Changes in Major Insect Pests of Pine Forests in Korea Over the Last 50 Years


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
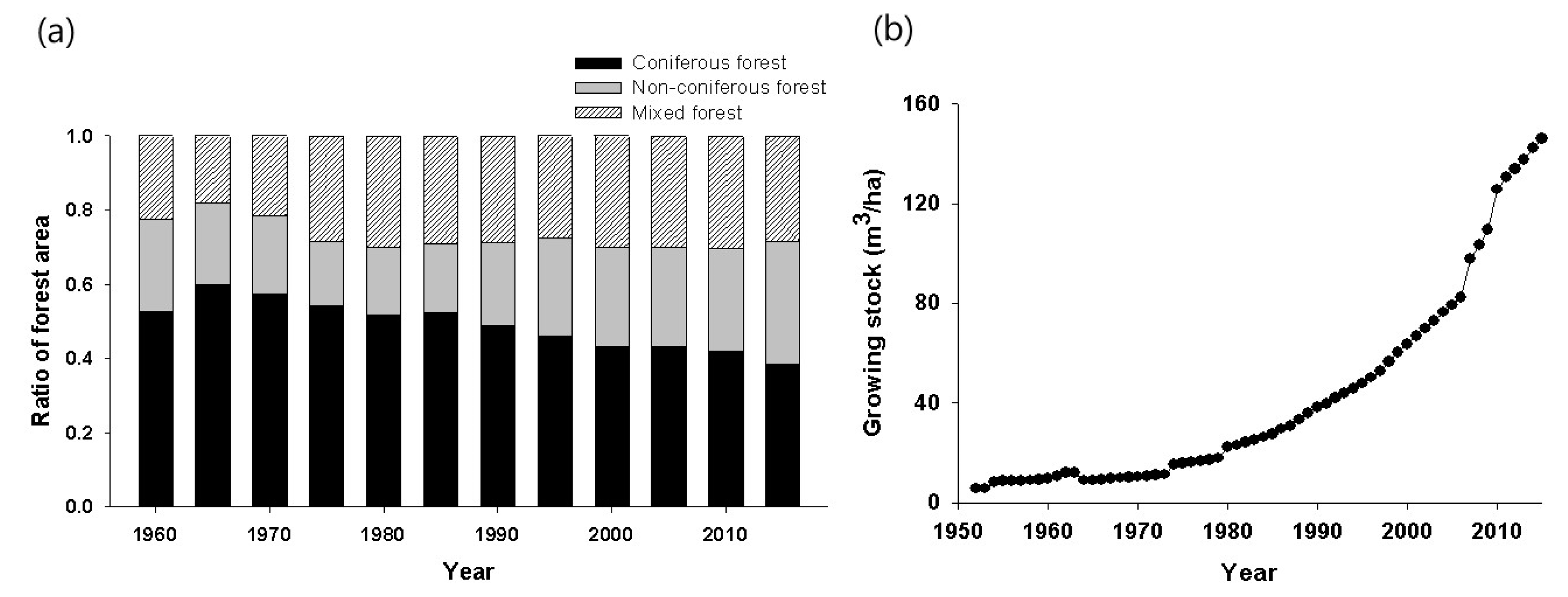
2. Forest Change and Monitoring Records in Korea
3. Monitoring of Korean Forest Pests
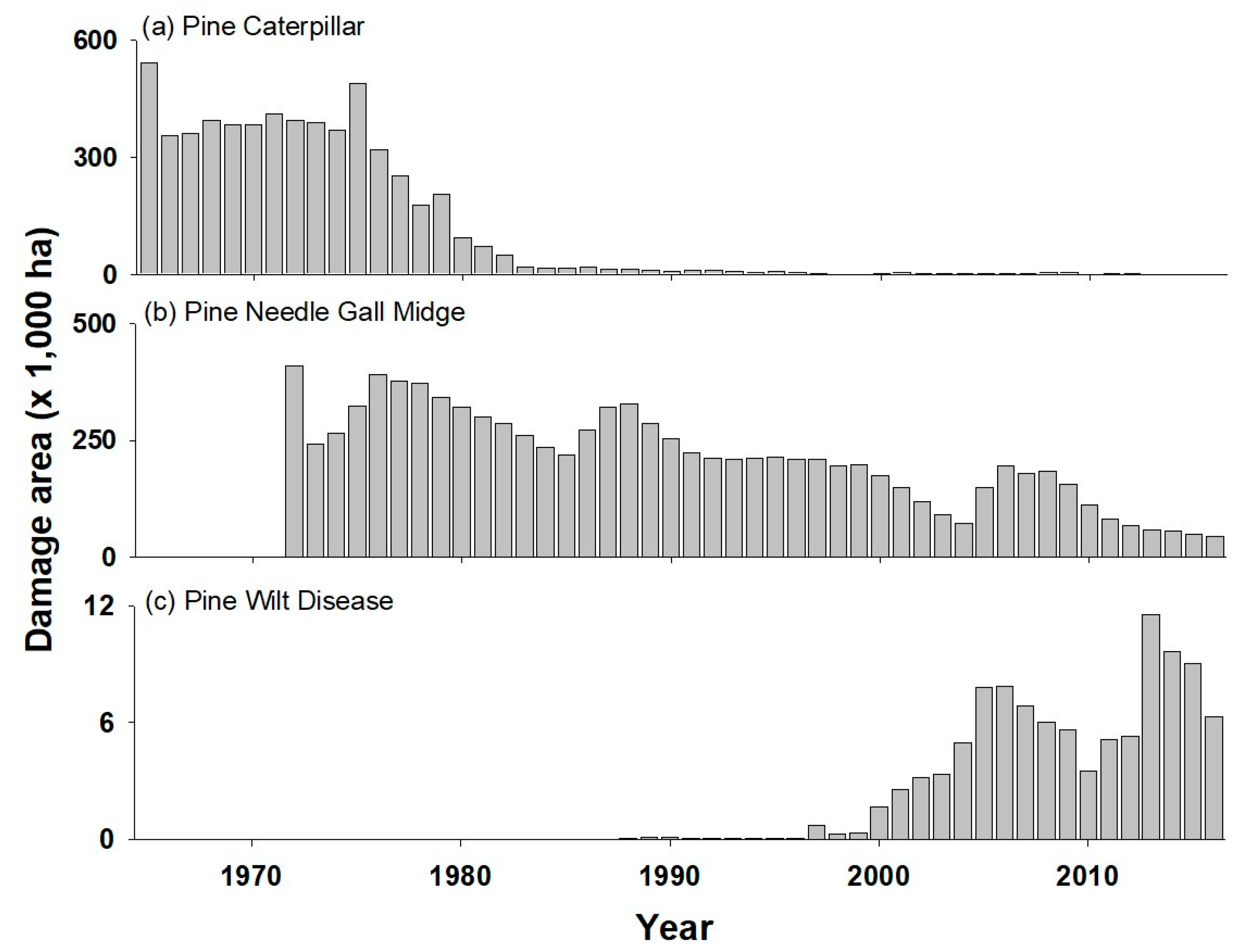
4. Changes in Major Pests in Pine Forests
4.1. Pine Caterpillar (PC)
4.1.1. Occurrence History and Ecology
4.1.2. Environmental Factors and Management
4.2. Pine Needle Gall Midge (PNGM)
4.2.1. Occurrence History and Ecology
4.2.2. Environmental Factors and Management
4.3. Pine Wilt Disease (PWD)
4.3.1. Occurrence History and Ecology
4.3.2. Environmental Factors and Management
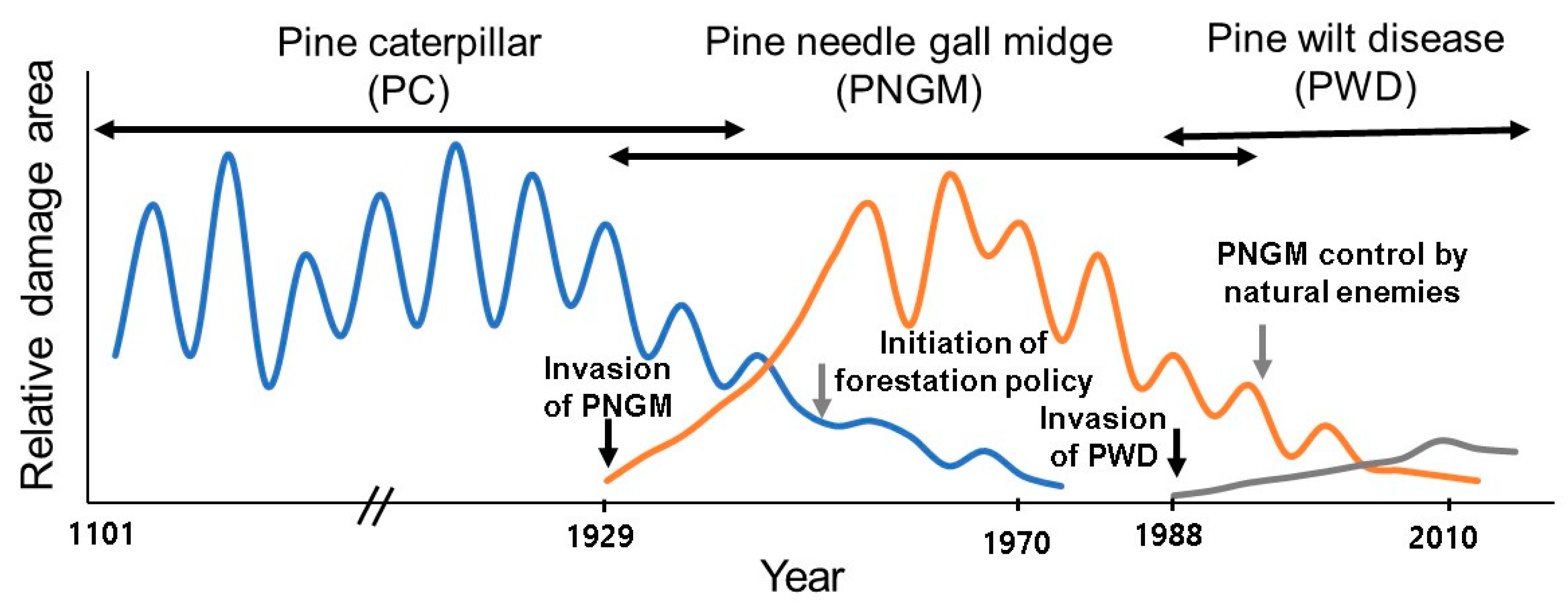
5. Causes of Changes in Dominance of Insect Pests in Korean Forests
6. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walter, J.A.; Ives, A.R.; Tooker, J.F.; Johnson, D.M. Life history and habitat explain variation among insect pest population subject to global change. Ecosphere 2018, 9, e02274. [Google Scholar] [CrossRef]
- Hentschel, R.; Möller, K.; Wenning, A.; Degenhardt, A.; Schröder, J. Importance of ecological variables in explaining population dynamics of three important pine pest insects. Front. Plant Sci. 2018, 9, 1667. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.J.; Bauer, L.S.; Abell, K.J.; Ulyshen, M.D.; Van Driesche, R.G. Population dynamics of an invasive forest insect and associated natural enemies in the aftermath of invasion: Implications for biological control. J. Appl. Ecol. 2015, 52, 1246–1254. [Google Scholar] [CrossRef]
- Guyot, V.; Castagneyrol, B.; Vialatte, A.; Deconchat, M.; Jactel, H. Tree diversity reduces pest damage in mature forest across Europe. Biol. Lett. 2018, 12, 20151037. [Google Scholar] [CrossRef] [PubMed]
- Speer, J.H.; Swetman, T.W.; Wickman, B.E.; Youngblood, A. Changes in pandora moth outbreak dynamics during the past 622 years. Ecology 2001, 82, 679–697. [Google Scholar] [CrossRef]
- Möller, K.; Hentschel, R.; Wenning, A.; Schröder, J. Improved outbreak prediction for common pine sawfly (Diprion pini L.) by analyzing floating ‘climatic windows’ as keys for changes in voltinism. Forests 2017, 8, 319. [Google Scholar] [CrossRef]
- Li, S.; Daudin, J.J.; Piou, D.; Robinet, C.; Jactel, H. Periodicity and synchrony of pine processionary moth outbreaks in France. For. Ecol. Manag. 2015, 354, 309–317. [Google Scholar] [CrossRef]
- Schwerdtfeger, F. Ueber die Ursachen des Massenwechsels der Insekten. Ztg. Angeweine Entomol. 1941, 28, 254–303. [Google Scholar] [CrossRef]
- Turchin, P.; Taylor, A.D. Complex dynamics in ecological time series. Ecology 1992, 73, 289–305. [Google Scholar] [CrossRef]
- Tenow, O. The ourbreaks of Oporina autumnata Bkh. and Operophtera spp. (Lep. Geometridae) in the Scandinavian mountain chain and northern Finland 1862–1968. Zool. Bidr. Upps. Suppl. 1972, 2, 1–107. [Google Scholar]
- Tenow, O.; Nilssen, A.C.; Bylund, H.; Hogstad, O. Waves and synchrony in Epirrita autumnata / Operophtera brumata outbreaks. I. Lagged synchrony: Regionally, locally and among species. J. Anim. Ecol. 2007, 76, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.I.; Park, Y.-S. Dispersal patterns of exotic forest pests in South Korea. Insect Sci. 2012, 19, 535–548. [Google Scholar] [CrossRef]
- Korea Forest Research Institute (KFRI). Annual Report of Monitoring for Forest Insect Pests and Diseases in Korea; SeongMunSa: Seoul, Korea, 1968–2018. [Google Scholar]
- Korea Forest Service. Statistical Yearbook of Forestry; Korea Forest Service: Daejeon, Korea, 2018. [Google Scholar]
- Park, W.K.; Lee, K.-H. Changes in the species of woods used for Korean ancient and historic architectures. J. Archit. Hist. 2007, 16, 9–28. [Google Scholar]
- Moon, M.; Kim, S.-S.; Lee, D.-S.; Yang, H.; Park, C.-W.; Kim, H.; Park, Y.-S. Effects of forest management practices on moth communities in a Japanese larch (Larix kaempferi (Lamb.) Carrière) plantation. Forests 2018, 9, 574. [Google Scholar] [CrossRef]
- Bae, J.-S.; Joo, R.-W.; Lee, K.-B. Causes of Forest Degradation and Drivers of Forest Recovery in South Korea; Upgo MunHwa: Seoul, Korea, 2010. [Google Scholar]
- Bae, J.-S.; Lee, K.-B. Impacts of the substitution of firewood for home use on the forest greening after the 1945 liberation of Korea. J. Korean For. Soc. 2006, 95, 60–72. [Google Scholar]
- Lee, K.-B.; Bae, J.-S. Factors of success of the clearance policy for slash-and-burn fields in the 1970s. J. Korean For. Soc. 2007, 96, 325–337. [Google Scholar]
- Lee, D.-S.; Bae, Y.-S.; Byun, B.-K.; Lee, S.; Park, J.K.; Park, Y.-S. Occurrence prediction of the citrus flatid planthopper (Metcalfa pruinosa (Say, 1830)) in South Korea using a random forest model. Forest 2019, 10, 583. [Google Scholar] [CrossRef]
- Park, K.N.; Hyun, J.S. Studies on the effects of the pine needle gall midge, Thecodiplosis japonensis Uchida et Inouye, on the growth of the red pine, Pinus densiflora S. et Z. (II)—Growth impact on red pine. J. Korean For. Soc. 1983, 62, 87–95. [Google Scholar]
- Park, Y.-S.; Chung, Y.-J.; Moon, Y.-S. Hazard ratings of pine forests to a pine wilt disease at two spatial scales (individual trees and stands) using self-organizing map and random forest. Ecol. Inform. 2013, 13, 40–46. [Google Scholar] [CrossRef]
- Baek, S.L. The outbreak of pine caterpillar and colonial authorities’ response. Han’guk Munhwa Korean Cult. 2018, 81, 93–123. [Google Scholar]
- Choi, W.I.; Park, Y.K.; Park, Y.-S.; Lee, H.P.; Ryoo, M.I. Changes in voltinism in a pine caterpillar Dendrolimus spectabilis (Lepidoptera: Lasiocampidae) population: Implications of climatic change. Appl. Entomol. Zool. 2011, 46, 319–325. [Google Scholar] [CrossRef]
- Chung, K.H.; Ryu, J.; Kwon, S.H.; Im, M.S. Field studies on the attractiveness of pine caterpillar moths (Dendrolimus spectabilis Butler) to blacklight-traps. Korean J. Plant Prot. 1971, 10, 43–48. [Google Scholar]
- Kim, C.W.; Hyun, J.S. Studies on the control of pine moth, Dendrolimus spectabilis (Butler). Ent. Res. Bull. 1965, 1, 1–109. [Google Scholar]
- Shin, S.C.; Choi, K.S.; Choi, W.I.; Chung, Y.J.; Lee, S.G.; Kim, C.S. An New Illustrated Book of Forest Insect Pests; Upgo MunHwa: Seoul, Korea, 2008. [Google Scholar]
- Kwon, T.-S.; Park, Y.-K.; Oh, K.-S.; Kwon, Y.-D.; Shin, S.-C.; Kim, C.S.; Park, J.D.; Lee, H.-P. Increase in the number of generations in Dendrolimus spectabilis (Butler) (Lepidoptera: Lasiocampidae) in Korea. J. Korean For. Soc. 2002, 91, 149–155. [Google Scholar]
- Park, Y.G.; Kim, C.S. Studies on selecting taste of the pine-caterpillar larva: Dendrolimus spectabilis Butler on six pine species. Korean J. Breed. 1973, 5, 27–31. [Google Scholar]
- Hyun, J.S. Studies on the prevision for occurrence of pine moth, Dendrolimus spectabilis Butler. Ent. Res. Bull. 1968, 4, 57–80. [Google Scholar]
- Kokubo, A. Population fluctuations and natural mortalities of the pine-moth, Dendrolimus spectalbilis. Res. Popul. Ecol. 1965, 7, 23–34. [Google Scholar] [CrossRef]
- Bao, Y.; Wang, F.; Tong, S.; Na, L.; Han, A.; Zhang, J.; Bao, Y.; Han, Y.; Zhang, Q. Effect of drought on outbreaks of major forest pests, Pine Caterpillars (Dendrolimus spp.), in Shandong Province, China. Forests 2019, 10, 264. [Google Scholar] [CrossRef]
- Park, Y.-K. Changes in Population Biology of Pine Moth, Dendrolimus spectabilis (Butler) (Lepidoptera: Lasiocampidae) and Its Biological Control by Trichogramma dendrolimi Matsumura (Hymenoptera: Trichogrammatidae). Ph.D. Dissertation, Dongguk University, Seoul, Korea, 2001. [Google Scholar]
- Aoki, J. Beauveria bassiana (Bals.) Vuill. isolated from some lepidopterous species in Japan. Jpn. J. Appl. Entomol. Zool. 1971, 15, 222–227. [Google Scholar] [CrossRef]
- Pasquarella, V.A.; Elkinton, J.S.; Bradley, B.A. Extensive gypsy moth defoliation in Southern New England characterized using Landsat satellite observations. Biol. Invasions 2018, 20, 3047–3053. [Google Scholar] [CrossRef]
- Moreira, X.; Abdala-Roberts, L.; Rasmann, S.; Castagneyrol, B.; Mooney, K.A. Plant diversity effects on insect herbivores and their natural enemies: Current thinking, recent findings, and future directions. Curr. Opin. Insect Sci. 2016, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Le Borgne, H.; Hebert, C.; Dupuch, A.; Bichet, O.; Pinaud, D.; Fortin, D. Temporal dynamics in animal community assembly during post-logging succession in boreal forest. PLoS ONE 2018, 13, e0204445. [Google Scholar] [CrossRef] [PubMed]
- Soné, K.; Furuno, T. Annual changes in the infestation by the pine needle gall midge, Thecodiplosis japonensis Uchida et Inouye (Diptera: Cecidomyiidae), in a pine stand. J. Jpn. For. Soc. 1982, 64, 301–306. [Google Scholar]
- Park, K.-N.; Miura, T.; Hirashima, Y. Outbreaks history and present status of the pine needle gall midge in Korea. Esakia 1985, 23, 115–118. [Google Scholar]
- Lee, B.Y.; Chung, Y.J.; Park, K.N.; Byun, B.H.; Bae, W.I. Distribution of pine needle gall midge, Thecodiplosis japonensis Uchida et Inouye (Diptera: Cecidomyiidae), infestations in Korea: A brief history. FRI J. For. Sci. 1997, 56, 13–20. [Google Scholar]
- Shigesada, N.; Kawasaki, K.; Takeda, Y. Modeling stratified diffusion in biological invasions. Am. Nat. 1995, 146, 229–251. [Google Scholar] [CrossRef]
- Park, Y.-S.; Chung, Y.-J. Hazard rating of pine trees from a forest insect pest using artificial neural networks. For. Ecol. Manag. 2006, 222, 222–233. [Google Scholar] [CrossRef]
- Park, K.N.; Hyun, J.S. Studies on the effects of the pine needle gall midge, Thecodiplosis japonensis Uchida et Inouye, on the growth of the red pine, Pinus densiflora Siebold et Zuccarini (I): Changes of gall formation rate. J. Korean For. Soc. 1983, 61, 20–26. [Google Scholar]
- Chung, Y.-J.; Park, Y.-S.; Lee, B.-Y.; Chon, T.-S. Dynamic patterns of the infestation of pine needle gall midge, Thecodiplosis japonensis (Diptera: Cecidomyiidae), in endemic regions of Korea. FRI J. For. Sci. 1998, 59, 64–69. [Google Scholar]
- Ko, J.H. Studies on the Isostasius seoulis Ko, the Larval Parasite of the Pine Gall-Midge (Thecodiplosis japonensis Uchida et Inouye). III: The Life History; Research Report; Rural Development: Suwon, Korea, 1966. [Google Scholar]
- Choi, W.I.; Song, H.J.; Kim, D.S.; Lee, D.-S.; Lee, C.-Y.; Nam, Y.; Kim, J.-B.; Park, Y.-S. Dispersal patterns of pine wilt disease in the early stage of its invasion in South Korea. Forests 2017, 8, 411. [Google Scholar] [CrossRef]
- Nam, Y.; Choi, W.I. An empirical predictive model for the spring emergence of Thecodiplosis japonensis (Diptera: Cecidomyiidae): Model construction and validation on the basis of 25 years field observations data. J. Econ. Entomol. 2014, 107, 1136–1141. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.J.; Hyun, J.-S. Studies on the major factors affecting the population of the overwintered pine needle gall midge, Thecodiplosis japonensis Uchida et Inouye. Korean J. Plant Prot. 1986, 25, 1–9. [Google Scholar]
- Ryoo, M.I.; Chun, Y.S. Population ecology of pine gall midge (Thecodiplosia japonensis): Revisited. Nat. Resour. Res. 1996, 4, 118–129. [Google Scholar]
- Choi, W.I.; Ryoo, M.I.; Chung, Y.J.; Park, Y.-S. Geographical variation in the population dynamics of Thecodiplosis japonensis: Causes and effects on spatial synchrony. Popul. Ecol. 2011, 53, 429–439. [Google Scholar] [CrossRef]
- Choi, W.I.; Jeon, M.J.; Park, Y.S. Structural dynamics in the host-parasitoid system of the pine needle gall midge (Thecodiplosis japonensis) during invasion. PeerJ 2017, 5, e3610. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Chung, Y.-J.; Chon, T.-S.; Lee, B.-Y.; Lee, J.-H. Interactions between pine deedle gall midge, Thecodiplosis japonensis (Diptera: Cecidomyiidae), and its parasitoids in newly invaded areas. Korean J. Appl. Entomol. 2001, 40, 301–307. [Google Scholar]
- Jeon, M.-J.; Choi, W.I.; Choi, K.-S.; Chung, Y.-J.; Shin, S.-C. Population dynamics of Thecodiplosis jsponenste (Dlptera: Cecidomyiidae) under influence of parasitism by Inostemma matsutama and Inostemma seoulis (Hymenoptera: Platygastridae). J. Asia-Pac. Entomol. 2006, 9, 269–274. [Google Scholar] [CrossRef]
- Yoshida, N.; Hirashima, Y. Systematic studies on proctotrupoid and chalcidoid parasites of gall midges injurious to Pinus and Cryptomeria in Japan and Korea (Hymenoptera). Esakia 1979, 14, 113–133. [Google Scholar]
- Ko, J.H. A new species of Inostemma (Hymenoptera: Platygasteridae), a larval parasite of the pine gall midge, Thecodiplosis sp. (Diptera: Cecidomyoodae). Korean J. Plant Prot. 1980, 19, 35–38. [Google Scholar]
- Jeon, M.-J.; Lee, B.-Y.; Ko, J.-H.; Miura, T.; Hirashima, Y. Ecology of Platygaster matsutama and Inostemma seoulis (Hymenoptera: Platygastridae), egg-larval parasites of the pine needle gall midge, Thecodiplosis japonensis (Diptera, Cecidomyiidae). Esakia 1985, 23, 131–143. [Google Scholar]
- Soné, K. Ecology of host-parasitoid community in the pine needle gall midge, Thecodiplosis japonensis Uchida et Inouye (Diptera, Cecidomyiidae). J. Appl. Entomol. 1986, 102, 516–527. [Google Scholar] [CrossRef]
- Son, Y.; Chung, Y.-J.; Lee, J.-H. Differential thermal biology may explain the coexistence of Platygaster matsutama and Inostemma seoulis (Hymenoptera: Platygastridae) attacking Thecodiplosis japonensis (Diptera: Cecidomyiidae). J. Asia-Pac. Entomol. 2012, 15, 465–471. [Google Scholar] [CrossRef]
- Lee, D.-S.; Nam, Y.; Choi, W.I.; Park, Y.-S. Environmental factors influencing on the occurrence of pine wilt disease in Korea. Korean J. Ecol. Environ. 2017, 50, 374–380. [Google Scholar] [CrossRef]
- Togashi, K.; Shigesada, N. Spread of the pinewood nematode vectored by the Japanese pine sawyer: Modeling and analytical approaches. Popul. Ecol. 2006, 48, 271–283. [Google Scholar] [CrossRef]
- Mota, M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar]
- Kiritani, K.; Morimoto, N. Invasive insect and nematode pests from north America. Glob. Environ. Res. 2004, 8, 75–88. [Google Scholar]
- Shin, S.C. Pine Wilt Disease in Korea. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008. [Google Scholar]
- Lee, S.D.; Park, S.; Park, Y.-S.; Chung, Y.-J.; Lee, B.-Y.; Chon, T.-S. Range expansion of forest pest populations by using the lattice model. Ecol. Model. 2007, 203, 157–166. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Park, Y.-S.; Jeoung, C.-S.; Choi, W.-I.; Kim, Y.-K.; Jung, I.-H.; Shigesada, N.; Kawasaki, K.; Takasu, F.; Chon, T.-S. Spatially explicit model applied to pine wilt disease dispersal based on host plant infestation. Ecol. Model. 2017, 353, 54–62. [Google Scholar] [CrossRef]
- Robinet, C.; Roques, A.; Pan, H.; Fang, G.; Ye, J.; Zhang, Y.; Sun, J. Role of human-mediated dispersal in the spread of the pinewood nematode in China. PLoS ONE 2009, 4, e4646. [Google Scholar] [CrossRef]
- Hirata, A.; Nakamura, K.; Nakao, K.; Kominami, Y.; Tanaka, N.; Ohashi, H.; Takano, K.T.; Takeuchi, W.; Matsui, T. Potential distribution of pine wilt disease under future climate change scenarios. PLoS ONE 2017, 12, e0182837. [Google Scholar] [CrossRef]
- Kwon, T.S.; Lim, J.H.; Sim, S.J.; Kwon, Y.D.; Son, S.K.; Lee, K.Y.; Kim, Y.T.; Park, J.W.; Shin, C.H.; Ryu, S.B.; et al. Distribution patterns of Monochamus alternatus and M. saltuarius (Coleoptera: Cerambycidae) in Korea. J. Korean For. Soc. 2006, 95, 543–550. [Google Scholar]
- Togashi, K. Vector-Nematode relationships and epidemiology in pine wilt disease. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 162–183. [Google Scholar]
- Kwon, H.J.; Jung, J.-K.; Jung, C.; Han, H.; Koh, S.-H. Dispersal capacity of Monochamus saltuarius on flight mills. Entomol. Exp. Appl. 2018, 166, 420–427. [Google Scholar] [CrossRef]
- Hu, S.J.; Ning, T.; Fu, D.Y.; Haack, R.A.; Zhang, Z.; Chen, D.D.; Ma, X.Y.; Ye, H. Dispersal of the japanese pine sawyer, Monochamus alternatus (Coleoptera: Cerambycidae), in mainland China as inferred from molecular data and associations to indices of human activity. PLoS ONE 2013, 8, e57568. [Google Scholar] [CrossRef] [PubMed]
- Kawai, M.; Shoda-Kagaya, E.; Maehara, T.; Zhou, Z.; Lian, C.; Iwata, R.; Yamane, A.; Hogetsu, T. Genetic structure of pine sawyer Monochamus alternatus (Coleoptera: Cerambycidae) populations in Northeast Asia: Consequences of the spread of pine wilt disease. Environ. Entomol. 2006, 35, 569–579. [Google Scholar] [CrossRef]
- Nakamura-Matori, K. Vector-host tree relationships and the abiotic environment. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 144–161. [Google Scholar]
- Jikumaru, S.; Togashi, K. Effects of temperature on the post-diapause development of Monochamus saltuarius (Gebler) (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1996, 31, 145–148. [Google Scholar] [CrossRef]
- Jung, C.S.; Koh, S.-H.; Nam, Y.; Ahn, J.J.; Choi, W.I. A forecasting model for predicting the spring emergence of Monochamus saltuarius (Coleoptera: Cerambycidae) on Korean white pine, Pinus koraiensis. J. Econ. Entomol. 2015, 108, 1830–1836. [Google Scholar] [CrossRef] [PubMed]
- Park, C.G.; Kim, D.S.; Lee, S.M.; Moon, Y.S.; Chung, Y.J.; Kim, D.-S. A forecasting model for the adult emergence of overwintered Monochamus alternatus (Coleoptera: Cerambycidae) larvae based on degree-days in Korea. Appl. Entomol. Zool. 2014, 49, 35–42. [Google Scholar] [CrossRef]
- Jikumaru, S.; Togashi, K. Temperature effects on the transmission of Bursaphelenchus xylophilus (Nemata: Aphelenchoididae) by Monochamus alternatus (Coleoptera: Cerambycidae). J. Nematol. 2000, 32, 110–116. [Google Scholar]
- Taketani, A.; Okuda, M.; Hosoda, R. The meteorological analysis on the epidemic mortality of pine trees, with special reference to the effective accumulated temperature. J. Jpn. For. Soc. 1975, 57, 169–175. [Google Scholar]
- Korea Forest Research Institute (KFRI). System Design and Structure Analyses for Pine Wilt Disease Control; KFRI: Seoul, Korea, 2010. [Google Scholar]
- Futai, K. Role of asymptomatic carrier trees in epidemic spread of pine wilt disease. J. For. Res. 2003, 8, 253–260. [Google Scholar] [CrossRef]
- Hong, J.-I.; Koh, S.-H.; Chung, Y.J.; Shin, S.S.; Kim, G.-H.; Choi, K.-S. Biological characteristics of Sclerodermus harmandi (Hymenoptera: Bethylidae) parasitized on cerambycid. Korean J. Appl. Entomol. 2008, 47, 133–139. [Google Scholar] [CrossRef]
- Kim, J.K.; Won, D.-S.; Park, Y.C.; Koh, S.-H. Natural enemies of wood borers and seasonal occurrence of major natural enemies of Monochamus slatuarius on pine trees. J. Korean For. Soc. 2010, 99, 439–445. [Google Scholar]
- Zhang, Y.; Yang, Z. Studies on the natural enemies and biocontrol of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Plant Prot. 2006, 32, 9–14. [Google Scholar]
- Shimazu, M. Potential of the Cerambycid-Parasitic Type of Beauveria brongniartii (Deuterpmycotina: Hyphomycetes) for microbial control of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1994, 29, 127–130. [Google Scholar] [CrossRef]
- Petersen-Silva, R.; Pujade-Villar, J.; Naves, P.; Sousa, E.; Belokobylskij, S. Parasitoids on Monochamus galloprovincialis (Coleoptera, Cerambycidae), vector of the pine wood nematode, with identification key for Palearctic region. ZooKeys 2012, 251, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Naves, P.; Kenis, M.; Sousa, E. Parasitoids associated with Monochamus galloprovincialis (Oliv.) (Coleoptera: Cerambycidae) within the pine wilt nematode-affected zone in Portugal. J. Pest Sci. 2005, 78, 57–62. [Google Scholar] [CrossRef]
- Kwon, T.-S.; Shin, J.H.; Lim, J.-H.; Kim, Y.-K.; Lee, E.J. Management of pine wilt disease in Korea through preventative silvicultural control. For. Ecol. Manag. 2011, 261, 562–569. [Google Scholar] [CrossRef]
- Kamata, N. Outbreaks of forest defoliating insects in Japan, 1950–2000. Bull. Entomol. Res. 2002, 92, 109–117. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, W.I.; Nam, Y.; Lee, C.Y.; Choi, B.K.; Shin, Y.J.; Lim, J.-H.; Koh, S.-H.; Park, Y.-S. Changes in Major Insect Pests of Pine Forests in Korea Over the Last 50 Years. Forests 2019, 10, 692. https://0-doi-org.brum.beds.ac.uk/10.3390/f10080692
Choi WI, Nam Y, Lee CY, Choi BK, Shin YJ, Lim J-H, Koh S-H, Park Y-S. Changes in Major Insect Pests of Pine Forests in Korea Over the Last 50 Years. Forests. 2019; 10(8):692. https://0-doi-org.brum.beds.ac.uk/10.3390/f10080692
Chicago/Turabian StyleChoi, Won Il, Youngwoo Nam, Cha Young Lee, Byoung Ki Choi, Yu Jin Shin, Jong-Hwan Lim, Sang-Hyun Koh, and Young-Seuk Park. 2019. "Changes in Major Insect Pests of Pine Forests in Korea Over the Last 50 Years" Forests 10, no. 8: 692. https://0-doi-org.brum.beds.ac.uk/10.3390/f10080692