The Influences of Disturbance Histories and Soil Properties on Aboveground Biomass through Plant Functional Traits in a Tropical Rainforest

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
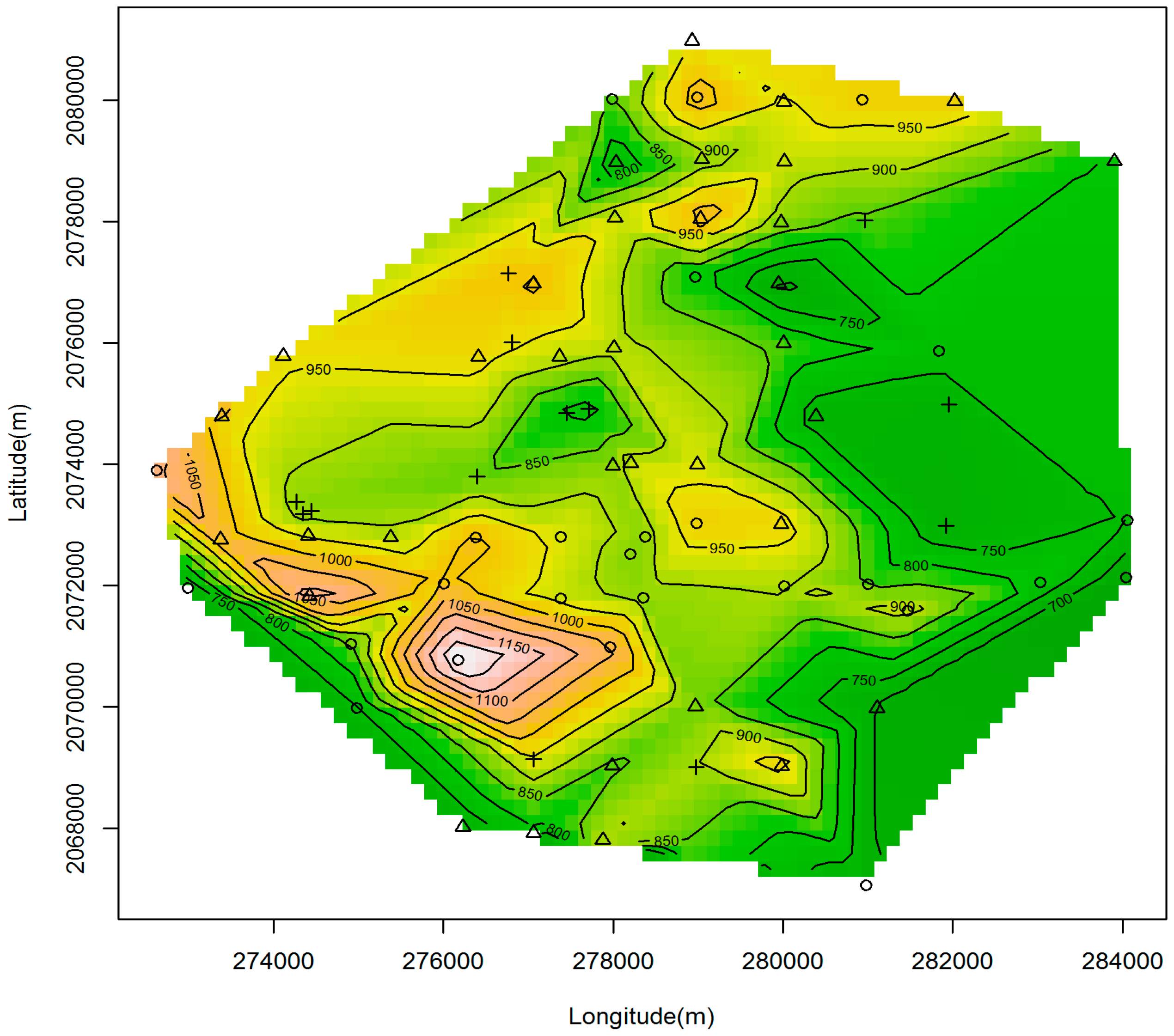
2.1. Study Sites
2.2. Soil Data
2.3. Species and Traits
2.4. Ecosystem Function
2.5. Statistical Analysis
2.5.1. Variations in Community Structure among Forests with Different Disturbance Intensities
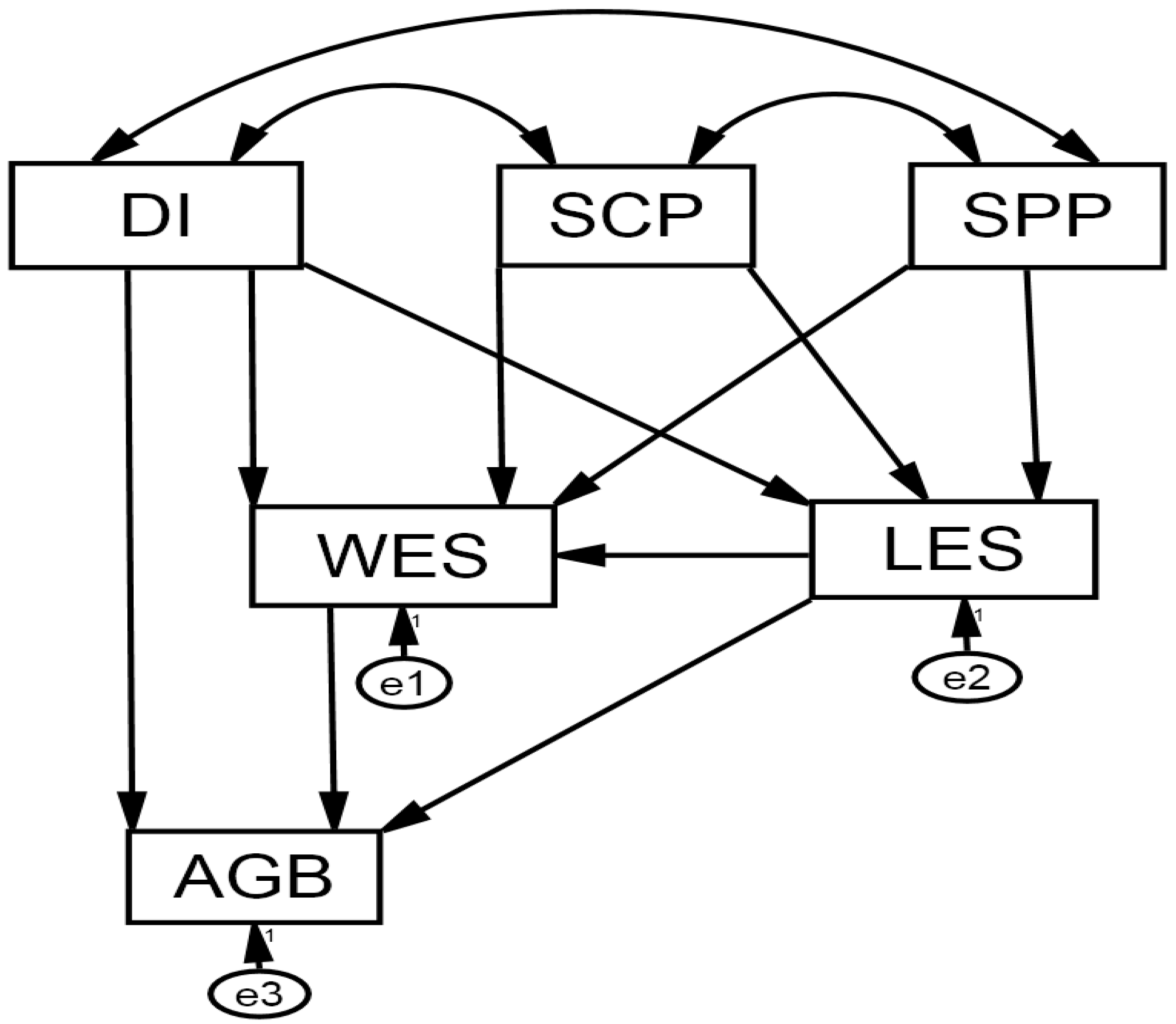
2.5.2. Linkage between Environmental Conditions and Aboveground Biomass through Functional Traits
3. Results
3.1. Variations in Community Structure among Forests with Different Disturbance Intensities
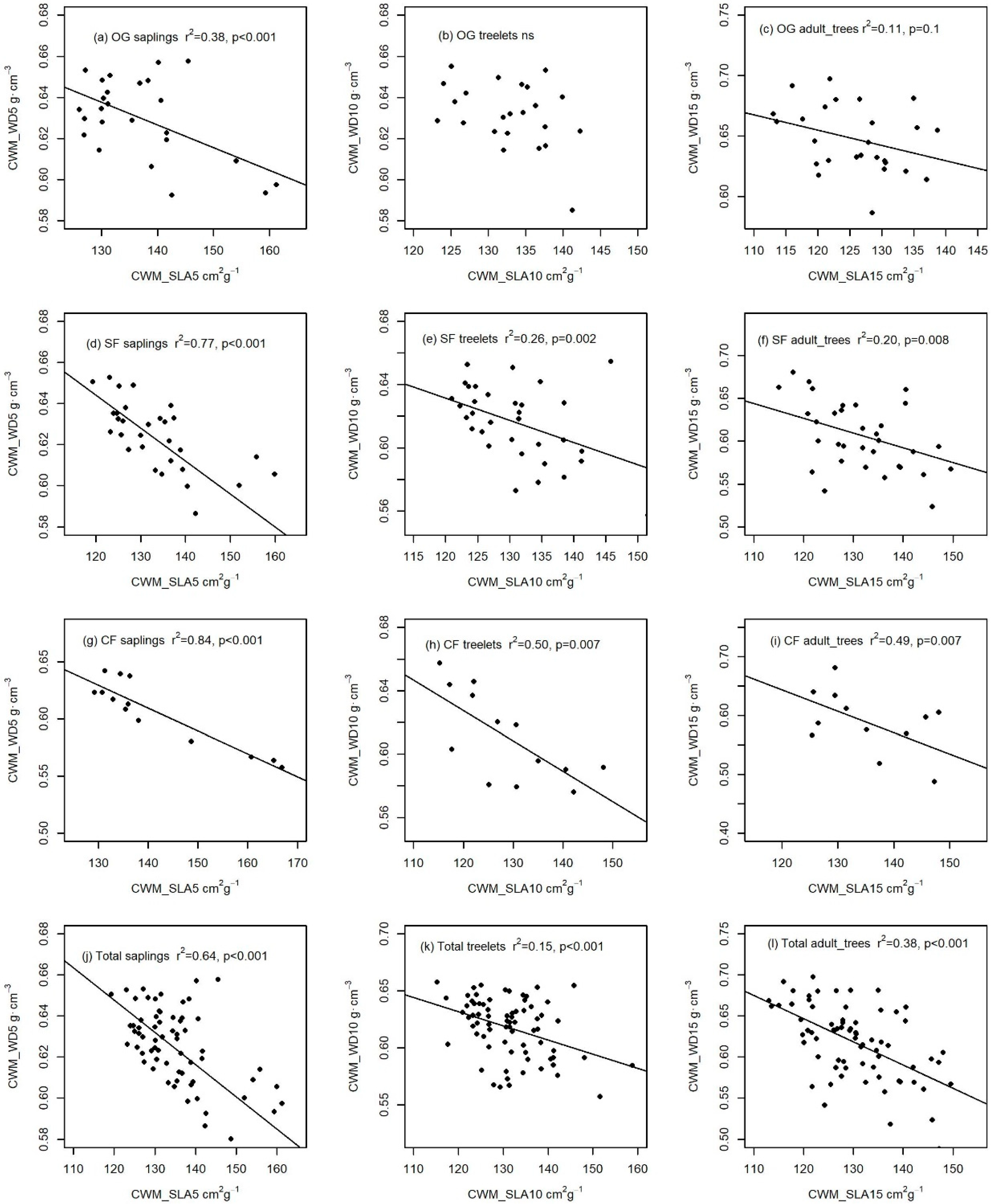
3.2. The Relationships between Community Mean Traits among Forests with Different Disturbance Intensities
3.3. Linkage between Environmental Conditions and Aboveground Biomass through Functional Traits
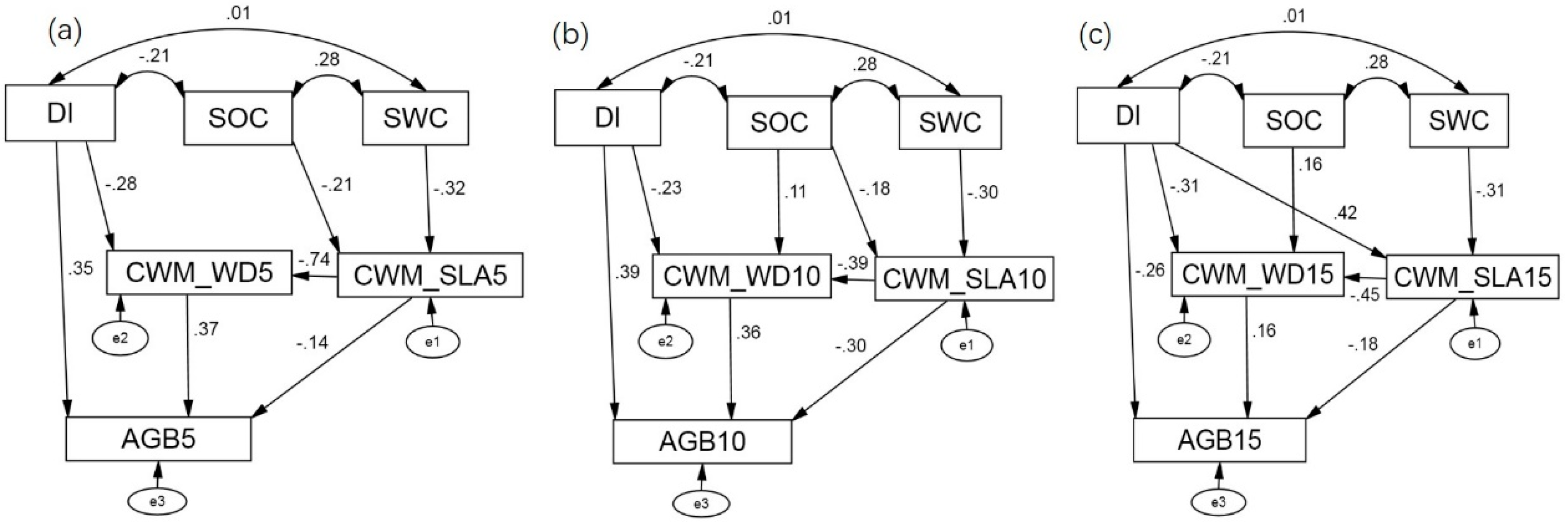
3.3.1. Fitness of Model
3.3.2. Direct Effects
3.3.3. Indirect and Total Effects
4. Discussion
4.1. Variations in Community Structure among Forests with Different Disturbance Intensities
4.2. The Relationships between Community Mean Traits among Forests with Different Disturbance Intensities
4.3. Linkage between Environmental Conditions and Aboveground Biomass through Functional Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; O’Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, B.J.; Matulich, K.L.; Hooper, D.U.; Byrnes, J.E.; Duffy, E.; Gamfeldt, L.; Balvanera, P.; O’Connor, M.I.; Gonzalez, A.; O’Connor, M.I. The functional role of producer diversity in ecosystems. Am. J. Bot. 2011, 98, 572–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef]
- Cardinale, B. Impacts of Biodiversity Loss. Science 2012, 336, 552–553. [Google Scholar] [CrossRef] [PubMed]
- Petchey, O.L. On the statistical significance of functional diversity effects. Funct. Ecol. 2004, 18, 297–303. [Google Scholar] [CrossRef]
- McGill, B.; Enquist, B.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Messier, J.; McGill, B.J.; Lechowicz, M.J. How do traits vary across ecological scales? A case for trait-based ecology. Ecol. Lett. 2010, 13, 838–848. [Google Scholar] [CrossRef]
- Keddy, P.A. Assembly and response rules: Two goals for predictive community ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef]
- Ackerly, D. Functional strategies of chaparral shrubs in relation to seasonal water deficit and disturbance. Ecol. Monogr. 2004, 74, 25–44. [Google Scholar] [CrossRef]
- Katabuchi, M.; Kurokawa, H.; Davies, S.J.; Tan, S.; Nakashizuka, T. Soil resource availability shapes community trait structure in a species-rich dipterocarp forest. J. Ecol. 2011, 100, 643–651. [Google Scholar] [CrossRef]
- Suding, K.N.; Lavorel, S.; Chapin, F.S.; Cornelissen, J.H.C.; Díaz, S.; Garnier, E.; Goldberg, D.; Hooper, D.U.; Jackson, S.T.; Navas, M.-L. Scaling environmental change through the community-level: A trait-based response-and-effect framework for plants. Glob. Change Boil. 2008, 14, 1125–1140. [Google Scholar] [CrossRef]
- Grace, J.B. Structural Equation Modeling and Natural Systems; Cambridge University Press: London, UK, 2006. [Google Scholar]
- Shipley, B. Cause and Correlation in Biology: A User’s Guide to Path Analysis, Structural Equations and Causal Inference; Cambridge University Press: London, UK, 2002. [Google Scholar]
- De Bello, F.; Lavorel, S.; Díaz, S.; Harrington, R.; Cornelissen, J.H.C.; Bardgett, R.D.; Berg, M.P.; Cipriotti, P.; Feld, C.K.; Hering, D.; et al. Towards an assessment of multiple ecosystem processes and services via functional traits. Biodivers. Conserv. 2010, 19, 2873–2893. [Google Scholar] [CrossRef]
- Ding, Y.; Zang, R.; Liu, S.; He, F.; Letcher, S.G. Recovery of woody plant diversity in tropical rain forests in southern China after logging and shifting cultivation. Boil. Conserv. 2012, 145, 225–233. [Google Scholar] [CrossRef]
- Jiang, Y.X.; Wang, B.S.; Zang, R.G.; Jin, J.H.; Liao, W.B. Tropical Forest Biodiversity and Its Forming Mechanism in Hainan Island; Science Press: Beijing, China, 2002. [Google Scholar]
- Zang, R.G.; An, S.Q.; Tao, J.Q. Mechanism of Biodiversity Maintenance of Tropical Forests in Hainan ISLAND; Science Press: Beijing, China, 2004. [Google Scholar]
- Editorial Committee of Flora of China. Flora Reipublicae Popularis Sinicae; Chinese Edition of Flora of China; Science Press: Beijing, China, 2004. [Google Scholar]
- Zhang, W.R.; Yang, G.C.; Tu, X.N. The Forestry Industry Standard of the People’s Republic of China-Determination of Forest Soil; China Standard Press: Beijing, China, 1999. [Google Scholar]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 30 July 2014).
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Change Boil. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M.; Falster, D.S.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- Ordoñez, J.C.; Van Bodegom, P.M.; Witte, J.-P.M.; Wright, I.J.; Reich, P.B.; Aerts, R. A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Glob. Ecol. Biogeogr. 2009, 18, 137–149. [Google Scholar] [CrossRef]
- Pidgen, K.; Mallik, A.U. Ecology of Compounding Disturbances: The Effects of Prescribed Burning After Clearcutting. Ecosystems 2012, 16, 170–181. [Google Scholar] [CrossRef]
- Nishimura, T.B.; Suzuki, E.; Kohyama, T.; Tsuyuzaki, S. Mortality and growth of trees in peat-swamp and heath forests in Central Kalimantan after severe drought. Plant Ecol. 2007, 193, 165–177. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Bernard, C.S.; Breman, F.C.; Van Beek, M.; Salim, A.; Sheil, D. Wood Density as a Conservation Tool: Quantification of Disturbance and Identification of Conservation-Priority Areas in Tropical Forests. Conserv. Boil. 2008, 22, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, J.H.C.; Van Logtestijn, R.S.P.; Freschet, G.T.; Aerts, R. Substantial nutrient resorption from leaves, stems and roots in a subarctic flora: What is the link with other resource economics traits? New Phytol. 2010, 186, 879–889. [Google Scholar]
- Mokany, K.; Ash, J.; Roxburgh, S. Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland. J. Ecol. 2008, 96, 884–893. [Google Scholar] [CrossRef]
- Chao, K.-J.; Phillips, O.L.; Gloor, E.; Monteagudo, A.; Martinez, R.V.; Torres-Lezama, A.; Torres-Lezama, A. Growth and wood density predict tree mortality in Amazon forests. J. Ecol. 2008, 96, 281–292. [Google Scholar] [CrossRef]
- Meigs, G.W.; Donato, D.C.; Campbell, J.L.; Martin, J.G.; Law, B.E. Forest Fire Impacts on Carbon Uptake, Storage, and Emission: The Role of Burn Severity in the Eastern Cascades, Oregon. Ecosystems 2009, 12, 1246–1267. [Google Scholar] [CrossRef]
- Byrne, B. Structural Equation Modeling with AMOS: Basic Concepts, Applications, and Programming; Lawrence Erlbaum Associates: Copenhagen, Denmark, 2001. [Google Scholar]
- Chazdon, R.L. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 51–71. [Google Scholar] [CrossRef]
- Letcher, S.G.; Chazdon, R.L. Rapid Recovery of Biomass, Species Richness, and Species Composition in a Forest Chronosequence in Northeastern Costa Rica. Biotropica 2009, 41, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Cannon, C.H.; Peart, D.R.; Leighton, M. Tree Species Diversity in Commercially Logged Bornean Rainforest. Science 1998, 281, 1366–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredeen, A.L.; Bois, C.H.; Janzen, D.T.; Sanborn, P.T. Comparison of coniferous forest carbon stocks between old-growth and young second-growth forests on two soil types in central British Columbia, Canada. Can. J. For. Res. 2005, 35, 1411–1421. [Google Scholar] [CrossRef]
- Bonnell, T.R.; Reyna-Hurtado, R.; Chapman, C.A. Post-logging recovery time is longer than expected in an East African tropical forest. For. Ecol. Manag. 2011, 261, 855–864. [Google Scholar] [CrossRef]
- Chazdon, R.L. Chance and Determinism in Tropical Forest Succession; Wiley-Blackwell Publishing: Boston, MA, USA, 2008. [Google Scholar]
- Wright, I.J.; Ackerly, D.D.; Bongers, F.; Harms, K.E.; Ibarra-Manriquez, G.; Martinez-Ramos, M.; Mazer, S.J.; Muller-Landau, H.C.; Paz, H.; Pitman, N.C.; et al. Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests. Ann. Bot.-Lond. 2007, 99, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Bucci, S.J.; Scholz, F.G.; Franco, A.C.; Bustamante, M.; Goldstein, G.; Meinzer, F. Functional convergence in hydraulic architecture and water relations of tropical savanna trees: From leaf to whole plant. Tree Physiol. 2004, 24, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Sterck, F.J.; Bongers, F. Crown development in tropical rain forest trees: Patterns with tree height and light availability. J. Ecol. 2001, 89, 1–13. [Google Scholar] [CrossRef]
- Douma, J.C.; De Haan, M.W.A.; Aerts, R.; Witte, J.-P.M.; Van Bodegom, P.M. Succession-induced trait shifts across a wide range of NW European ecosystems are driven by light and modulated by initial abiotic conditions. J. Ecol. 2011, 100, 366–380. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; Van Der Putten, W.H.; Wall, D.H. Ecological Linkages between Aboveground and Belowground Biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef] [Green Version]
- De Deyn, G.B.; Cornelissen, J.H.C.; Bardgett, R.D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol. Lett. 2008, 11, 516–531. [Google Scholar] [CrossRef]
- Pearson, T.R.H.; Burslem, D.F.R.P.; Goeriz, R.E.; Dalling, J.W. Interactions of gap size and herbivory on establishment, growth and survival of three species of neotropical pioneer trees. J. Ecol. 2003, 91, 785–796. [Google Scholar] [CrossRef]
- Würth, M.K.; Pelaez-Riedl, S.; Wright, S.J.; Körner, C. Non-structural carbohydrate pools in a tropical forest. Oecologia 2005, 143, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Minden, V.; Kleyer, M. Testing the effect-response framework: Key response and effect traits determining above-ground biomass of salt marshes. J. Veg. Sci. 2011, 22, 387–401. [Google Scholar] [CrossRef]
- Thomas, S.; Malczewski, G. Wood carbon content of tree species in Eastern China: Interspecific variability and the importance of the volatile fraction. J. Environ. Manag. 2007, 85, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Almeida, S.; Arroyo, L.; Erwin, T.; Killeen, T.J.; Laurance, S.G.; Laurance, W.F.; Lewis, S.L.; Lloyd, J.; Monteagudo, A.; Neill, D.A.; et al. Variation in wood density determines spatial patterns in Amazonian forest biomass. Glob. Change Boil. 2004, 10, 545–562. [Google Scholar]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Laughlin, D.C. Nitrification is linked to dominant leaf traits rather than functional diversity. J. Ecol. 2011, 99, 1091–1099. [Google Scholar] [CrossRef]
- Thompson, K.; Askew, A.P.; Grime, J.P.; Dunnett, N.P.; Willis, A.J. Biodiversity, ecosystem function and plant traits in mature and immature plant communities. Funct. Ecol. 2005, 19, 355–358. [Google Scholar] [CrossRef]
- Baraloto, C.; Rabaud, S.; Molto, Q.; Blanc, L.; Fortunel, C.; Hérault, B.; Dávila, N.; Mesones, I.; Ríos, M.; Valderrama, E.; et al. Disentangling stand and environmental correlates of aboveground biomass in Amazonian forests. Glob. Change Boil. 2011, 17, 2677–2688. [Google Scholar] [CrossRef]
- Paoli, G.D.; Curran, L.M.; Slik, J.W.F. Soil nutrients affect spatial patterns of aboveground biomass and emergent tree density in southwestern Borneo. Oecologia 2008, 155, 287–299. [Google Scholar] [CrossRef]
- Comeau, P.G.; Kimmins, J.P. Above- and below-ground biomass and production of lodgepole pine on sites with differing soil moisture regimes. Can. J. For. Res. 1989, 19, 447–454. [Google Scholar] [CrossRef]
- De Castilho, C.V.; Magnusson, W.E.; De Araújo, R.N.O.; Luizão, R.C.; Luizão, F.J.; Lima, A.P.; Higuchi, N. Variation in aboveground tree live biomass in a central Amazonian Forest: Effects of soil and topography. For. Ecol. Manag. 2006, 234, 85–96. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DI | SOC | SWC | CWM_SLA | CWM_WD | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tot. | Dir. | Ind. | Tot. | Dir. | Ind. | Tot. | Dir. | Ind. | Tot. | Dir. | Ind. | Tot. | Dir. | Ind. | ||
| Saplings | CWM_SLA5 | −0.212 | −0.212 | −0.322 | −0.322 | |||||||||||
| CWM_WD5 | −0.283 | −0.283 | 0.157 | 0.157 | 0.239 | 0.239 | −0.742 | −0.742 | ||||||||
| AGB5 | 0.249 | 0.354 | −0.104 | 0.088 | 0.088 | 0.134 | 0.134 | −0.417 | −0.144 | −0.273 | 0.368 | 0.368 | ||||
| Treelets | CWM_SLA10 | −0.176 | −0.176 | −0.296 | −0.296 | |||||||||||
| CWM_WD10 | −0.232 | −0.232 | 0.176 | 0.108 | 0.068 | 0.115 | 0.115 | −0.389 | −0.389 | |||||||
| AGB10 | 0.306 | 0.39 | −0.084 | 0.117 | 0.117 | 0.132 | 0.132 | −0.444 | −0.304 | −0.141 | 0.361 | 0.361 | ||||
| Adult trees | CWM_SLA15 | 0.422 | 0.422 | −0.31 | −0.31 | |||||||||||
| CWM_WD15 | −0.5 | −0.309 | −0.191 | 0.163 | 0.163 | 0.14 | 0.14 | −0.452 | −0.452 | |||||||
| AGB15 | −0.417 | −0.259 | −0.158 | 0.026 | 0.026 | 0.079 | 0.079 | −0.255 | −0.182 | −0.073 | 0.162 | 0.162 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bu, W.; Zhang, C.; Huang, J.; Zang, R.; Ding, Y.; Xu, H.; Lin, M.; Wang, J. The Influences of Disturbance Histories and Soil Properties on Aboveground Biomass through Plant Functional Traits in a Tropical Rainforest. Forests 2019, 10, 774. https://0-doi-org.brum.beds.ac.uk/10.3390/f10090774
Bu W, Zhang C, Huang J, Zang R, Ding Y, Xu H, Lin M, Wang J. The Influences of Disturbance Histories and Soil Properties on Aboveground Biomass through Plant Functional Traits in a Tropical Rainforest. Forests. 2019; 10(9):774. https://0-doi-org.brum.beds.ac.uk/10.3390/f10090774
Chicago/Turabian StyleBu, Wensheng, Cancan Zhang, Jihong Huang, Runguo Zang, Yi Ding, Han Xu, Mingxian Lin, and Jinsong Wang. 2019. "The Influences of Disturbance Histories and Soil Properties on Aboveground Biomass through Plant Functional Traits in a Tropical Rainforest" Forests 10, no. 9: 774. https://0-doi-org.brum.beds.ac.uk/10.3390/f10090774