR). Finally, the products were purified and quantified (Agilent Bioanalyzer 2100). The index-coded samples were clustered and sequenced with an Illumina Hiseq 2500, according to the manufacturer’s instructions.
2.3. De Novo Assembly of the Sequences
Clean data were obtained from raw sequencing data, using the following treatments: with self-written Perl scripts, we removed reads containing an adapter sequence, uncertain bases (reads with >10% N), and sequences more than 50% bases with error rates >1%. Pollution of the data was ruled out by comparing with the NT database. Assembly was performed with Trinity (version: trinityrnaseq-r20131110) [38], and redundancy was treated with TGICL [39].
2.4. Annotation
All unigenes were aligned with Blast (The Basic Local Alignment Search Tool) against the NR, SWISSPROT, KEGG, and KOG databases (cut-off by 1e-5), and we selected the highest sequence similarity targets as the functional annotation of the transcripts. Then, Gene Ontology (GO) annotation was performed with Blast 2GO [40,41], and the molecular function, biological process, and cellular component of the genes were assigned [42].
We identified the chemosensory genes in S. bifasciatus using contig tBLASTx searches with a query sequence of chemosensory genes from other insects, such as Drosophila melanogaster, Bombyx mori, and other long-horned beetles. Open reading frames (ORFs) of the candidate genes were found, which were further verified using BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
2.5. Gene Expression Quantification
FPKM (fragments per kilobase of exon per million fragments mapped) values were used to measure the expression level of the genes [43]. FPKM value and standard error were obtained from the three biological replicates. Differentially expressed genes (DEGs) of the two S. bifasciatus populations were selected using DESeq (http://bioconductor.org/packages/release/bioc/html/DESeq.html) [44]. The compatible-hits-norm model was used to normalize the Unigene expression levels and DEGs [45], and the false discovery rate (FDR) method (p ≤ 0.05) was used to identify DEGs [46].
2.6. SNP Calling
We used the Genome Analysis Toolkit (GATK2) software to carry out SNP calling [47]; the GATK standard filter method and other parameters were used to filter the Raw vcffiles (cluster Window Size: 35; MQ0 ≥ 4; and (MQ0/(1.0 × DP)) > 0.1, QUAL < 10, QUAL < 30.0, or QD < 5.0, or HRun > 5) and retain the SNPs with distance >5. The SNPs that showed stable diversity between the two S. bifasciatus populations or between sexes were selected as conserved SNPs.
2.7. Data Analysis
The expression levels of the identified chemosensory genes were compared according to the FPKM values and based on the three biological replicates. Significant difference between the two populations were tested using ANOVA test (in SPSS 18, SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Antennal Transcriptomes Assembly and Annotation
The transcriptomic sequence data were generated using an antenna cDNA library and Illumina HiSeq TM2000/MiSeq technology. About 6 Gbp of clean sequence data were obtained for each sample. Assembly resulted in 55,355 contigs, with an N50 of 2813 bp. The number of contigs longer than 1 kb was 29,791. Gene annotation to several databases obtained gene annotation information for 35,188 genes.
3.2. Chemosensory Gene Identification of S. bifasciatus
We focused on olfactory gene families to perform a detailed analysis of the transcriptomes. In total, 18 OBPs, 21 CSPs, 66 ORs, 24 GRs, 14 IRs, and 4 SNMPs in S. bifasciatus were identified (Supplementary Table S1).
As the trapping effects of the attractant differed obviously between Shandong and Beijing, we further analyzed the possible reasons for this situation. As the expression levels and gene sequence variations were two factors that may have been related to the trapping effects, we further analyzed the differences of these aspects in the two S. bifasciatus populations.
3.3. Chemosensory Gene Expression Characteristics of Two S. bifasciatus Populations
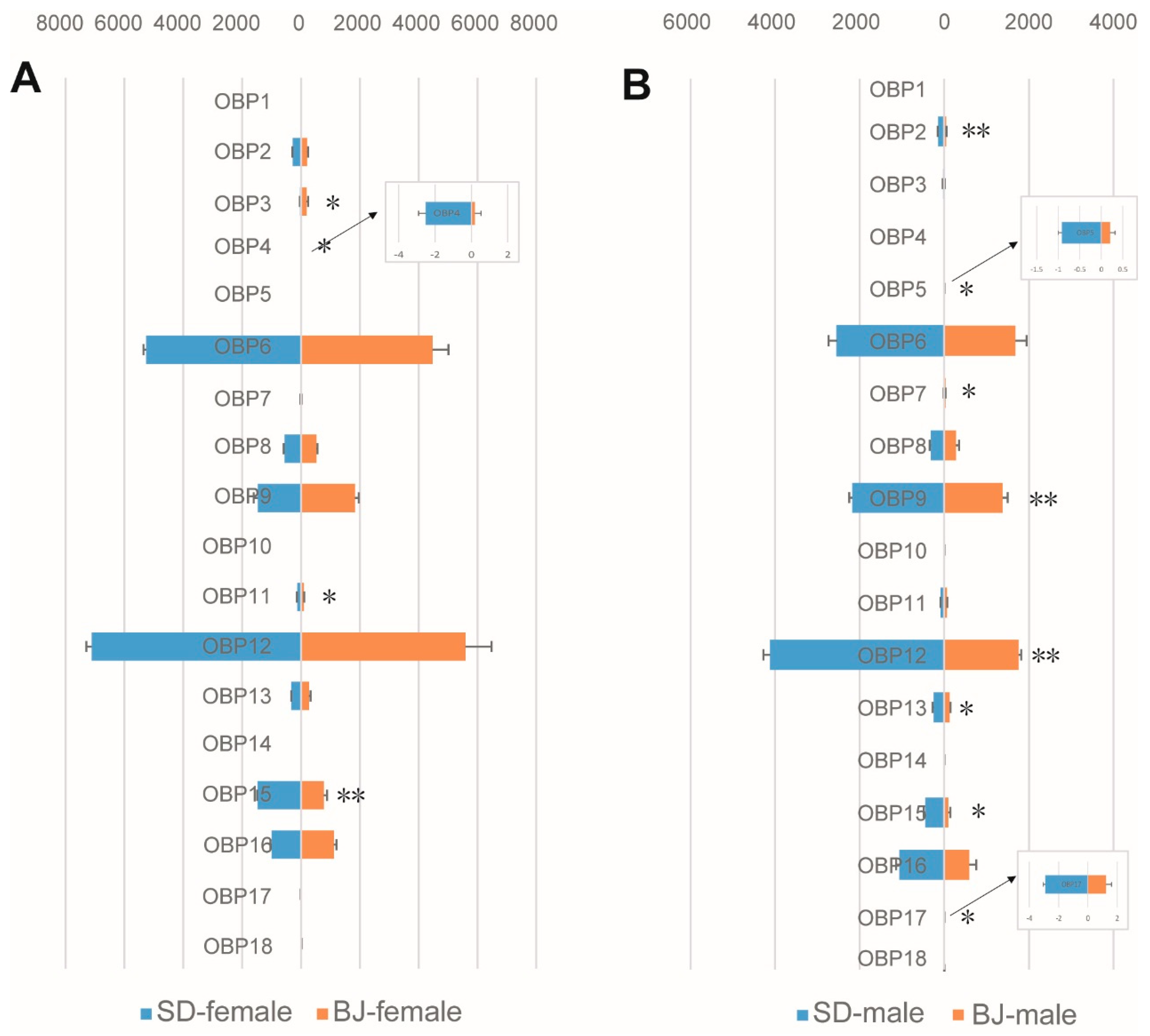
Chemosensory gene expressions of the two S. bifasciatus populations in both sexes were analyzed. Two types of binding proteins (OBPs and CSPs) related to chemosensory were compared between the two populations firstly. Among the 18 identified OBPs, 4 OBPs (OBP3, OBP4, OBP11, and OBP15) were expressed significantly differently between females of the two populations (Figure 1A), while 8 (OBP2, OBP5, OBP7, OBP9, OBP12, OBP13, OBP15, and OBP17) were expressed significantly differently between males of the two populations (Figure 1B). These results indicate that more gene expression varieties were found in males than females of the two populations. A different situation was detected in CSPs, another binding protein of the olfactory system (Figure S1), which means that only a few genes (2 between females and 3 between males of the two populations) were significantly expressed between the two populations.
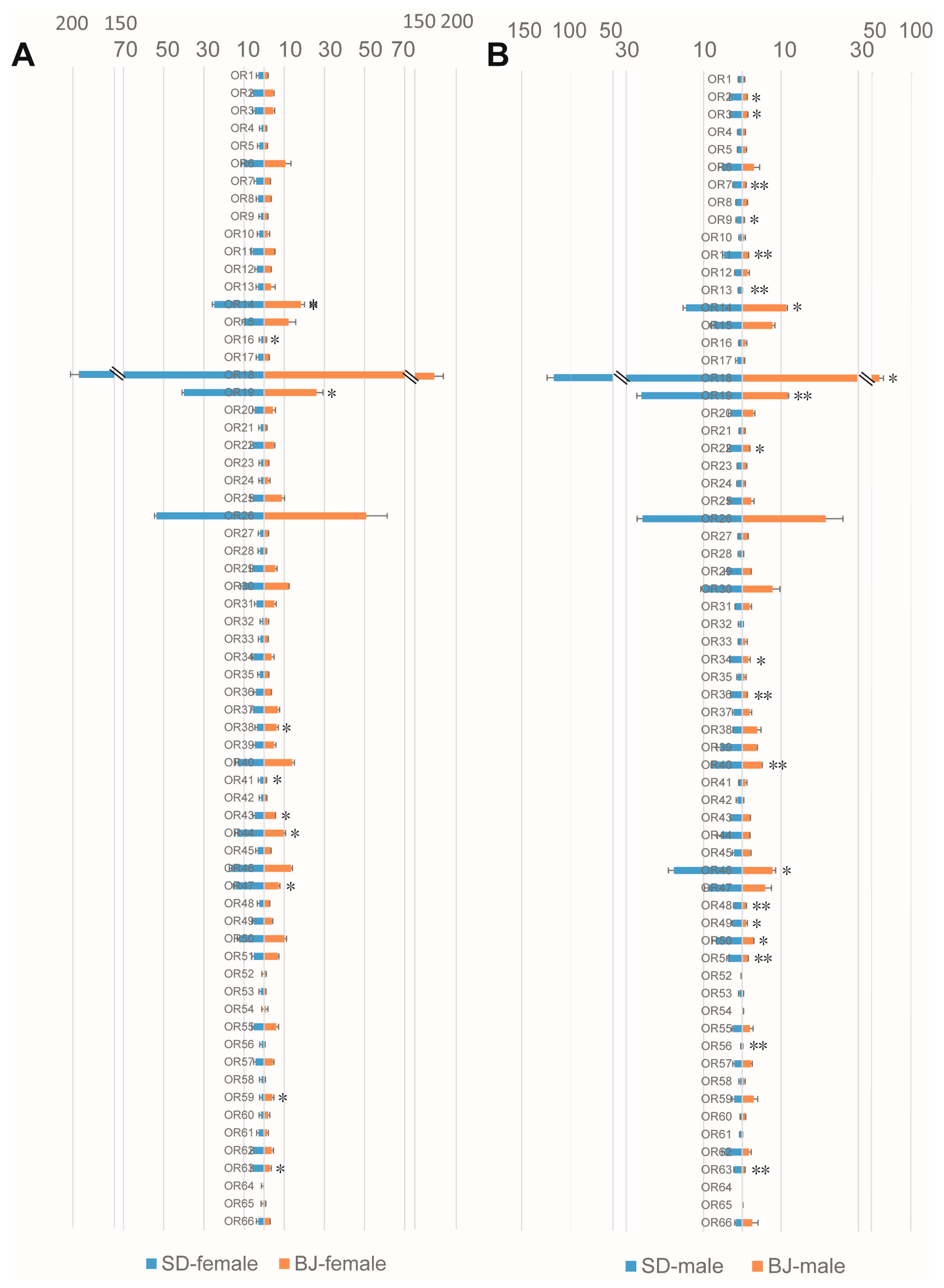
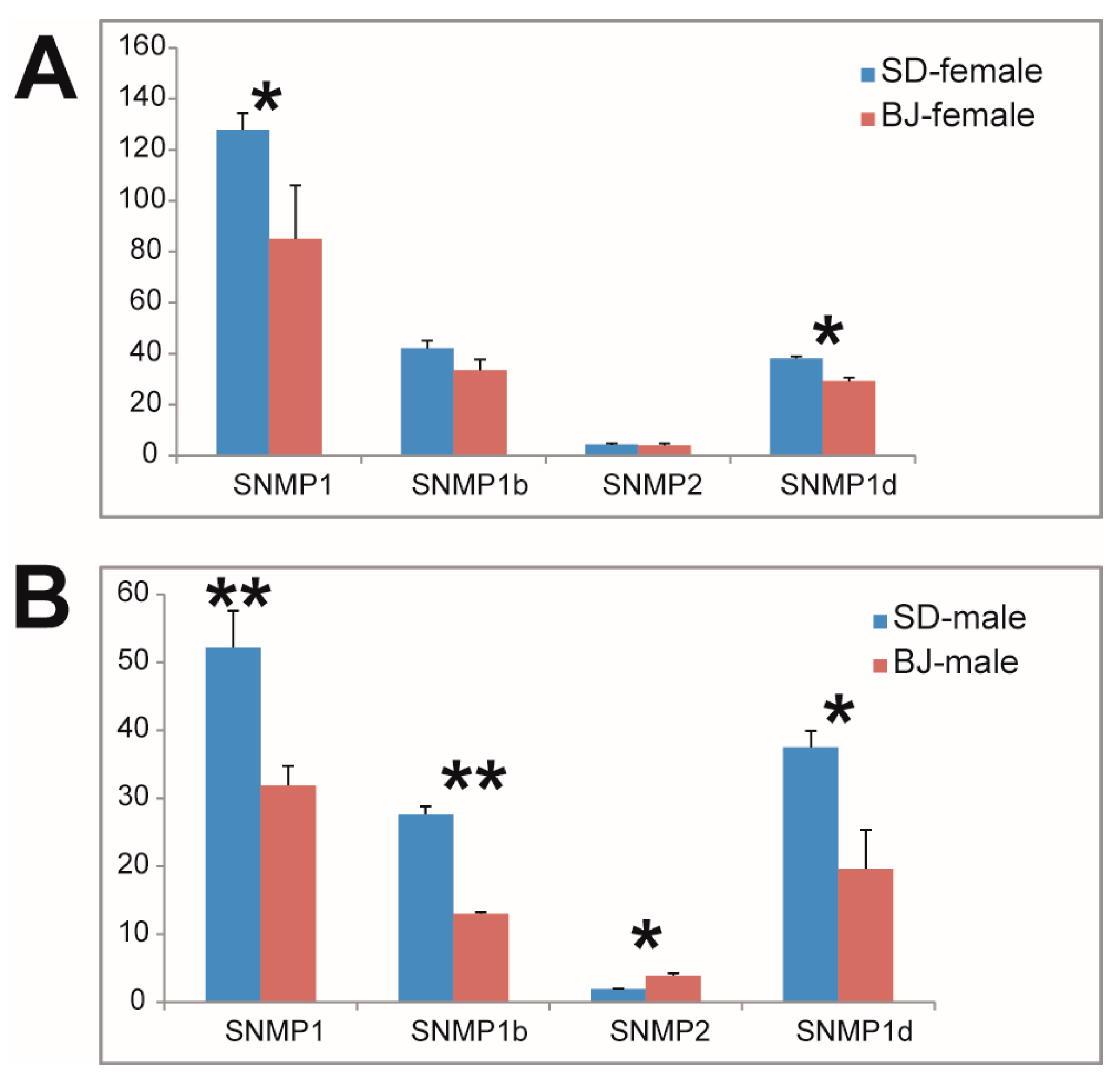
Among the three type of receptor genes related to chemosensory, we found that the ORs possessed most gene expression differences. Similar to OBPs, varieties of males were more than that of females between the two populations: 10 ORs (OR14, OE16, OR19, OR38, OR41, OR43, OR44, OR47, OR59, and OR63) were expressed significantly differently between females of the two populations (Figure 2A), while 20 ORs (OR2, OR3, OR7, OR9, OR11, OR13, OR14, OR18, OR19, OR22, OR34, OR36, OR40, OR46, OR48, OR49, OR50, OR51, OR56, and OR63) were expressed significantly differently between males of the two populations (Figure 2B). The expression differences of GRs and IRs in the two populations were also compared (Figures S2 and S3), and we found that the differently expressed genes of these two types of receptor were less than that of ORs. Three (in females) and 6 (in males) GRs were differently expressed between the two populations (Figure S2), and 2 (in females) and 4 (in males) IRs were expressed differently between the two populations (Figure S3). It is worth mentioning that SNMPs expressed obviously different in male antenna from the two populations, which means that all the four identified SNMPs expressed significantly differently between males from the two populations (Figure 3). These differently expressed genes may have mediated the olfactory differences of the two S. bifasciatus populations.
3.4. SNP Analysis of Chemosensory Genes in Two S. bifasciatus Populations
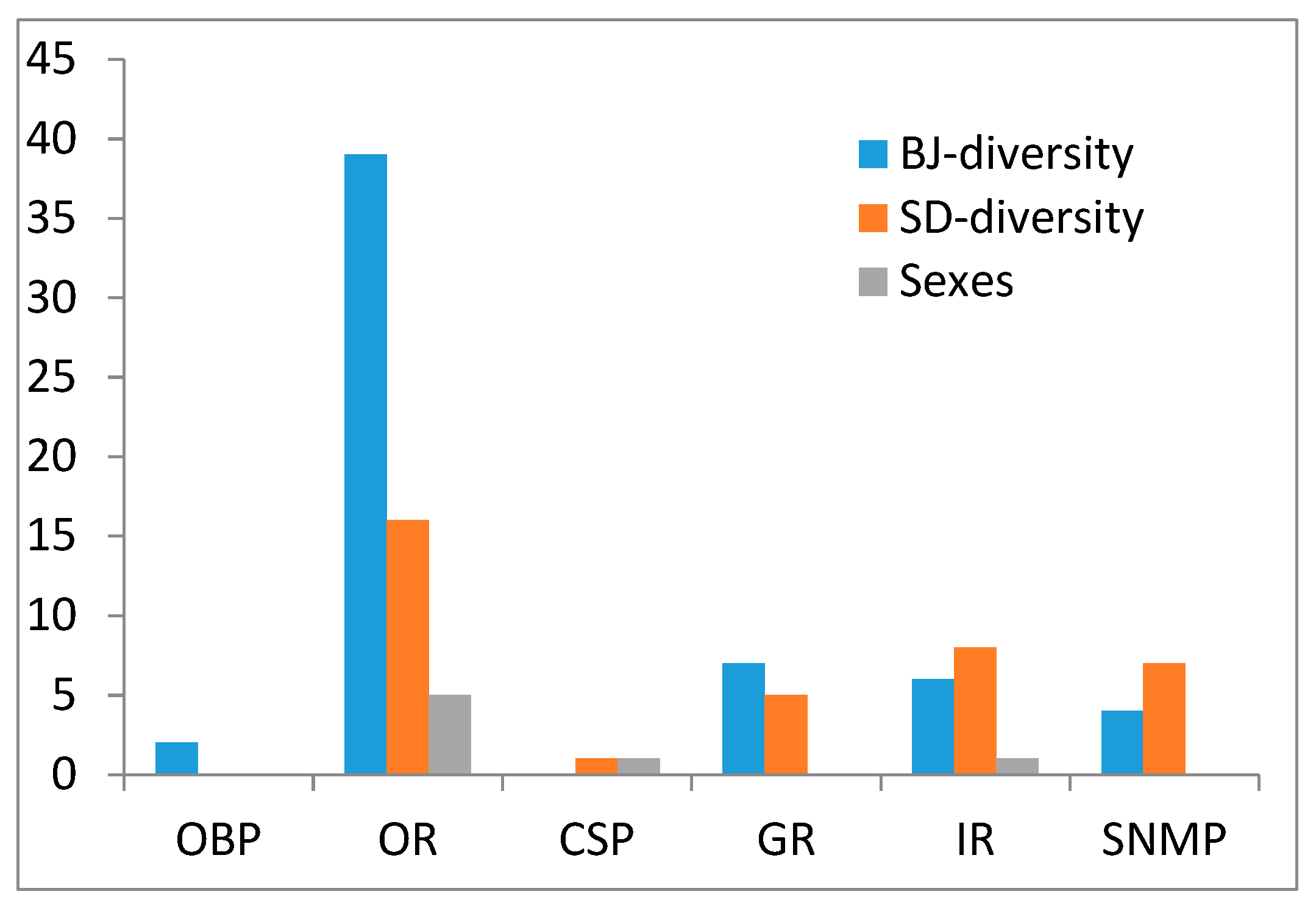
We collected the conserved SNP sites of the chemosensory genes of both sexes in the two S. bifasciatus populations, where 2 SNPs in OBPs, 2 SNPs in CSPs, 63 SNPs in ORs, 12 SNPs in GR, 16 SNPs in IRs, and 11 SNPs in SNMPs were obtained (Supplementary Table S2). Then we analyzed the SNP sites of each gene. If the acid bases at the SNP site were the same in the three repeats of Shandong population but different in that of the Beijing population, we record this SNP site as Beijing diversity SNP site; reversely, if the acid bases at the SNP site were the same in the three repeats of the Beijing population but different in that of the Shandong population, it was recorded as Shandong diversity SNP site. Similar rules were used in the SNP site recording related to sexes. Figure 4 illustrates the total SNP site numbers that we found as Beijing diversity, Shandong diversity, and diversity between sexes in each chemosensory gene type. Interestingly, most of the SNPs in OBPs and ORs were Beijing diversity ones, while fewer showed diversity in the Shandong population and only several SNPs were correlated to the sexes of S. bifasciatus; however, the Beijing diversity and Shandong diversity SNPs number were similar in CSPs, GRs, IRs, and SNMPs.
4. Discussion
Little is known about the olfactory mechanisms of S. bifasciatus, which are very serious cypress pests [3,4]. At the same time, the olfactory response of S. bifasciatus populations from different places to the same attractant are varied (unpublished data). The genetic basis related to physiological differences of insects may be gene expression level differences or sequences variation [48,49,50]. We found that in males of S. bifasciatus, the expression levels of OBPs, ORs, and SNMPs of the Beijing population were obviously lower than those of the Shandong population, and most conserved SNPs of OBPs and ORs from the two populations were the ones that showed more diversity in the Beijing population.
Chemosensory mechanism studies of longhorn beetles have been few, up to now, and chemosensory genes from only a few species have been identified [51]. We identified 18 OBPs, 21 CSPs, 66 ORs, 24 GRs, 14 IRs, and 4 SNMPs in S. bifasciatus, and these numbers were compared to those of Anoplophora glabripennis (42 OBPs, 12 CSPs, 37 ORs, 11 GRs, 4 IRs, and 2 SNMPs), a longhorn beetle whose genome has been sequenced [52]. Thus, our results applied the chemosensory genes of S. bifasciatus relatively completely, and this will facilitate the chemosensory study of S. bifasciatus and longhorn beetles.
Previous trapping variation in S. bifasciatus from two different sites indicated that the olfactory systems of these two populations were different (unpublished data). The reasons could be quantitative or qualitative diversities of their chemosensory genes [48,49,50]. Thus, to explore the quantitative differences in the olfactory systems of these two S. bifasciatus populations, we analyzed the expression levels of S. bifasciatus from the two sites. Interestingly, both of these aspects were different in these two S. bifasciatus populations. Many OBP, OR, and SNMP genes expressions were lower in males of S. bifasciatus from Beijing than those from Shandong. However, the attracting effect of the lure in the Shandong population was worse than in the Beijing population (unpublished data). We can deduce that either the olfactory levels were not the main reason for the attraction differences, or that the expression level was negatively related to the attraction effects of the lure we used.
To analyze the qualitative differences between the olfactory systems of the two S. bifasciatus populations, olfactory gene SNP analysis was performed. For OBPs, ORs, the SNP sites referring to the diversity of the Beijing population were more than that of the Shandong population, which may explain the fact that our lure could attract the Beijing population better, as the diversity of the chemosensory genes may contribute to the recognition of lures [53]. Few studies related to the SNP variations between populations of forest pests have been carried out [53,54], and our work has applied a new sight-to-reason analysis of the population olfactory differences of the same forest pests. However, further analysis of OBP and OR gene functions is necessary to explain the chemosensory recognition diversity of the two S. bifasciatus populations.
5. Conclusions
To explore the olfactory variety of S. bifasciatus, we used next-generation sequencing to analyze the antennal transcriptome of both sexes of the two populations. The olfactory-related genes were identified. Furthermore, expression levels and SNPs of the olfactory-related genes between the two populations were analyzed. We found that in males, the expression levels of many OBP, OR, and SNMP genes of S. bifasciatus from Beijing were obviously lower than those from Shandong, and most conserved SNPs in OBPs and ORs from the two populations showed more diversity in the Beijing population. This work provides a foundation for future research into the molecular olfactory mechanisms of S. bifasciatus, as well as other longhorn beetles, and a basis for the development of new and accurate attractants for the management of this forest pest.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1999-4907/10/8/655/s1, Table S1: Identified chemosensory genes in S. bifasciatus and their unigene ID, Table S2: Collected conserved SNPs in two S. bifasciatus populations, Figure S1: Comparison of chemosensory protein (CSP) genes expression levels in both sexes of the two S. bifasciatus populations. The horizontal axis refers to the expression level of the genes (FPKM values, mean ± SE, n = 3), Figure S2: Comparison of gustatory receptor (GR) genes expression levels in both sexes of the two S. bifasciatus populations. The horizontal axis refers to the expression level of the genes (FPKM values, mean ± SE, n = 3), Figure S3: Comparison of ionotropic receptor (IR) genes expression levels in both sexes of the two S. bifasciatus populations. The vertical axis refers to the expression level of the genes (FPKM values, mean ± SE, n = 3).
Author Contributions
S.Z. (Sufang Zhang) designed the experiments and performed the bioinformatic analysis, as well as drafting the manuscript; S.S. assisted in identification of the chemosensory genes; S.Z. (Shiyu Zhang), X.K., H.W., and F.L. collected the insects.; Z.Z. designed the experiments and modified the manuscript.
Funding
This research was funded by The National Key Research and Development Program of China, grant number 2018YFC1200400 and 2017YFD0600102, The Fundamental Research Funds for the Central Non-profit Research Institution of CAF, grant number CAFYBB2018SZ006 and CAFYBB2017QB003, and The National Nature Science Foundation of China, grant number 31670657.
Acknowledgments
We thank the BioMarker Company for transcriptome sequencing.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gao, S.; Xu, Z.; Gong, X. Progress in research on semanotus bifasciatus. For. Pest Dis. 2007, 26, 19–22. [Google Scholar]
- Wang, S. Quarantine Objects of Forest Plant in China; China Forestry Publishing House: Beijing, China, 1996. [Google Scholar]
- Wu, X.; Wang, J.; Liu, H.; Dong, l.; Jin, Y. Chemical analysis and electroantennogram responses in Semanotus bifasciatus adults to Platycladus orientalis. Chin. Bull. Entomol. 2007, 44, 671–675. [Google Scholar]
- Kong, X.; Zhang, Z.; Wang, H.; Yang, J.; Hu, Y. Analysis of Platycladus orientalis volatiles and their elecctroantennogram responses with Semanotus bifasciatus. For. Res. 2005, 18, 260–266. [Google Scholar]
- Sun, Y. Sensilla and Behavioral Responses of Adults of Semanotus bifasciatus (Coleopetera: Cerambycidae) to Volatile Compounds of Platycladus orientalis; Beijing Forest Universtiy: Beijing, China, 2008. [Google Scholar]
- Su, C.Y.; Menuz, K.; Carlson, J.R. Olfactory perception: Receptors, cells, and circuits. Cell 2009, 139, 45–59. [Google Scholar] [CrossRef]
- Vogt, R.G.; Miller, N.E.; Litvack, R.; Fandino, R.A.; Sparks, J.; Staples, J.; Friedman, R.; Dickens, J.C. The insect SNMP gene family. Insect Biochem. Mol. Biol. 2009, 39, 448–456. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Z.; Wang, H.; Kong, X. Antennal transcriptome analysis and comparison of olfactory genes in two sympatric defoliators, Dendrolimus houi and Dendrolimus kikuchii (lepidoptera: Lasiocampidae). Insect Biochem. Mol. Boil. 2014, 52, 69–81. [Google Scholar] [CrossRef]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef]
- Touhara, K.; Vosshall, L.B. Sensing odorants and pheromones with chemosensory receptors. Annu. Rev. Physiol. 2009, 71, 307–332. [Google Scholar] [CrossRef]
- Kaupp, U.B. Olfactory signaling in vertebrates and insects: Differences and commonalities. Nat. Rev. Neurosci. 2010, 11, 188–200. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Dahanukar, A.; Weiss, L.A.; Carlson, J.R. The molecular basis of CO2 reception in drosophila. Proc. Natl. Acad. Sci. USA 2007, 104, 3574–3578. [Google Scholar] [CrossRef]
- Robertson, H.M.; Kent, L.B. Evolution of the gene lineage encoding the carbon dioxide receptor in insects. J. Insect Sci. 2009, 9, 19. [Google Scholar] [CrossRef]
- Clyne, P.J.; Warr, C.G.; Freeman, M.R.; Lessing, D.; Kim, J.; Carlson, J.R. A novel family of divergent seven-transmembrane proteins: Candidate odorant receptors in Drosophila. Neuron 1999, 22, 327–338. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Amrein, H.; Morozov, P.S.; Rzhetsky, A.; Axel, R. A spatial map of olfactory receptor expression in the Drosophila antenna. Cell 1999, 96, 725–736. [Google Scholar] [CrossRef]
- Hallem, E.A.; Carlson, J.R. Coding of odors by a receptor repertoire. Cell 2006, 125, 143–160. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Hansson, B.S. A unified nomenclature system for the insect olfactory coreceptor. Chem. Senses 2011, 36, 497–498. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Stocker, R.F. Molecular architecture of smell and taste in Drosophila. Annu. Rev. Neurosci. 2007, 30, 505–533. [Google Scholar] [CrossRef]
- Sanchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef]
- Vogt, R.G. Biochemical diversity of odor detection: OBPs, ODEs and SNMPs. In Insect Pheromone Biochemistry and Molecular Biology; Academic Press: London, UK, 2003; pp. 391–446. [Google Scholar]
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef]
- Engsontia, P.; Sanderson, A.P.; Cobb, M.; Walden, K.K.O.; Robertson, H.M.; Brown, S. The red flour beetle’s large nose: An expanded odorant receptor gene family in Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 387–397. [Google Scholar] [CrossRef]
- Krieger, J.; Grosse-Wilde, E.; Gohl, T.; Dewer, Y.M.E.; Raming, K.; Breer, H. Genes encoding candidate pheromone receptors in a moth (Heliothis virescens). Proc. Natl. Acad. Sci. USA 2004, 101, 11845–11850. [Google Scholar] [CrossRef]
- Robertson, H.M.; Wanner, K.W. The chemoreceptor superfamily in the honey bee, Apis mellifera: Expansion of the odorant, but not gustatory, receptor family. Genome Res. 2006, 16, 1395–1403. [Google Scholar] [CrossRef]
- Tanaka, K.; Uda, Y.; Ono, Y.; Nakagawa, T.; Suwa, M.; Yamaoka, R.; Touhara, K. Highly selective tuning of a silkworm olfactory receptor to a key mulberry leaf volatile. Curr. Biol. 2009, 19, 881–890. [Google Scholar] [CrossRef]
- Gong, D.-P.; Zhang, H.-J.; Zhao, P.; Xia, Q.-Y.; Xiang, Z.-H. The odorant binding protein gene family from the genome of silkworm, Bombyx mori. BMC Genom. 2009, 10, 332. [Google Scholar] [CrossRef]
- Zhou, J.-J.; Kan, Y.; Antoniw, J.; Pickett, J.A.; Field, L.M. Genome and est analyses and expression of a gene family with putative functions in insect chemoreception. Chem. Senses 2006, 31, 453–465. [Google Scholar] [CrossRef]
- Zhou, J.J.; He, X.L.; Pickett, J.A.; Field, L.M. Identification of odorant-binding proteins of the yellow fever mosquito Aedes aegypti: Genome annotation and comparative analyses. Insect Mol. Biol. 2008, 17, 147–163. [Google Scholar] [CrossRef]
- Bengtsson, J.M.; Trona, F.; Montagné, N.; Anfora, G.; Ignell, R.; Witzgall, P.; Jacquin-Joly, E. Putative chemosensory receptors of the codling moth, Cydia pomonella, identified by antennal transcriptome analysis. PLoS ONE 2012, 7, e31620. [Google Scholar] [CrossRef]
- Grosse-Wilde, E.; Kuebler, L.S.; Bucks, S.; Vogel, H.; Wicher, D.; Hansson, B.S. Antennal transcriptome of Manduca sexta. Proc. Natl. Acad. Sci. USA 2011, 108, 7449–7454. [Google Scholar] [CrossRef]
- Legeai, F.; Malpel, S.; Montagne, N.; Monsempes, C.; Cousserans, F.; Merlin, C.; Francois, M.-C.; Maibeche-Coisne, M.; Gavory, F.; Poulain, J.; et al. An expressed sequence tag collection from the male antennae of the noctuid moth Spodoptera littoralis: A resource for olfactory and pheromone detection research. BMC Genom. 2011, 12, 86. [Google Scholar] [CrossRef]
- Zhang, Y.-N.; Jin, J.-Y.; Jin, R.; Xia, Y.-H.; Zhou, J.-J.; Deng, J.-Y.; Dong, S.-L. Differential expression patterns in chemosensory and non-chemosensory tissues of putative chemosensory genes identified by transcriptome analysis of insect pest the purple stem borer Sesamia inferens (walker). PLoS ONE 2013, 8, e69715. [Google Scholar]
- Mitchell, R.F.; Hughes, D.T.; Luetje, C.W.; Millar, J.G.; Soriano-Agatmn, F.; Hanks, L.M.; Robertson, H.M. Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect Biochem. Mol. Biol. 2012, 42, 499–505. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.; Grosse-Wilde, E.; Keeling, C.; Bengtsson, J.; Yuen, M.; Li, M.; Hillbur, Y.; Bohlmann, J.; Hansson, B.; Schlyter, F. Antennal transcriptome analysis of the chemosensory gene families in the tree killing bark beetles, Ips typographus and Dendroctonus ponderosae (coleoptera: Curculionidae: Scolytinae). BMC Genom. 2013, 14, 1–16. [Google Scholar] [CrossRef]
- Zhang, S.-F.; Liu, H.-H.; Kong, X.-B.; Wang, H.-B.; Liu, F.; Zhang, Z. Identification and expression profiling of chemosensory genes in Dendrolimus punctatus walker. Front. Physiol. 2017, 8, 8. [Google Scholar] [CrossRef]
- Bogdanova, E.A.; Shagin, D.A.; Lukyanov, S.A. Normalization of full-length enriched cDNA. Mol. BioSystems 2008, 4, 205–212. [Google Scholar] [CrossRef]
- Zhulidov, P.A.; Bogdanova, E.A.; Shcheglov, A.S.; Vagner, L.L.; Khaspekov, G.L.; Kozhemyako, V.B.; Matz, M.V.; Meleshkevitch, E.; Moroz, L.L.; Lukyanov, S.A.; et al. Simple cDNA normalization using kamchatka crab duplex-specific nuclease. Nucleic Acids Res. 2004, 32, e37. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. Tigr gene indices clustering tools (tgicl): A software system for fast clustering of largest datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2go: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the blast2go suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential Expression of RNA-seq Data at the Gene Level—The Deseq Package; European Molecular Biology Laboratory (EMBL): Heidelberg, Germany, 2012. [Google Scholar]
- Bullard, J.H.; Purdom, E.; Hansen, K.D.; Dudoit, S. Evaluation of statistical methods for normalization and differential expression in mRNA-seq experiments. BMC Bioinform. 2010, 11, 94. [Google Scholar] [CrossRef]
- Noble, W.S. How does multiple testing correction work? Nat. Biotechnol. 2009, 27, 1135–1137. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A mapreduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- He, Y.Q.; Feng, B.; Guo, Q.S.; Du, Y. Age influences the olfactory profiles of the migratory oriental armyworm mythimna separate at the molecular level. BMC Genom. 2017, 18, 32. [Google Scholar] [CrossRef]
- Qiu, C.Z.; Zhou, Q.Z.; Liu, T.-T.; Fang, S.M.; Wang, Y.W.; Fang, X.; Huang, C.L.; Yu, Q.Y.; Chen, C.H.; Zhang, Z. Evidence of peripheral olfactory impairment in the domestic silkworms: Insight from the comparative transcriptome and population genetics. BMC Genom. 2018, 19, 788. [Google Scholar] [CrossRef]
- Gadenne, C.; Barrozo, R.B.; Anton, S. Plasticity in insect olfaction: To smell or not to smell? Annu. Rev. Èntomol. 2016, 61, 317–333. [Google Scholar] [CrossRef]
- Zhan, W.; Zhang, S.; Geng, H.; Wang, Y.; Guo, K.; Chen, J. Research progress on olfactory recognition proteins of longhorn beetle. J. Henan Agric. Sci. 2018, 47, 1–6. [Google Scholar]
- McKenna, D.D.; Scully, E.D.; Pauchet, Y.; Hoover, K.; Kirsch, R.; Geib, S.M.; Mitchell, R.F.; Waterhouse, R.M.; Ahn, S.-J.; Arsala, D.; et al. Genome of the Asian longhorned beetle (Anoplophora glabripennis), a globally significant invasive species, reveals key functional and evolutionary innovations at the beetle–plant interface. Genome Boil. 2016, 17, 227. [Google Scholar] [CrossRef]
- Wang, P.; Lyman, R.F.; Mackay, T.F.C.; Anholt, R.R.H. Natural variation in odorant recognition among odorant-binding proteins in drosophila melanogaster. Genetics 2010, 184, 759–767. [Google Scholar] [CrossRef]
- Wang, P.; Lyman, R.F.; Shabalina, S.A.; Mackay, T.F.C.; Anholt, R.R.H. Association of polymorphisms in odorant-binding protein genes with variation in olfactory response to benzaldehyde in drosophila. Genetics 2007, 177, 1655–1665. [Google Scholar] [CrossRef]
Figure 1.
Comparison of odorant binding protein (OBP) genes expression levels in both sexes of the two S. bifasciatus populations. The horizontal axis refers to the expression level of the genes (FPKM values, mean ± SE, n = 3). (A): OBPs expression level comparison between females of the two population; (B): OBPs expression level comparison between males of the two population. BJ, Beijing population; SD, Shandong population. * means the difference is significant at the 0.05 level; ** means the difference is significant at the 0.01 level.
Figure 1.
Comparison of odorant binding protein (OBP) genes expression levels in both sexes of the two S. bifasciatus populations. The horizontal axis refers to the expression level of the genes (FPKM values, mean ± SE, n = 3). (A): OBPs expression level comparison between females of the two population; (B): OBPs expression level comparison between males of the two population. BJ, Beijing population; SD, Shandong population. * means the difference is significant at the 0.05 level; ** means the difference is significant at the 0.01 level.

Figure 2.
Comparison of odorant receptor (OR) genes expression levels in both sexes of the two S. bifasciatus populations. The horizontal axis refers to the expression level of the genes (FPKM values, mean ± SE, n = 3). (A): ORs expression level comparison between females of the two population; (B): ORs expression level comparison between males of the two population. BJ, Beijing population; SD, Shandong population. * means the difference is significant at the 0.05 level; ** means the difference is significant at the 0.01 level.
Figure 2.
Comparison of odorant receptor (OR) genes expression levels in both sexes of the two S. bifasciatus populations. The horizontal axis refers to the expression level of the genes (FPKM values, mean ± SE, n = 3). (A): ORs expression level comparison between females of the two population; (B): ORs expression level comparison between males of the two population. BJ, Beijing population; SD, Shandong population. * means the difference is significant at the 0.05 level; ** means the difference is significant at the 0.01 level.

Figure 3.
Comparison of sensory neuron membrane protein (SNMP) genes expression levels in both sexes of the two S. bifasciatus populations. The vertical axis refers to the expression level of the genes (FPKM values, mean ± SE, n = 3). (A): SNMPs expression level comparison between females of the two population; (B): SNMOs expression level comparison between males of the two population. BJ, Beijing population; SD, Shandong population. * means the difference is significant at the 0.05 level; ** means the difference is significant at the 0.01 level.
Figure 3.
Comparison of sensory neuron membrane protein (SNMP) genes expression levels in both sexes of the two S. bifasciatus populations. The vertical axis refers to the expression level of the genes (FPKM values, mean ± SE, n = 3). (A): SNMPs expression level comparison between females of the two population; (B): SNMOs expression level comparison between males of the two population. BJ, Beijing population; SD, Shandong population. * means the difference is significant at the 0.05 level; ** means the difference is significant at the 0.01 level.

Figure 4.
Three categories of collected conserved SNPs. BJ, Beijing; SD, Shandong.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, S.; Shen, S.; Zhang, S.; Wang, H.; Kong, X.; Liu, F.; Zhang, Z. Chemosensory Characteristics of Two Semanotus bifasciatus Populations. Forests 2019, 10, 655. https://0-doi-org.brum.beds.ac.uk/10.3390/f10080655
AMA Style
Zhang S, Shen S, Zhang S, Wang H, Kong X, Liu F, Zhang Z. Chemosensory Characteristics of Two Semanotus bifasciatus Populations. Forests. 2019; 10(8):655. https://0-doi-org.brum.beds.ac.uk/10.3390/f10080655
Chicago/Turabian StyleZhang, Sufang, Sifan Shen, Shiyu Zhang, Hongbin Wang, Xiangbo Kong, Fu Liu, and Zhen Zhang. 2019. "Chemosensory Characteristics of Two Semanotus bifasciatus Populations" Forests 10, no. 8: 655. https://0-doi-org.brum.beds.ac.uk/10.3390/f10080655
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.