Disentangling the Roles of Topography, Patch, and Land Use on Conservation Trait Status of Specialist Birds in Marginal Forest Land Use Types


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Taxa, Specialization, and Trait Status
2.3. Study Predictors
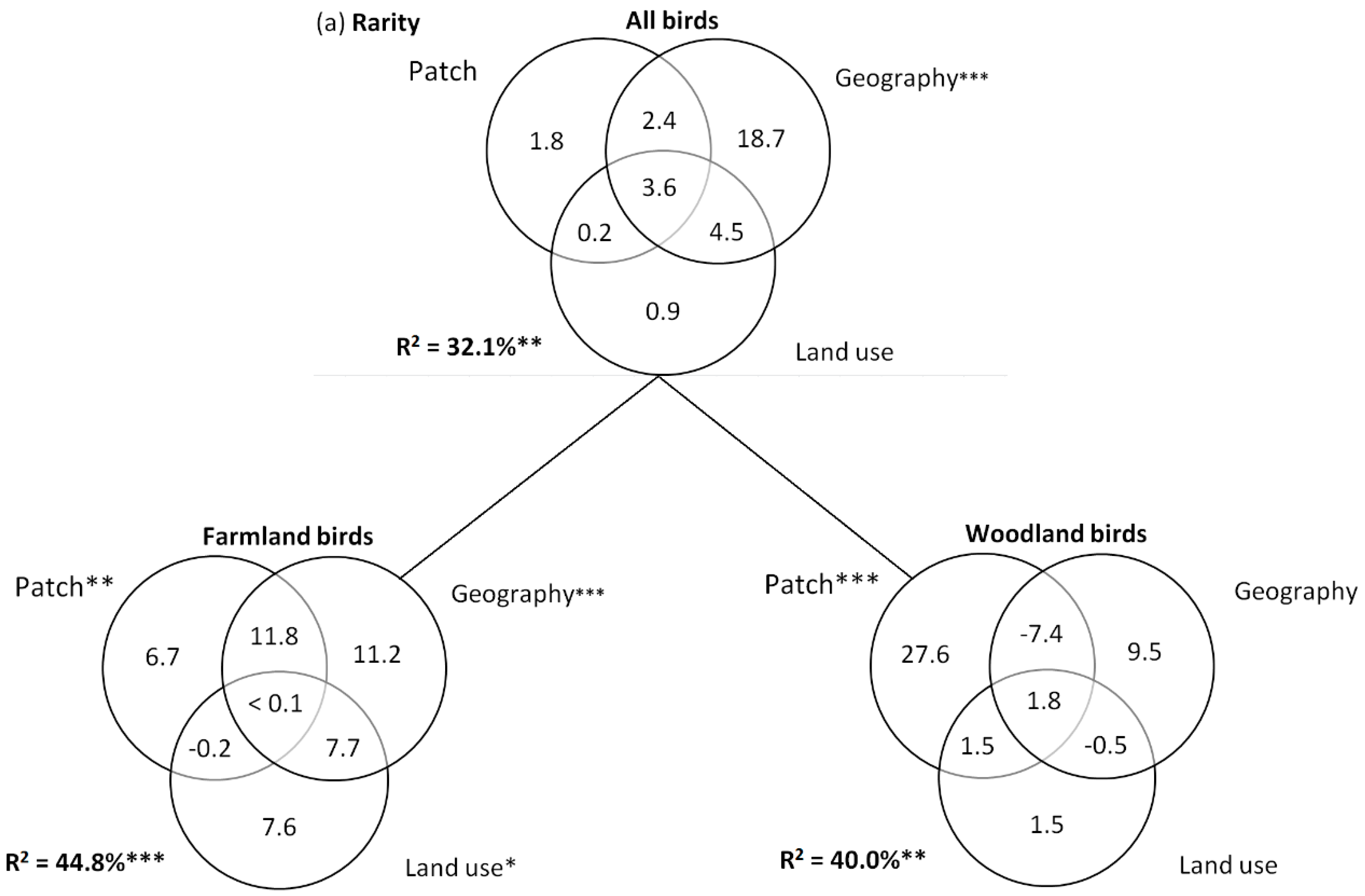
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Rae, L.F.; Whitaker, D.M.; Warkentin, I.G. Multiscale impacts of forest degradation through browsing by hyperabundant moose (Alces alces) on songbird assemblages. Divers. Distrib. 2013, 20, 382–395. [Google Scholar] [CrossRef]
- Müller, J.; Hothorn, T.; Pretzsch, H. Long-term effects of logging intensity on structures, birds, saproxylic beetles and wood-inhabiting fungi in stands of European beech Fagus sylvatica L. Forest Ecol. Manag. 2007, 242, 297–305. [Google Scholar]
- Gates, S.; Donald, P.F. Local extinction of British farmland birds and the prediction of further loss. J. Appl. Ecol. 2000, 37, 806–820. [Google Scholar] [CrossRef]
- Reif, J.; Voříšek, P.; Bejček, V.; Petr, J. Agricultural intensification and farmland birds: New insights from a central European country. Ibis 2008, 150, 596–605. [Google Scholar] [CrossRef]
- McGill, B.J. Matters of scale. Science 2010, 328, 575–576. [Google Scholar] [CrossRef] [Green Version]
- Jackson, H.B.; Fahrig, L. Are ecologists conducting research at the optimal scale? Glob. Ecol. Biogeogr. 2015, 24, 52–63. [Google Scholar] [CrossRef]
- Horak, J.; Peltanova, A.; Podavkova, A.; Safarova, L.; Bogusch, P.; Romportl, D.; Zasadil, P. Biodiversity responses to land use in traditional fruit orchards of a rural agricultural landscape. Agric. Ecosyst. Environ. 2013, 178, 71–77. [Google Scholar] [CrossRef]
- Devictor, V.; Julliard, R.; Clavel, J.; Jiguet, F.; Lee, A.; Couvet, D. Functional biotic homogenization of bird communities in disturbed landscapes. Glob. Ecol. Biogeogr. 2008, 17, 252–261. [Google Scholar] [CrossRef]
- Cepak, J.; Klňava, P.; Škopek, J.; Schröpfer, L.; Jelínek, M.; Hořák, D.; Formánek, J.; Zárybnický, J. Atlas Migrace Ptáků České Republiky a Slovenska; Aventinum: Prague, Czech Republic, 2008. [Google Scholar]
- Reif, J.; Marhoul, P.; Čížek, O.; Konvička, M. Abandoned military training sites are an overlooked refuge for at-risk open habitat bird species. Biodiv. Conserv. 2011, 20, 3645–3662. [Google Scholar] [CrossRef]
- Salek, M. Spontaneous succession on opencast mining sites: Implications for bird biodiversity. J. Appl. Ecol. 2012, 49, 1417–1425. [Google Scholar] [CrossRef]
- Bailey, D.; Schmidt-Entling, M.H.; Eberhart, P.; Herrmann, J.D.; Hofer, G.; Kormann, U.; Herzog, F. Effects of habitat amount and isolation on biodiversity in fragmented traditional orchards. J. Appl. Ecol. 2010, 47, 1003–1013. [Google Scholar] [CrossRef]
- Horak, J. Insect taxa with similar habitat requirements may differ in response to the environment in heterogeneous patches of traditional fruit orchards. J. Insect Conserv. 2014, 18, 637–642. [Google Scholar] [CrossRef]
- Fortin, D.; Arnold, G.W. The influence of road verges on the use of nearby small shrubland remnants by birds in the central wheatbelt of Western Australia. Wildlife Res. 1997, 24, 679–689. [Google Scholar] [CrossRef]
- Horak, J. Fragmented habitats of traditional fruit orchards are important for dead wood-dependent beetles associated with open canopy deciduous woodlands. Naturwissenschaften 2014, 101, 499–504. [Google Scholar] [CrossRef] [PubMed]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Janeček, V.; Rada, P.; Rom, J.; Horák, J. Rural artifacts in a city: Determinants of spatiotemporally continuous fruit orchards in an urban area. Urban For. Urban Green. 2019, 41, 33–38. [Google Scholar] [CrossRef]
- Stastny, K.; Bejcek, V.; Vorisek, P.; Flousek, J. Population trends of farmland and woodland birds in the Czech Republic in 1982–2001 and their use as indicators. Sylvia 2004, 40, 27–48. [Google Scholar]
- Stastny, K.; Bejcek, V.; Hudec, K. Atlas of Breeding Bird Distribution in the Czech Republic; Aventinum: Prague, Czech Republic, 2006. [Google Scholar]
- Barbaro, L.; Rossi, J.P.; Vetillard, F.; Nezan, J.; Jactel, H. The spatial distribution of birds and carabid beetles in pine plantation forests: The role of landscape composition and structure. J. Biogeogr. 2007, 34, 652–664. [Google Scholar] [CrossRef]
- Pysek, P.; Jarošík, V.; Hulme, P.E.; Kühn, I.; Wild, J.; Arianoutsou, M.; Winter, M. Disentangling the role of environmental and human pressures on biological invasions across Europe. Proc. Natl. Acad. Sci. USA 2010, 107, 12157–12162. [Google Scholar] [CrossRef] [Green Version]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward selection of explanatory variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef]
- European Commision. Forest Cover Map. 2019. Available online: http://forest.jrc.ec.europa.eu/. (accessed on 6 December 2019).
- Rangel, T.F.; Diniz-Filho, J.A.F.; Bini, L.M. SAM: A comprehensive application for spatial analysis in macroecology. Ecography 2010, 33, 46–50. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Horak, J.; Pavlicek, J. Tree level indicators of species composition of saproxylic beetles in old-growth mountainous spruce–beech forest through variation partitioning. J. Insect Conserv. 2013, 17, 1003–1009. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Horak, J.; Rom, J.; Rada, P.; Safarova, L.; Koudelkova, J.; Zasadil, P.; Hald, J.P.; Holusa, J. Renaissance of a rural artifart in a city with a million people: Biodiversity responses to an agroforestry restoration in a large urban traditional fruit orchard. Urban Ecosyst. 2018, 21, 263–270. [Google Scholar]
- Cushman, S.A.; McGarigal, K. Hierarchical, multi-scale decomposition of species–environment relationships. Landsc. Ecol. 2002, 17, 637–646. [Google Scholar] [CrossRef]
- Reif, J.; Vermouzek, Z. Collapse of farmland bird populations in an Eastern European country following its EU accession. Conserv. Lett. 2019, 12, e12585. [Google Scholar] [CrossRef]
- Freemark, K.E.; Kirk, D.A. Birds on organic and conventional farms in Ontario: Partitioning effects of habitat and practices on species composition and abundance. Biol. Conserv. 2001, 101, 337–350. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Luoto, M.; Virkkala, R.; Rainio, K. Effects of habitat cover, landscape structure and spatial variables on the abundance of birds in an agricultural–forest mosaic. J. Appl. Ecol. 2004, 41, 824–835. [Google Scholar] [CrossRef]
- Hobson, K.A.; Kirk, D.A.; Smith, A.R. A multivariate analysis of breeding bird species of western and central Canadian boreal forests: Stand and spatial effects. Ecoscience 2000, 7, 267–280. [Google Scholar] [CrossRef]
- Haila, Y.; Nicholls, A.O.; Hanski, I.K.; Raivio, S. Stochasticity in bird habitat selection: Year-to-year changes in territory locations in a boreal forest bird assemblage. Oikos 1996, 76, 536–552. [Google Scholar] [CrossRef]
- Siriwardena, G.M.; Crick, H.Q.P.; Baillie, S.R.; Wilson, J.D. Agricultural land-use and the spatial distribution of granivorous lowland farmland birds. Ecography 2000, 23, 702–719. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Category | Predictor | Mean | SE | Min | Max |
|---|---|---|---|---|---|
| Patch | Canopy openness | 78.1 | 2.2 | 16.0 | 100.0 |
| Shrub coverage | 16.4 | 2.1 | 0.0 | 70.0 | |
| Surrounding | 2.6 | 0.1 | 1.0 | 4.0 | |
| Land use | Rural and urban | 3.0 | 0.3 | 0.9 | 11.8 |
| Forest | 19.2 | 1.5 | 4.6 | 51.5 | |
| Grassland | 12.5 | 0.6 | 3.6 | 31.1 | |
| Arable | 60.0 | 1.9 | 27.2 | 78.5 | |
| Topography | Altitude | 324.3 | 4.5 | 268.8 | 411.3 |
| Area | 6483.5 | 755.3 | 891.0 | 35,391.0 | |
| Longitude | 50.0 | 0.0 | 49.9 | 50.0 | |
| Latitude | 16.2 | 0.0 | 16.1 | 16.3 |
| Species | Preference | Red-List | Grids | Individuals | Presence |
|---|---|---|---|---|---|
| Acrocephalus palustris | General | 52 | 4 | 4 | |
| Aegithalos caudatus | Woodland | 37 | 1 | 1 | |
| Buteo buteo | Woodland | 2 | 1 | 1 | |
| Carduelis cannabina | Farmland | 26 | 4 | 4 | |
| Carduelis carduelis | Farmland | 5 | 18 | 14 | |
| Carduelis chloris | Farmland | 0 | 54 | 52 | |
| Certhia brachydactyla | Woodland | 192 | 3 | 3 | |
| Columba palumbus | Woodland | 2 | 11 | 10 | |
| Coturnix coturnix | Farmland | NT | 41 | 2 | 2 |
| Cyanistes caeruleus | Woodland | 1 | 93 | 67 | |
| Delichon urbicum | General | NT | 1 | 4 | 3 |
| Dendrocopos major | Woodland | 1 | 30 | 30 | |
| Emberiza citronella | Farmland | 0 | 65 | 53 | |
| Erithacus rubecula | Woodland | 1 | 26 | 26 | |
| Ficedula albicollis | Woodland | NT | 297 | 1 | 1 |
| Ficedula hypoleuca | Woodland | NT | 287 | 2 | 2 |
| Fringilla coelebs | Woodland | 0 | 66 | 62 | |
| Garrulus glandarius | Woodland | 3 | 18 | 18 | |
| Hippolais icterina | Woodland | 38 | 17 | 17 | |
| Hirundo rustica | Farmland | 1 | 45 | 33 | |
| Jynx torquilla | Woodland | VU | 196 | 7 | 7 |
| Lanius collurio | Farmland | NT | 3 | 16 | 16 |
| Lanius excubitor | Farmland | VU | 186 | 1 | 1 |
| Locustella naevia | General | 76 | 2 | 2 | |
| Motacilla alba | General | 0 | 12 | 12 | |
| Motacilla cinerea | General | 37 | 1 | 1 | |
| Muscicapa striata | Woodland | 46 | 2 | 2 | |
| Oriolus oriolus | Woodland | 170 | 1 | 1 | |
| Parus major | Woodland | 0 | 79 | 68 | |
| Passer domesticus | General | 4 | 47 | 28 | |
| Passer montanus | Farmland | 36 | 73 | 38 | |
| Perdix perdix | Farmland | NT | 136 | 1 | 1 |
| Periparus ater | Woodland | 34 | 1 | 1 | |
| Phoenicurus ochruros | General | 1 | 36 | 35 | |
| Phoenicurus phoenicurus | Woodland | 31 | 4 | 4 | |
| Phylloscopus collybita | Woodland | 3 | 56 | 53 | |
| Phylloscopus trochilus | Woodland | 10 | 4 | 4 | |
| Pica pica | Farmland | 24 | 10 | 10 | |
| Picus viridis | Woodland | 40 | 2 | 2 | |
| Poecile montanus | Woodland | 136 | 1 | 1 | |
| Poecile palustris | Woodland | 49 | 14 | 14 | |
| Prunella modularis | Woodland | 15 | 1 | 1 | |
| Pyrrhula pyrrhula | Woodland | 68 | 1 | 1 | |
| Saxiola rubetra | Farmland | 103 | 1 | 1 | |
| Serinus serinus | General | 3 | 17 | 17 | |
| Sitta europaea | Woodland | 1 | 2 | 2 | |
| Streptopelia decaocto | General | 18 | 10 | 10 | |
| Streptopelia turtur | Farmland | 32 | 2 | 2 | |
| Sturnus vulgaris | Farmland | 3 | 93 | 44 | |
| Sylvia atricapilla | Woodland | 1 | 55 | 52 | |
| Sylvia borin | Woodland | 46 | 7 | 7 | |
| Sylvia communis | Farmland | 12 | 21 | 20 | |
| Sylvia curruca | Farmland | 14 | 26 | 25 | |
| Turdus merula | Woodland | 1 | 70 | 68 | |
| Turdus philomelos | Woodland | 1 | 35 | 35 | |
| Turdus pilaris | General | 38 | 16 | 10 | |
| Turdus viscivorus | Woodland | 42 | 2 | 2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zasadil, P.; Romportl, D.; Horák, J. Disentangling the Roles of Topography, Patch, and Land Use on Conservation Trait Status of Specialist Birds in Marginal Forest Land Use Types. Forests 2020, 11, 103. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010103
Zasadil P, Romportl D, Horák J. Disentangling the Roles of Topography, Patch, and Land Use on Conservation Trait Status of Specialist Birds in Marginal Forest Land Use Types. Forests. 2020; 11(1):103. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010103
Chicago/Turabian StyleZasadil, Petr, Dušan Romportl, and Jakub Horák. 2020. "Disentangling the Roles of Topography, Patch, and Land Use on Conservation Trait Status of Specialist Birds in Marginal Forest Land Use Types" Forests 11, no. 1: 103. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010103